1. Introduction
Equid herpesvirus-1 (EHV-1), a member of the Alphaherpesvirinae subfamily, causes respiratory disease, abortion, equid herpesvirus-associated myeloencephalopathy (EHM), chorioretinopathy, and infection of gonads of the intact male following a cell-associated viremia in peripheral blood mononuclear cells (PBMC) [
1]. EHV-1 is typically spread by direct nose-to-nose contact followed by viral replication in the upper respiratory tract epithelium. This active lytic infection includes viremia and spread to the vascular endothelia of secondary sites of infection. After or even during the lytic phase, EHV-1 prepares for its characteristic chronic-persistent phase. This likely life-long infection phase is known as latent infection. In contrast to lytic infection, where transcriptional activity of the entire viral genome is fully upregulated, transcriptional activity during latency is likely limited to a single region known as the immediate early gene (IE) [
2]. Current thoughts are that a latent infection is established early in life during first-time respiratory tract infection with EHV-1. It has been shown that the trigeminal ganglion (TG) is a site of EHV-1 latency that is likely reached via retrograde transportation along nerve axons [
3,
4]. In addition to the TG, EHV-1 latency has also been shown in the mandibular and retropharyngeal lymph node as part of the respiratory tract-associated lymphoid tissue (RALT), as well as in randomly collected circulating CD8+ T-lymphocytes [
4,
5]. However, we have recently provided evidence that cell-associated viremia might also be the vehicle of additional latency establishment at other sites by Day 30 post-infection (p.i.) [
6]. In other members of the Alphaherpesvirinae, such as the Feline herpesvirus-1 (FeHV-1) and the human Varicella-Zoster Virus (VZV) additional latency sites have been detected at several sites of neural parenchyma. For the Bovine herpesvirus-1 (BoHV-1) and the Suid herpesvirus-1 (SHV-1) additional sites have been detected in lymphoid tissue [
7,
8,
9,
10,
11,
12,
13]. We postulate that cell-associated viremia in PBMC is likely the carrier vehicle to other lymphatic or neural tissue sites in addition to TG and RALT.
In 2006, Nugent et al. showed a single nucleotide point (SNP) mutation in the EHV-1 DNA polymerase gene at position 2254 resulting in an amino acid switch (N vs. D752). The D-variants, retrospectively, have shown to be more often associated with clinical myelopathy than N-variants. This might be explained by a higher magnitude and duration of viremia of the D-variants [
14]. Furthermore, EHV-4 is a closely related (equine alpha-) herpesvirus to EHV-1. Both viruses have a type-specific glycoprotein D (gD) in their respective envelopes, which is important for effective viral cell-to-cell spread. The capacity of EHV-1 gD for cell-to-cell transmission is thought to be greater than that for EHV-4 [
15]. Therefore, we established three experimental infection groups for the present study. Group one horses were infected with the EHV-1 Ab4 wild-type (WT) strain with confirmed D752. Group two horses were infected with an EHV-1 Ab4 viral strain mutant where D was switched with N in the polymerase gene (EHV-1 Ab4 N752). Group three horses were infected with an Ab4 strain where the EHV-1 gD gene sequence was exchanged with the corresponding EHV-4 gD sequence (EHV-1 Ab4 gD4). Clinical and immunological data following infection with the different viruses has already been described previously [
16].
Horses from all three groups were subsequently euthanized at 70 days post infection (dpi). The objective of the current study was to investigate the distribution of viral genomic DNA into various lymphoid and neural tissues by Day 70 p.i. and to investigate whether there is a difference in viral genomic DNA distribution pattern of EHV-1 Ab4 WT, the polymerase mutant (EHV-1 Ab4 N752) and the gD EHV-1/EHV-4 replacement mutant (EHV-1 Ab4 gD4). Our hypothesis was that there will be a wider distribution of EHV-1 beyond the currently accepted paradigm and that differences will exist between viral mutants.
3. Discussion
In the present study, EHV-1 gDNA was detected in a variety of neural and lymphatic tissues throughout the body at 70 dpi. This extends our current knowledge about EHV-1 and shows additional sites of latency beyond the TG and RALT including the mesenteric lymph nodes and spleen, as well as a variety of somatic sensory and S/PS ganglia. Late gene expression and specific EHV immunolabeling were consistently absent further suggesting the viral DNA to be a result of an arrested, potentially latent stage of EHV-1 chronic-persistent infection.
The TG as well as the mandibular and retropharyngeal lymph nodes have long been reported sites of EHV-1 latency [
1,
3,
4,
6]. Our findings are consistent with these reports. Furthermore, detection of EHV-1 Ab4 WT in the TG and the retropharyngeal lymph node has been recently described [
6]. Previously, the bronchial lymph node of horses has also been described as a site of EHV-1 latency [
4]. Interestingly, our study shows that while we detected EHV-1 gDNA in this location, the bronchial lymph node was the least likely location of the RALT group to contain EHV-1 gDNA. On the other hand, our study reports EHV-1 gDNA detection in the equine pharyngeal roof, which has not been previously shown. These latter findings are consistent with those shown for other members of the Alphaherpesvirinae, such as BoHV-1 and SHV-1, which have been detected in pharyngeal lymphoid tissue of latently infected calves and piglets respectively, suggesting that the tonsil is a site for viral persistence [
10,
11,
12,
13]. It is plausible that the pharyngeal roof with many primary lymph follicles and the secondary lymph nodes of the upper respiratory tract are major sites for EHV-1 chronic-persistent infection in horses, however, follow-up studies in a larger number of horses of different ages, breeds and at different stages following primary infection or reactivation should examine this question further. Of interest, with this site would be the proximity to the upper respiratory epithelium, as it would allow for quick reactivation and viral nasal shedding. However, the route of infection, dose of viral inoculum and nasopharyngeal instillation of the inoculum, could also be the reason that the lymphoid tissue of the pharyngeal roof and the draining lymph nodes of the upper respiratory tract are primary sites of viral DNA recovery in our study, rather than the further physically distanced bronchial lymph node.
The detection of EHV-1 gDNA in the spleen and mesenteric lymph node samples collected 70 dpi is a novel finding. However, the presence of the EHV-1 Ab4 WT strain in the mesenteric lymph node 70 dpi has been described previously by Giessler et al. from our group [
6]. Interestingly, the spleen has been described as a site of BoHV-1 latency in experimentally infected cattle [
17]. Until now, BoHV-1 was the only Alphaherpesvirus member in which splenic persistence had been documented and it has been shown that in this case immunocompetent lymphocytes reach the spleen via blood vessels and proliferate locally [
18]. During viremia, EHV-1 gDNA has been identified in CD8+ T-lymphocytes, B-lymphocytes, monocytes, and less frequently in CD4+ T-lymphocytes of experimentally infected horses [
19]. This finding suggests that during a productive EHV-1 viremia, the virus may reach the spleen and could be retained in splenic germinal centers during later stages of infection. Alternatively, the detection of EHV-1 gDNA in splenic tissue may be due to its presence in circulating persistently infected lymphocytes rather than true localization to splenic tissue. Furthermore, lymphocytes home repeatedly to secondary lymphoid organs, including the spleen and lymph nodes, reside in these organs transiently, and return to the circulation [
18]. In addition, in this study, all but one horse had EHV-1 gDNA positive mesenteric lymph nodes that were also positive in various other RALT and/or spleen. Although viral gDNA was present in a variety of lymphoid tissue, PBMC isolated on Day 70 were negative for viral gDNA in all but one horse. This was in contrast to several other studies that have reported persistent EHV-1 infection in PBMC and might be explained by the fact that EHV-1 DNA is only present at very low levels, which were not detected with our assay system. Alternatively, presence in circulating PBMC maybe more intermittent or depend on how long ago a previous infection or reactivation has occurred. It remains to be determined whether the abdominal lymphoid tissues represent a true localization for EHV-1 during viral persistence or if this finding is a consequence of recirculation of infected lymphocytes and therefore a transient phenomenon.
In addition to the identification of various lymphoid tissues, this study also identified novel sites for EHV-1 persistence or latency in a variety of neural tissues. Localization of latency in the TG is well-documented for EHV-1 and assumed to occur following retrograde axonal transport along the trigeminal nerve after primary infection of the nasopharyngeal mucosa. However, the mechanisms through which EHV-1 reached various other neural locations throughout the body in the subjects of this study remains unknown. The presence of EHV-1 gDNA in the ciliary (GCi) and cranial cervical (GCcr) ganglia could be due to the close proximity of primary viral replication in the conjunctiva or in the guttural pouch (GP) epithelium. The virus may establish persistence at these sites using retrograde axonal transport, similar to the TG pathway. These findings are consistent with a recent publication where FeHV-1 was detected in a variety of neural tissues of experimentally infected cats [
8,
9]. Alternatively, in particular, the detection of primary lymphoid follicles of the GP epithelium and the proximity of enrolled GCcr preparations in this study could have been the cause for false-positive results in the GCcr tissue analysis.
However, EHV-1 gDNA was also detected in the farther distanced caudal cervical (GCca), mesenteric (GM) and celiac (GCoe) ganglia, as well as in the ST and DRG. Please note, that EHV-1 Ab4 WT in the ST 70 dpi has been demonstrated previously [
6]. As these sites are not in the vicinity to the primary site of infection, it is not clear how the virus reached these ganglia to establish a putative latent infection. The presence of VZV, a close relative to EHV-1, has also been demonstrated in various sensory and autonomic ganglia in addition to TG [
7,
20,
21,
22,
23,
24,
25,
26]. Furthermore, there is growing evidence that VZV most likely uses infected lymphocytes as vehicles to reach ganglion cells during cell-associated viremia [
27,
28,
29]. A study by Giessler et al., 2020 from our group recently showed EHV-1 gDNA positive in-situ hybridization (ISH) signal in support or interstitial cells that gather around neural cell bodies in ganglia by Day 30 p.i. and CD3+ T-lymphocytes could be located frequently in close proximity to the neuronal cell body using IHC. In contrast, ISH positive neural cell bodies were seen by Day 70 p.i. [
6]. Although the non-neural ISH signal could not be definitely attributed to either lymphocytes or peripheral glial cells (satellite glial cells), these findings combined with the numerous EHV-1 gDNA positive abdominal neural tissues detected in the present study, suggest viremia as an alternative route for latency establishment for EHV-1.
Viremia is also crucial to EHV-1 pathogenesis and the prerequisite for complications of abortion or myelopathy. Our findings are highly suggestive that this route also plays a role in the transport of virus to distant neural sites, rather than axonal transportation. Viral DNA has been detected in all PBMC subpopulations of experimentally infected horses during cell-associated viremia of acute infection followed by lymphocytic/monocytic extravasation or endothelial (contact-induced) infection [
19,
30]. An alternative explanation could be that our detected viral DNA remains in the endothelial cells of arterioles, venules, or support cells in the vicinity to ganglia or within perivascular mononuclear cells, as previously described during lytic infection of the uterine tissue and the spinal cord [
31,
32]. At this point we have yet to elucidate whether EHV-1 gDNA is present in ganglion cells; in ganglion support (or satellite) cells, or in infiltrative leukocytes or histiocytes. Our attempts to correlate absent/low, moderate or high lymphocytic/monocytic infiltration with results of EHV-1 gDNA quantification appeared not sensitive enough to provide an answer to this question, as most of the neural parenchyma contained only mild lymphocytic infiltrates.
EHV-1 latency defined as gDNA and latency associated transcripts (LAT) as mRNA have been reported to be established by Day 21 p.i. [
1]. Samples in the present study were obtained around Day 70 p.i. As expected, this was enough time to establish a chronic-persistent, non-lytic/arrested, potentially latent EHV-1 infection in the animals of this study. EHV-1 latency has previously been defined as a stage characterized by absence of viral replication, where transcriptional activity is limited to a region antisense to the immediate early (IE) gene [
1,
2,
5,
33,
34,
35]. In this study, we assumed EHV-1’s arrested/latent state when a sample was EHV-1 gDNA positive while simultaneously, late (L) gene mRNA expression and (lytic) protein expression was absent [
4,
36,
37].
Efstathiou et al. (2005) described the several weeks p.i. phase of HSV-1 infection as an ongoing process of transition from simultaneous low-grade lytic and latent infection activity in different cells of the same tissue. Their study suggests that cells with lytic replication will go into apoptosis, while those with harboring latent virus will survive. However, the latter cells are in a hostage situation where latent virus can reactivate and cause de novo infection and initiate renewed spread. Our results suggest that EHV-1 is present at multiple sites throughout the body, in an arrested and very restricted, quiet phase of infection. However, it is still possible that EHV-1 is still replicating at a very low level at 70 dpi, and that the replicative pace may differ between sites. The simultaneous presence of a lymphocytic/monocytic infiltrate in basically most of the neural samples in our study could be an indicator of an actively involved immune system to clear infection at these sites and shows the limitation in our observations to a single p.i. time point. However, the wide-spread distribution pattern in the horses in this study highlights the complexity of EHV-1’s chronic-persistent infection and suggests that a onetime limited tissue analysis close to primary sites of replication is probably insufficient for determination of a true latency status diagnosis in an individual horse.
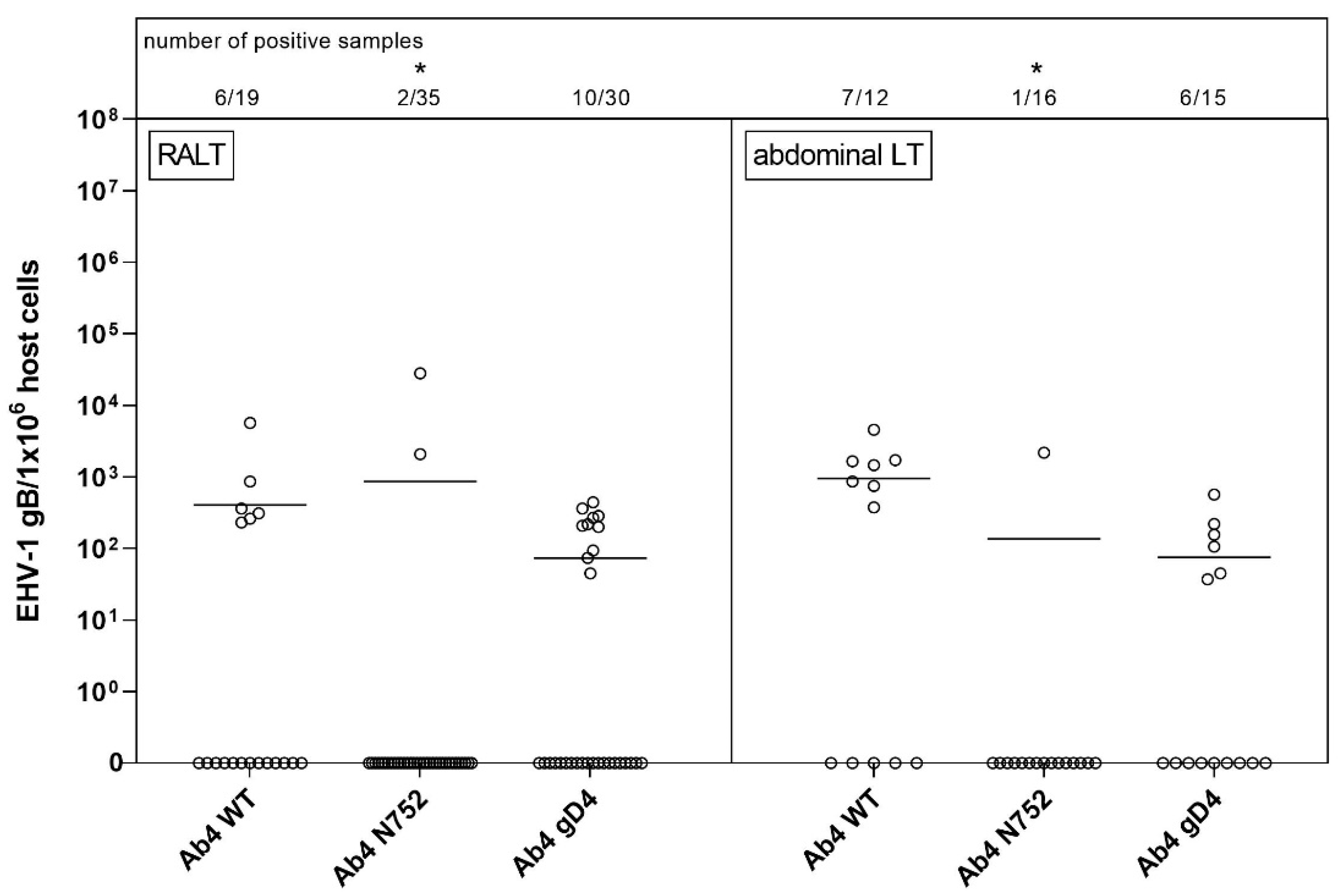
Comparison between groups was used to determine the presence of tissue preferences among EHV-1 Ab4 WT and the two mutants with assumed differing neuropathogenic potential and significant differences in viremia. Interestingly, we showed a clear difference in the distribution of EHV-1 gDNA between the groups. The polymerase mutant (Ab4 N752) caused clinically significantly different disease compared to Ab4 WT and Ab4 gD4. Infection with this EHV-1 Ab4 N752 mutant caused the most severe respiratory clinical signs after infection [
16]. However, viral DNA was found in less than 10% of the analyzed RALT samples in this group. A study by Quintana et al. (2011), where the equine airway epithelium was assessed immunologically in vitro, suggested that early events in the respiratory tract may shape downstream responses and clinical outcome [
38,
39]. In addition, local virus-specific mucosal immunity is believed to represent a first line of defense against EHV-1 infection and may impede viremia [
40]. In the present study, horses infected with Ab4 N752 did not show a significant secondary fever and exhibited significantly less viremia. Nevertheless, EHV-1 gDNA was also detected in various neural tissues, suggesting that chronic-persistent infection reached these sites rather effectively. The exact mechanisms by which different sites of latency were reached, and potential cell types for which EHV-1 gDNA spread to ganglionic samples requires further investigation.
Horses infected with EHV-1 Ab4 WT showed significantly higher viremia than the other groups. Three horses in this group developed clinical signs of EHM. None of the horses in the other groups displayed signs of EHM [
16]. Despite the absence of severe clinical signs in horses infected with EHV-1 Ab4 gD4, and the possible role of gD4 in disease attenuation, no significant differences in viral genome distribution were identified between EHV-1 Ab4 gD4 and EHV-1 Ab4 WT. In both groups, viral gDNA could be detected in neural and lymphoid tissues throughout the horses’ body. Overall, it is likely that EHV-1 uses both hematogenous and neural (axonal) spread for establishment of persistence/latency but that there may be differences between viral strains and mutants, and between EHV-1 and EHV-4 in whether hematogenous or neural spread is the preferred route.
Some of the limitations of the study are the lack of a control group, which was not included due to ethical reasons. In addition, parallel infection of all three groups would have been ideal to provide the exact same experimental conditions, but implementation was not possible due to human resources and available facilities. Sample collection of the correct part of the DRG nerve root in a time sensitive containment of necropsy proved to be difficult due to rather disperse nature of DRG in equids. To increase neuroanatomical accuracy in future, sampling protocols have been amended. Most importantly, our results represent a single time point (70 dpi), which, ideally should be compared to one or more even later time points p.i. (months/year), although this is highly impracticable.
4. Materials and Methods
4.1. Animals
A total of 25 Western Stock yearling horses (14 colts and 11 fillies, age: 10–15 months) were infected on three separate occasions as described elsewhere between June 2014 and Mai 2015 [
16,
41]. The study was approved by the institutional animal care and use committee of Michigan State University, MI, USA, and details of this study are published by Holz et al. 2017 and 2019 [
16,
41].
4.2. Experimental Design
A total of 25 Western Stock yearling horses were divided into three groups with 8, 9 and 8 subjects per group. All horses had serum neutralizing antibody titers against EHV-1 and EHV-4 of less than 1:4 and 1:40, respectively. Horses were infected via pharyngeal instillation of 5 × 10
7 TCID50/mL with either wild type EHV-1, neuropathogenic strain Ab4 suspension (group Ab4 WT; n = 8), an Ab4 mutant with aspartate (D) substituted by asparagine (N) at position 752 (Ab4 N752; n = 9), or an Ab4 mutant where the EHV-1 glycoprotein D (gD) nucleotide sequence was replaced by EHV-4 gD equivalent (Ab4 gD4; n = 8) [
9,
12,
13].
At Day 70 (±3 days) p.i. venous blood was collected in heparinized tubes following sedation with detomidine (0.012 mg/kg i.v.) and prior to euthanasia with an intravenous pentobarbital overdose (380 mg/kg i.v.). PBMC were isolated using ficoll hypaque gradient centrifugation and were stored in liquid nitrogen until further analysis. A postmortem examination and sample collection was carried out immediately and only two or three horses were euthanized per day to allow for a comprehensive postmortem examination and tissue sampling. All horses of a particular group (Ab4 WT; Ab4 N752; or Ab4 gD4) were processed within one week.
4.3. Tissue Collection
Tissue collected included mandibular and retropharyngeal lymph nodes, pharyngeal roof (Waldeyer’s ring) and bronchial lymph node, as representatives of RALT, as well as mesenteric lymph node and spleen. In addition to the TG, the spinal DRG of the lumbar spinal cord, as well as S/PS ganglia from the head, neck, thorax and abdomen (ciliary, cranial and caudal cervical ganglion, mesenteric/celiac ganglion and sympathetic trunk) were collected. All tissues were fixated in 10% neutral-buffered formalin and embedded in paraffin after 24–36 h of fixation and subsequent trimming.
4.4. Tissue Processing
Tissue blocks were sectioned at 4 µm thickness. Prior to sectioning the microtome was treated with RNase killer solution spray (RNase ZAP™, Sigma-Aldrich, Darmstadt, Germany). Six serial sections were cut, with the third section set aside for H&E staining. The remaining five sections were placed in a sterile, RNAse free Eppendorf tube and stored at −80 °C until further processing.
H&E stained neural tissue slides were evaluated for presence of ganglion (neuronal) cell bodies using light microscopy. Where no ganglion cells were identified, paraffin blocks were sectioned further, (six cuts with Cut #3 for light microscopy assessment) until ganglion cells were identified, or for up to a total of three attempts. Sections that did not contain ganglion cells were excluded from further analyses and are marked ‘no ganglion cells’ in
Figure 1. Where ganglion cells were present, the sections were scored by a veterinary neuropathologist (K.M.) for histopathological changes, cell degeneration and mononuclear cell infiltration, using a scoring key of no changes, mild, moderate, or severe changes (
Table S1).
4.5. Quantitative PCR (qPCR)—Genomic and Reverse Transcriptase PCR (RT-PCR)
For determination of ‘latent’ EHV-1, collected tissues were screened for both genomic viral DNA (gDNA) and viral mRNA of a transcriptional product of the viral late (L) gene as previously described [
33]. This method associates the identification of viral gDNA in the absence of transcriptional activity (absence of viral mRNA), with the confirmation of EHV-1 in its non-replicative ‘latent’ phase (chronic-persistent infection), while detection of both gDNA and L-gene mRNA is interpreted as ‘lytic’ EHV-1 being present (replication phase) [
4,
36].
Total DNA and RNA were extracted from formalin- fixed, paraffin-embedded (FFPE) sample sections using AllPrep DNA/RNA FFPE Kit (Qiagen, Hilden, Germany) following the manufacturer′s instructions. Total DNA and RNA were extracted from PBMC samples using the QIAamp DNA Blood Mini Kit and RNeasy Mini Kit, respectively, following the manufacturer′s instructions, (Qiagen, Hilden, Germany). DNA/RNA free water was included as extraction control in each extraction.
RNA extracted samples were tested to confirm absence of gDNA by real-time qPCR using the housekeeping gene equine glyceraldehyde-3-phosphate dehydrogenase (eqGAPDH) as a marker [
42]. Samples that were eqGAPDH gDNA positive were treated with the RQ1 RNase-free DNase Kit (Promega) following the manufacturer’s instructions, to remove any remaining gDNA. Once the absence of gDNA was confirmed, the RNA concentration was measured by nanodrop (NanoDrop™ Spectrophotometer ND-1000, Peqlab Biotechnologie GmbH, Erlangen, Germany). Subsequently, up to 5 µg of RNA was reverse transcribed using the QuantiNova Reverse Transcription Kit (Qiagen, Hilden, Germany) with random primers, following the manufacturer’s instructions. To assess if the reverse transcription was successful, the QuantiNova Internal Control RNA (QN IC RNA) was included in each reaction according to manufacturer′s recommendations. The cDNA was further diluted 1:5 with RNase/DNase-free water and cDNA samples were stored at −20 °C until further analysis.
Unless stated otherwise, all qPCR reactions for DNA and cDNA analysis were performed with the same thermal profile including an initial 95 °C step for 2 min, followed by 40 cycles of 95 °C for 10 s and 60 °C for 60 s (and hold 60 °C for 60 s). The qPCR was performed in a total reaction volume of 20 µL using 1 x SensiFAST™ Probe Lo-ROX Kit (Bioline, Luckenwalde, Germany) and 5 µL of the template. Amplification and detection were performed in strips of eight PCR tubes and caps (BRAND®, Wertheim, Germany) using the Stratagene Mx3000P cycler (Agilent Technologies, Waldbronn, Germany). All reactions included a non-template control (DNA/RNA-free water), an extraction control and a positive control for EHV-1 (EHV-1 gDNA extracted from EHV-1 infected RK13 cells), and an extraction control for equine tissue (DNA extracted from equine liver).
For detection of EHV-1 gDNA and of reverse transcribed L gene cDNA in tissue samples, a qPCR targeting a region of the glycoprotein B gene within the open reading frame 33 (ORF 33) was performed as previously published [
43]. Forward and reverse primers were added to a final concentration of 450 nM each, and the probe to a final concentration of 100 nM. For absolute quantitation and generation of a standard curve a tenfold dilution series of a defined EHV-1 DNA plasmid (courtesy of W. Azab and N. Osterrieder) was tested [
15]. The number of cells was estimated using beta-2-microglobulin (B2M) as a housekeeping gene extrapolated to a standard curve generated with oligonucleotides specific to equine B2M [
34]. Viral DNA and mRNA concentrations were expressed as copies per million cells, considering that each diploid eukaryotic cell contains two copies of the B2M gene [
33,
44]. All primers and probes sequences are presented in
Table 1.
4.6. EHV-1 Specific Immunohistochemistry
As a further control for translational gene activity, which would be expected in lytic infection activity, all gDNA-positive/cDNA-negative tissue sections were further assessed immunohistochemically. An EHV-1/ERV polyclonal caprine antiserum (VMRD, Pullman, WA, USA) was used to detect (presumably lytic) virus protein expression. EHV-1 gDNA PCR positive lung tissue from an EHV-1 aborted fetus was used as positive control, and PCR negative lung from archived samples was used as negative control. Sections were processed according to an established EHV-1 IHC protocol. In short: following deparaffinization, tissue sections were mounted on slides and antigen retrieval was performed with citrate buffer (0.1 M, pH 6.0) and heating in a microwave oven (700 W, 20 min). Samples were incubated with rabbit antiserum (1:10, 30 min), directly followed by incubation with the polyclonal caprine EHV-1/ERV antiserum (1:1600, 60 min) without washing. After incubation, slides were rinsed with tris buffered saline (TBS) and incubated with a secondary rabbit anti-goat biotinylated antibody (Vector Laboratories LTD, Burlingame, CA, USA) conjugated to horseradish peroxidase, incubated with avidin-biotin-complex (1:100, 30 min) and visualized by 3,3′- diaminobenzidine (DAB) (Vector laboratories LTD, Burlingame, CA, USA). Samples were rinsed with running tap water to stop detection and counterstaining was performed with filtered Mayer’s haemalaun (AppliChem GmbH, Darmstadt, Germany).
4.7. Statistical Analysis
Statistical analysis was performed using IBM-SPSS-Statistics 24.0 software (IBM Deutschland GmbH, Ehningen, Germany). Differences between viral mutant distribution pattern in tissues (the affinity to neural and/or lymphoid tissues) within each group and among groups were compared using cross tabulation and U tests for group ranked data. The affinity of the virus to a certain tissue type was defined by the frequency and the viral genomic load found in that tissue type. The correlation coefficient of tissue qPCR results and degree of lymphocytic/histiocytic influx was calculated using cross tabulation and chi-squared test. The Mann–Whitney U test was used to analyze the correlation between cell number and qPCR results. p-values < 0.05 were considered statistically significant.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}