
Investigation of an EHV-1 Outbreak in the United States Caused by a New H752 Genotype

Abstract
1. Introduction
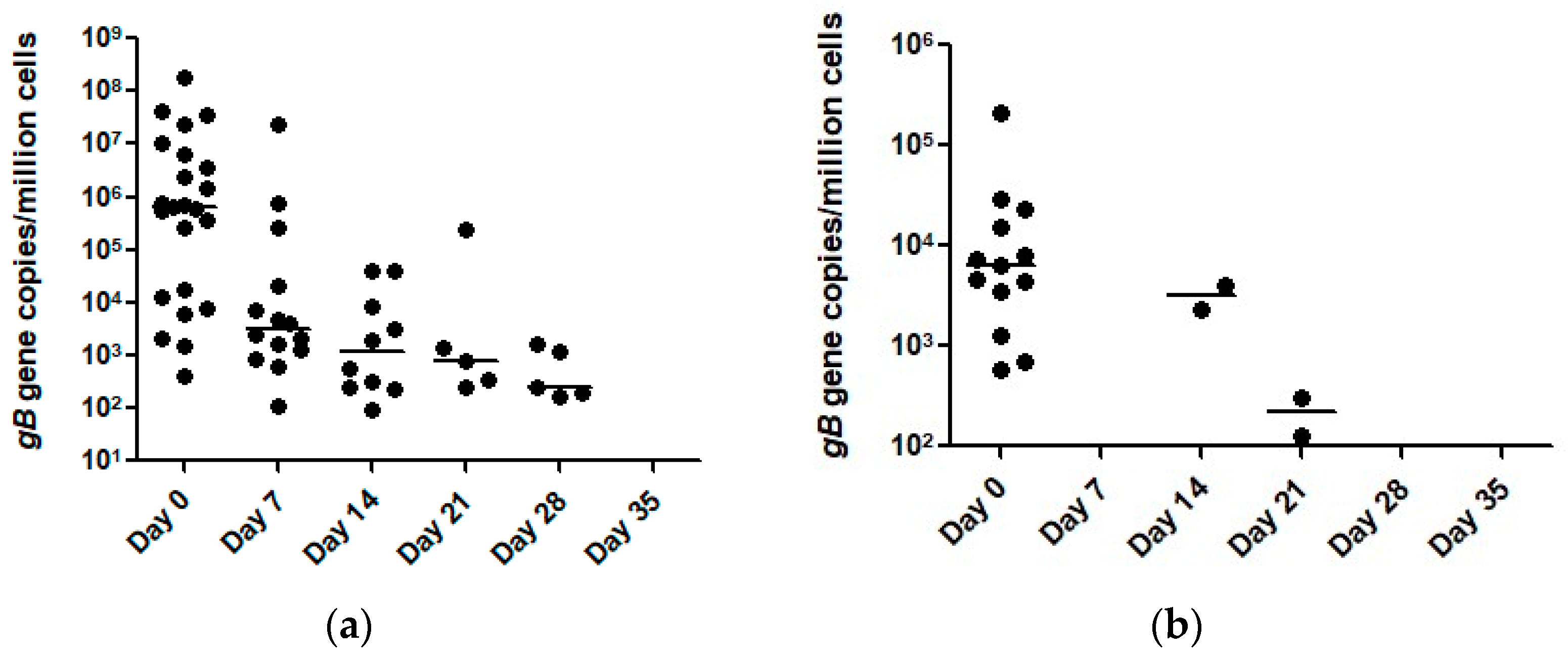
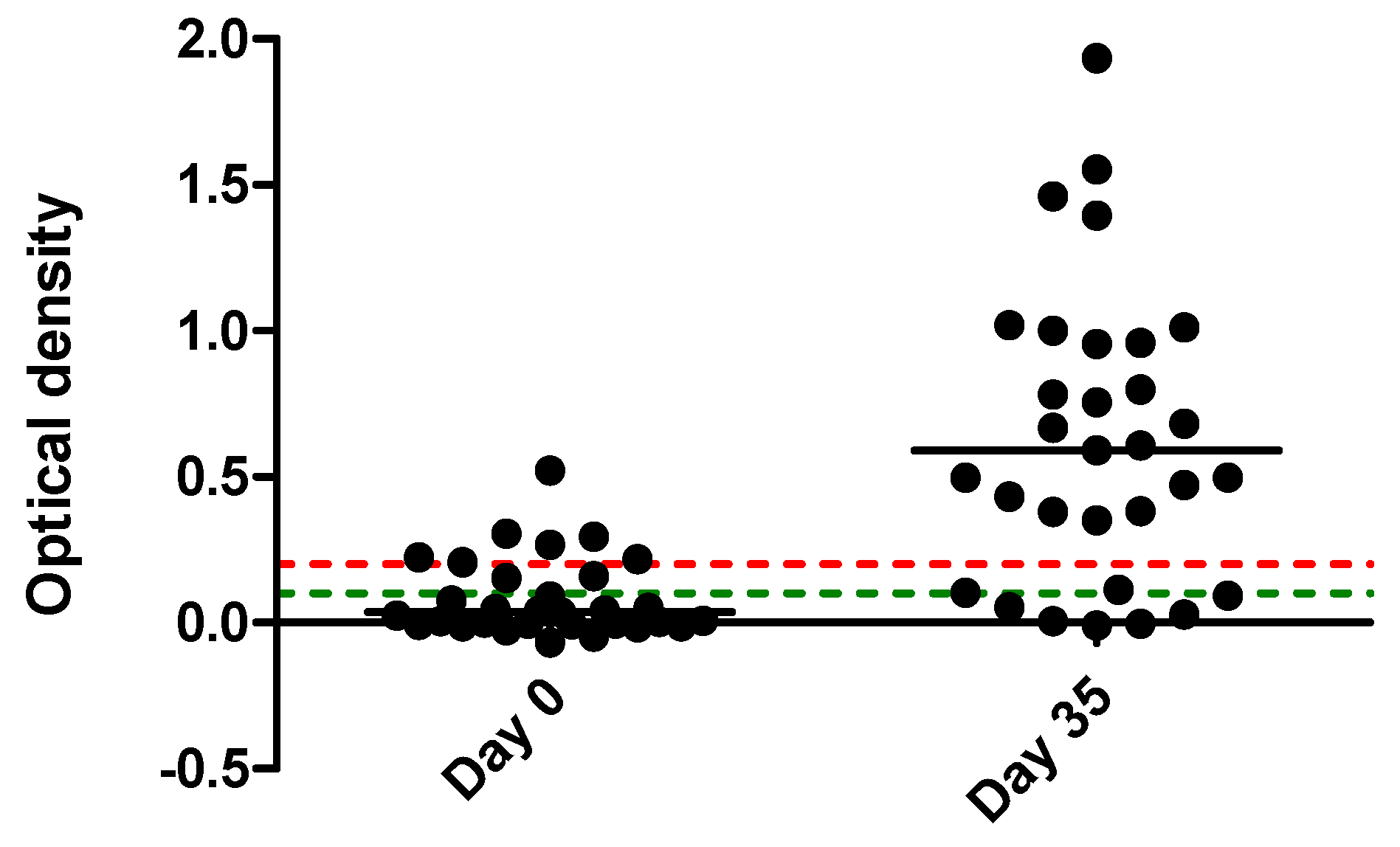
2. Results
3. Discussion
4. Materials and Methods
4.1. Horses
4.2. Quantitative Polymerase Chain Reaction (qPCR) for Detection of EHV-1
4.3. Serology
4.4. Sequencing of an ORF 30 Gene Fragment
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ostlund, E.N. The equine herpesviruses. Vet. Clin. N. Am. Equine. Pract. 1993, 9, 283–294. [Google Scholar] [CrossRef]
- Henninger, R.W.; Reed, S.M.; Saville, W.J.; Allen, G.P.; Hass, G.F.; Kohn, C.W.; Sofaly, C. Outbreak of neurologic disease caused by equine herpesvirus-1 at a university equestrian center. J. Vet. Intern. Med. 2007, 21, 157–165. [Google Scholar] [CrossRef]
- Burgess, B.A.; Tokateloff, N.; Manning, S.; Lohmann, K.; Lunn, D.P.; Hussey, S.B.; Morley, P.S. Nasal shedding of equine herpesvirus-1 from horses in an outbreak of equine herpes myeloencephalopathy in Western Canada. J. Vet. Intern. Med. 2012, 26, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Traub-Dargatz, J.L.; Pelzel-McCluskey, A.M.; Creekmore, L.H.; Geiser-Novotny, S.; Kasari, T.R.; Wiedenheft, A.M.; Bush, E.J.; Bjork, K.E. Case-control study of a multistate equine herpesvirus myeloencephalopathy outbreak. J. Vet. Intern. Med. 2013, 27, 339–346. [Google Scholar] [CrossRef]
- McFadden, A.M.; Hanlon, D.; McKenzie, R.K.; Gibson, I.; Bueno, I.M.; Pulford, D.J.; Orr, D.; Dunowska, M.; Stanislawek, W.L.; Spence, R.P.; et al. The first reported outbreak of equine herpesvirus myeloencephalopathy in New Zealand. N. Z. Vet. J. 2016, 64, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Lesté-Lasserre, C. Deadly viral outbreak ravages European horses. Science 2021, 371, 1297. [Google Scholar] [CrossRef]
- Nugent, J.; Birch-Machin, I.; Smith, K.C.; Mumford, J.A.; Swann, Z.; Newton, J.R.; Bowden, R.J.; Allen, G.P.; Davis-Poynter, N. Analysis of equid herpesvirus 1 strain variation reveals a point mutation of the DNA polymerase strongly associated with neuropathogenic versus nonneuropathogenic disease outbreaks. J. Virol. 2006, 80, 4047–4060. [Google Scholar] [CrossRef]
- Perkins, G.A.; Goodman, L.B.; Tsujimura, K.; Van de Walle, G.R.; Kim, S.G.; Dubovi, E.J.; Osterrieder, N. Investigation of the prevalence of neurologic equine herpes virus type 1 (EHV-1) in a 23-year retrospective analysis (1984-2007). Vet. Microbiol. 2009, 139, 375–378. [Google Scholar] [CrossRef]
- Sutton, G.; Thieulent, C.; Fortier, C.; Hue, E.S.; Marcillaud-Pitel, C.; Pléau, A.; Deslis, A.; Guitton, E.; Paillot, R.; Pronost, S. Identification of a new equid herpesvirus 1 DNA polymerase (ORF30) genotype with the isolation of a C(2254)/H(752) strain in French horses showing no major impact on the strain behaviour. Viruses 2020, 12, 1160. [Google Scholar] [CrossRef]
- Goodman, L.B.; Loregian, A.; Perkins, G.A.; Nugent, J.; Buckles, E.L.; Mercorelli, B.; Kydd, J.H.; Palù, G.; Smith, K.C.; Osterrieder, N.; et al. A point mutation in a herpesvirus polymerase determines neuropathogenicity. PLoS Pathog. 2007, 3, e160. [Google Scholar] [CrossRef]
- Allen, G.P. Development of a real-time polymerase chain reaction assay for rapid diagnosis of neuropathogenic strains of equine herpesvirus-1. J. Vet. Diagn. Invest. 2007, 19, 69–72. [Google Scholar] [CrossRef]
- Smith, K.L.; Li, Y.; Breheny, P.; Cook, R.F.; Henney, P.J.; Sells, S.; Pronost, S.; Lu, Z.; Crossley, B.M.; Timoney, P.J.; et al. New real-time PCR assay using allelic discrimination for detection and differentiation of equine herpesvirus-1 strains with A2254 and G2254 polymorphisms. J. Clin. Microbiol. 2012, 50, 1981–1988. [Google Scholar] [CrossRef] [PubMed]
- Leutenegger, C.M.; Madigan, J.E.; Mapes, S.; Thao, M.; Estrada, M.; Pusterla, N. Detection of EHV-1 neuropathogenic strains using real-time PCR in the neural tissue of horses with myeloencephalopathy. Vet. Rec. 2008, 162, 688–690. [Google Scholar] [CrossRef]
- Balasuriya, U.B.; Crossley, B.M.; Timoney, P.J. A review of traditional and contemporary assays for direct and indirect detection of equid herpesvirus 1 in clinical samples. J. Vet. Diagn. Invest. 2015, 27, 673–687. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, L.K.; Bentz, B.G.; Gilliam, L.L.; Ritchey, J.W.; Pusterla, N.; Eberle, R.; Holbrook, T.C.; McFarlane, D.; Rezabek, G.B.; Meinkoth, J.; et al. Efficacy of the early administration of valacyclovir hydrochloride for the treatment of neuropathogenic equine herpesvirus type-1 infection in horses. Am. J. Vet. Res. 2017, 78, 1126–1139. [Google Scholar] [CrossRef]
- Shiraki, K. Antiviral drugs against alphaherpesvirus. Adv. Exp. Med. Biol. 2018, 1045, 103–122. [Google Scholar] [CrossRef] [PubMed]
- Estell, K.E.; Dawson, D.R.; Magdesian, K.G.; Swain, E.; Laing, S.T.; Siso, S.; Mapes, S.; Pusterla, N. Quantitative molecular viral loads in 7 horses with naturally occurring equine herpesvirus-1 infection. Equine Vet. J. 2015, 47, 689–693. [Google Scholar] [CrossRef]
- Pusterla, N.; Hussey, G.S. Equine herpesvirus 1 myeloencephalopathy. Vet. Clin. N. Am. Equine Pract. 2014, 30, 489–506. [Google Scholar] [CrossRef]
- Walter, J.; Seeh, C.; Fey, K.; Bleul, U.; Osterrieder, N. Prevention of equine herpesvirus myeloencephalopathy—Is heparin a novel option? A case report. Tierarztl Prax Ausg G Grosstiere Nutztiere 2016, 44, 313–317. [Google Scholar] [CrossRef]
- Thirumalapura, N.; Tewari, D.; Simeone, A.; Brightbill, K.; Anis, E.; Wheeler-Aceto, H.; Dallap-Schaer, B. First reported detection of the equine herpesvirus 1 DNA polymerase 2254C/His752 variant in horses in the United States. J. Am. Vet. Med. Assoc. 2021, 258, 1181–1183. [Google Scholar] [CrossRef]
- Koppers-Lalic, D.; Reits, E.A.; Ressing, M.E.; Lipinska, A.D.; Abele, R.; Koch, J.; Marcondes Rezende, M.; Admiraal, P.; van Leeuwen, D.; Bienkowska-Szewczyk, K.; et al. Varicelloviruses avoid T cell recognition by UL49.5-mediated inactivation of the transporter associated with antigen processing. Proc. Natl. Acad. Sci. USA 2005, 102, 5144–5149. [Google Scholar] [CrossRef] [PubMed]
- van der Meulen, K.M.; Favoreel, H.W.; Pensaert, M.B.; Nauwynck, H.J. Immune escape of equine herpesvirus 1 and other herpesviruses of veterinary importance. Vet. Immunol. Immunopathol. 2006, 111, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Pusterla, N.; Wilson, W.D.; Mapes, S.; Finno, C.; Isbell, D.; Arthur, R.M.; Ferraro, G.L. Characterization of viral loads, strain and state of equine herpesvirus-1 using real-time PCR in horses following natural exposure at a racetrack in California. Vet. J. 2009, 179, 230–239. [Google Scholar] [CrossRef]
- Gilkerson, J.R.; Whalley, J.M.; Drummer, H.E.; Studdert, M.J.; Love, D.N. Epidemiological studies of equine herpesvirus 1 (EHV-1) in Thoroughbred foals: A review of studies conducted in the Hunter Valley of New South Wales between 1995 and 1997. Vet. Microbiol. 1999, 68, 15–25. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Horse | Clinical Signs | qPCR | Serology | |||
|---|---|---|---|---|---|---|
| Fever in °C (Peak/Duration) | Other Signs | Nasal Shedding | Viremia | Day 0 | Day 35 | |
| 1 | 38.8/6 | - | - | - | - | - |
| 2 | 39.3/2 | DLE | + | + | - | - |
| 3 | No | - | + | + | + | + |
| 4 | 40.6/3 | - | + | + | doubtful | + |
| 5 | 39.7/3 | - | + | + | - | + |
| 6 | 39.9/4 | - | + | + | + | + |
| 7 | 39.6/6 | - | + | - | + | + |
| 8 | 39.3/1 | - | + | - | - | + |
| 9 | 38.9/1 | - | + | - | - | + |
| 10 | 40.1/2 | A | - | + | - | doubtful |
| 11 | 38.8/1 | A | + | + | - | - |
| 12 | 38.9/1 | - | + | + | - | + |
| 13 | 40.1/3 | A | + | + | - | + |
| 14 | 38.7/2 | - | + | - | - | + |
| 15 | 39.2/1 | - | + | - | - | - |
| 16 | 38.7/1 | - | + | - | - | - |
| 17 | 38.8/1 | - | + | - | + | + |
| 18 | 40.9/5 | ND | + | + | - | + |
| 19 | 41.0/5 | A | + | + | - | + |
| 20 | 40.5/8 | A | + | - | - | + |
| 21 | No | - | + | - | - | + |
| 22 | 39.7/4 | A,ND | + | + | - | + |
| 23 | 40.1/5 | - | + | + | + | + |
| 24 | 39.4/2 | ND | - | + | doubtful | + |
| 25 | No | - | + | - | + | + |
| 26 | 39.1/4 | - | - | + | - | + |
| 27 | 39.3/4 | - | + | + | + | + |
| 28 | 40.6/6 | DLE | + | + | - | + |
| 29 | No | - | + | - | - | + |
| 30 | 40.9/2 | A,DLE,ND | - | + | - | doubtful |
| 31 | No | - | - | - | - | - |
| Total | Fever (26 horses) No fever (5) | A (7) DLE (3) ND (4) | negative (5) positive (26) | negative (13) positive (18) | negative (22) doubtful (2) positive (7) | negative (6) doubtful (2) positive (23) |
| Day | qPCR Results of Nasal Secretions | qPCR Results of Blood | ||
|---|---|---|---|---|
| pos/neg | Range (Median) | pos/neg | Range (Median) | |
| 0 | 23/8 | 412–1.78 × 108 (6.3 × 105) | 13/18 | 572–2.1 × 105 (6269) |
| 7 | 14/17 | 105–2.34 × 107 (3203) | 0/31 | 0 |
| 14 | 10/21 | 94–4.10 × 104 (1204) | 2/29 | 2328–3987 (3158) |
| 21 | 5/26 | 245–2.30 × 105 (800) | 2/29 | 127–311 (219) |
| 28 | 5/26 | 169–1636 (241) | 0/31 | 0 |
| 35 | 0/31 | 0 | 0/31 | 0 |
| Drug | Dose | Treatment Days (Median) | Horses Treated |
|---|---|---|---|
| Valacyclovir | Loading 30 mg/kg q8h PO for first 6 doses Maintenance 20 mg/kg q12h PO | 11–25 (12) | 31 |
| Flunixin meglumine | 0.5–1.1 mg/kg q12h to q24h PO or IV | 1–20 (4) | 26 |
| Sodium heparin | 50 IU/kg q12h SQ | 1–3 (3) | 26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pusterla, N.; Barnum, S.; Miller, J.; Varnell, S.; Dallap-Schaer, B.; Aceto, H.; Simeone, A. Investigation of an EHV-1 Outbreak in the United States Caused by a New H752 Genotype. Pathogens 2021, 10, 747. https://doi.org/10.3390/pathogens10060747
Pusterla N, Barnum S, Miller J, Varnell S, Dallap-Schaer B, Aceto H, Simeone A. Investigation of an EHV-1 Outbreak in the United States Caused by a New H752 Genotype. Pathogens. 2021; 10(6):747. https://doi.org/10.3390/pathogens10060747
Chicago/Turabian StylePusterla, Nicola, Samantha Barnum, Julia Miller, Sarah Varnell, Barbara Dallap-Schaer, Helen Aceto, and Aliza Simeone. 2021. "Investigation of an EHV-1 Outbreak in the United States Caused by a New H752 Genotype" Pathogens 10, no. 6: 747. https://doi.org/10.3390/pathogens10060747
APA StylePusterla, N., Barnum, S., Miller, J., Varnell, S., Dallap-Schaer, B., Aceto, H., & Simeone, A. (2021). Investigation of an EHV-1 Outbreak in the United States Caused by a New H752 Genotype. Pathogens, 10(6), 747. https://doi.org/10.3390/pathogens10060747