
Epitope-Specific Response of Human Milk Immunoglobulins in COVID-19 Recovered Women

 , , , ,
, , , ,
Abstract
1. Introduction
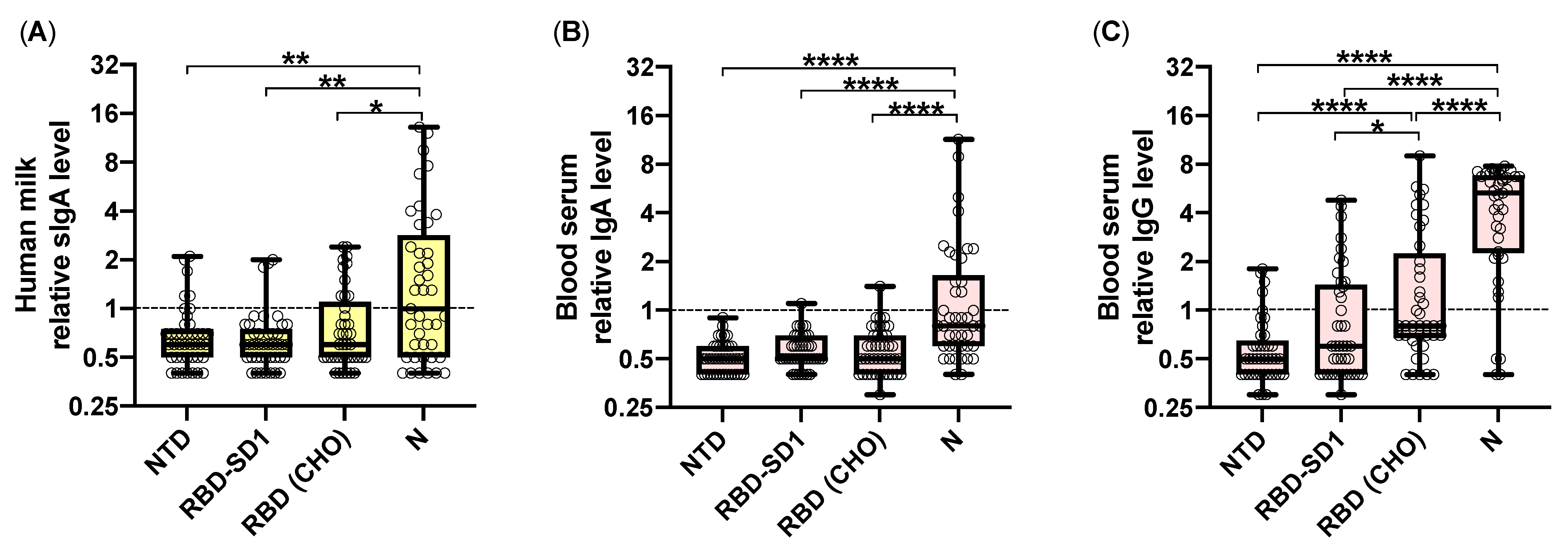
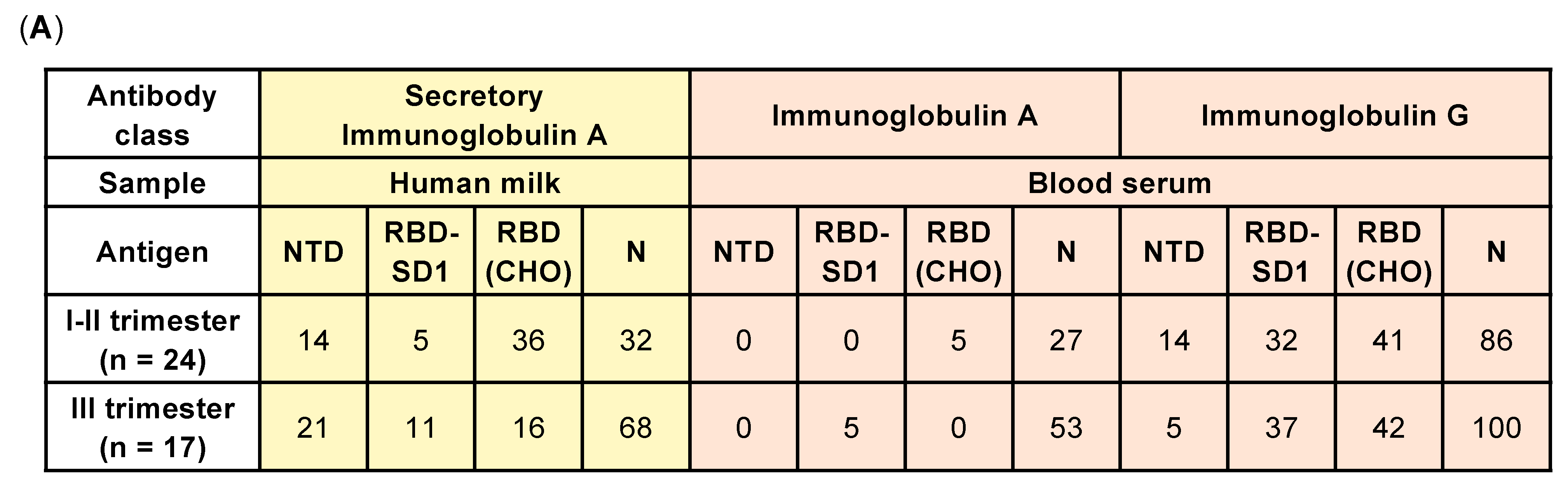
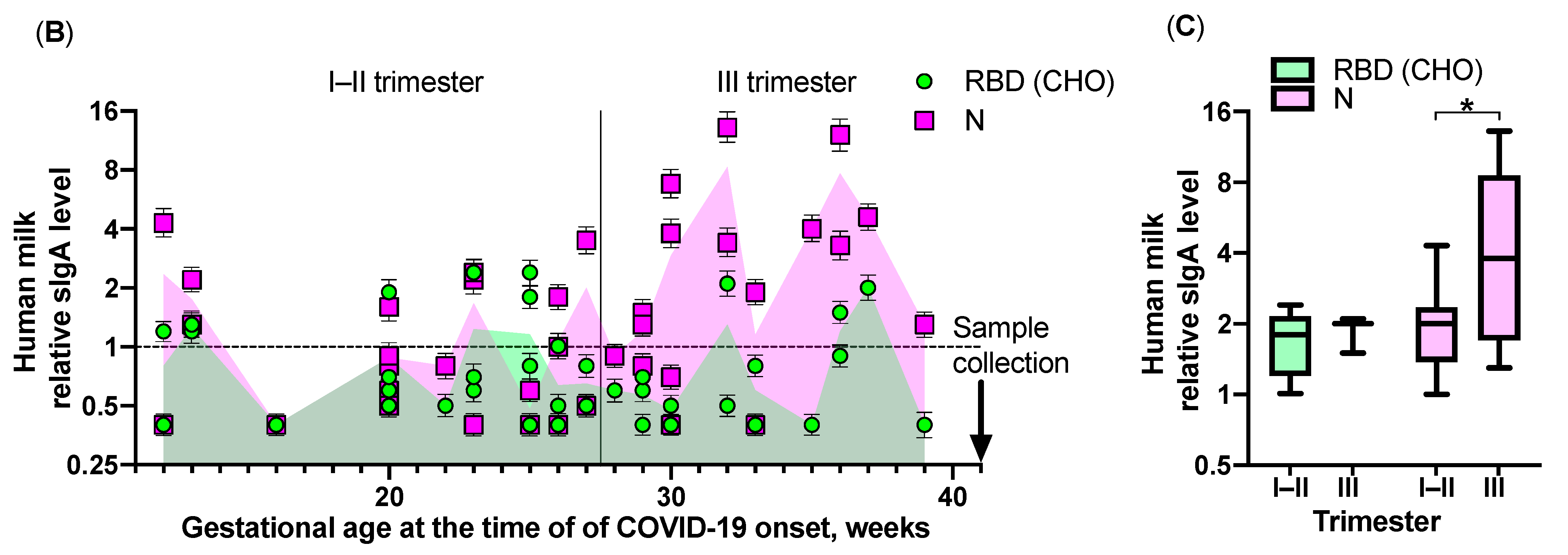
2. Results
3. Materials and Methods
3.1. Development of a Platform for Immunological Screening and Production of Recombinant Proteins and SARS-CoV-2 S- and N-Protein Fragments
3.2. ELISA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blumenthal, D.; Fowler, E.J.; Abrams, M.; Collins, S.R. Covid-19—Implications for the Health Care System. N. Engl. J. Med. 2020, 383, 1483–1488. [Google Scholar] [CrossRef]
- Aristodemou, K.; Buchhass, L.; Claringbould, D. COVID 19 crisis in the EU: The resilience of healthcare systems, government responses and their socio economic effects. Eurasian Econ. 2021. [Google Scholar] [CrossRef]
- Fox, A.; Marino, J.; Amanat, F.; Krammer, F.; Hahn-Holbrook, J.; Zolla-Pazner, S.; Powell, R.L. Robust and specific secretory IgA against SARS-CoV-2 detected in human milk. iScience 2020, 23, 101735. [Google Scholar] [CrossRef]
- Cui, X.; Zhao, Z.; Zhang, T.; Guo, W.; Guo, W.; Zheng, J.; Zhang, J.; Dong, C.; Na, R.; Zheng, L.; et al. A systematic review and meta-analysis of children with coronavirus disease 2019 (COVID-19). J. Med. Virol. 2021, 93, 1057–1069. [Google Scholar] [CrossRef] [PubMed]
- Raba, A.A.; Abobaker, A.; Elgenaidi, I.S.; Daoud, A. Novel coronavirus infection (COVID-19) in children younger than one year: A systematic review of symptoms, management and outcomes. Acta Paediatr. 2020, 109, 1948–1955. [Google Scholar] [CrossRef]
- Dong, Y.; Mo, X.; Hu, Y.; Qi, X.; Jiang, F.; Jiang, Z.; Tong, S. Epidemiological characteristics of 2143 pediatric patients with 2019 coronavirus disease in China. Pediatrics 2020, 145, e20200702. [Google Scholar] [CrossRef] [PubMed]
- Calina, D.; Docea, A.J.; Petrakis, D.; Egorov, A.V.; Ishmukhametov, A.A.; Gabibov, A.G.; Shtilman, V.I.; Kostoff, R.; Carvalho, F.; Vinceti, M.; et al. Towards effective COVID-19 vaccines: Updates, perspectives and challenges. Int. J. Mol. Med. 2020, 46, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Section of Breastfeeding. Breastfeeding and the Use of Human Milk. Pediatrics 2012, 129, e827–e841. [Google Scholar] [CrossRef] [PubMed]
- Victora, C.G.; Bahl, R.; Barros, A.J.D.; França, G.V.A.; Horton, S.; Krasevec, J.; Murch, S.; Sankar, M.J.; Walker, N.; Rollins, N.C. Breastfeeding in the 21st century: Epidemiology, mechanisms, and lifelong effect. Lancet 2016, 387, 475–490. [Google Scholar] [CrossRef]
- WHO. Clinical Management of Severe Acute Respiratory Infection (SARI) When COVID-19 Disease Is Suspected: Interim Guidance. World Health Organization. 2020. Available online: https://apps.who.int/iris/bitstream/handle/10665/331446/WHO-2019-nCoV-clinical-2020.4eng.pdf?sequence=1&isAllowed=y (accessed on 10 October 2020).
- Peng, S.; Zhu, H.; Yang, L.; Cao, L.; Huang, X.; Dynes, M.; Narayan, A.; Xia, J.; Chen, Y.; Zhang, P.; et al. A study of breastfeeding practices, SARS-CoV-2 and its antibodies in the breast milk of mothers confirmed with COVID-19. Lancet Reg. Health West. Pac. 2020, 4, 100045. [Google Scholar] [CrossRef] [PubMed]
- Parker, L.A.; Sullivan, S.; Krueger, C.; Mueller, M. Association of timing of initiation of breastmilk expression on milk volume and timing of lactogenesis stage II among mothers of very low-birth-weight infants. Breastfeed. Med. 2015, 10, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Edmond, K.M.; Zandoh, C.; Quigley, M.A.; Amenga-Etego, S.; Owusu-Agyei, S.; Kirkwood, B.R. Delayed breastfeeding initiation increases risk of neonatal mortality. Pediatrics 2006, 117, 380–386. [Google Scholar] [CrossRef]
- Groß, R.; Conzelmann, C.; Müller, J.A.; Stenger, S.; Steinhart, K.; Kirchhoff, F.; Münch, J. Detection of SARS-CoV-2 in human breastmilk. Lancet 2020, 395, 1757–1758. [Google Scholar] [CrossRef]
- Kirtsman, M.; Diambomba, Y.; Poutanen, S.M.; Malinowski, A.K.; Vlachodimitropoulou, E.; Parks, W.T.; Erdman, L.; Morris, S.K.; Shah, P.S. Probable congenital SARSCoV-2 infection in a neonate born to a woman with active SARS-CoV-2 infection. CMAJ Can. Med. Assoc. J. 2020, 192, 647–650. [Google Scholar] [CrossRef]
- Tam, P.C.K.; Ly, K.M.; Kernich, M.L.; Spurrier, N.; Lawrence, D.; Gordon, D.L.; Tucker, E.C. Detectable severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) in human breast milk of a mildly symptomatic patient with coronavirus disease 2019 (COVID-19). Clin. Infect. Dis 2020, 72, 128–130. [Google Scholar] [CrossRef]
- Chen, H.; Guo, J.; Wang, C.; Luo, F.; Yu, X.; Zhang, W.; Li, J.; Zhao, D.; Xu, D.; Gong, Q.; et al. Clinical characteristics and intrauterine vertical transmission potential of COVID-19 infection in nine pregnant women: A retrospective review of medical records. Lancet 2020, 395, 809–815. [Google Scholar] [CrossRef]
- Fan, C.; Lei, D.; Fang, C.; Li, C.; Wang, M.; Liu, Y.; Bao, Y.; Sun, Y.; Huang, J.; Guo, Y.; et al. Perinatal transmission of COVID-19 associated SARSCoV-2: Should we worry? Clin. Infect. Dis 2020, 72, 862–864. [Google Scholar] [CrossRef]
- Pace, R.M.; Williams, J.E.; Jarvinen, K.M.; Belfort, M.B.; Pace, C.D.W.; Lackey, K.A.; Gogel, A.C.; Nguyen-Contant, P.; Kanagaiah, P.; Fitzgerald, T.; et al. Characterization of SARS-CoV-2 RNA, Antibodies, and Neutralizing Capacity in Milk Produced by Women with COVID-19. mBio 2021, 12, e03192-20. [Google Scholar] [CrossRef] [PubMed]
- Ackerman, D.L.; Craft, K.M.; Doster, R.S.; Weitkamp, J.-H.; Aronoff, D.M.; Gaddy, J.A.; Townsend, S.D. Antimicrobial and Antibiofilm Activity of Human Milk Oligosaccharides against Streptococcus agalactiae, Staphylococcus aureus, and Acinetobacter baumannii. ACS Infect. Dis. 2018, 4, 315–324. [Google Scholar] [CrossRef]
- Quitadamo, P.A.; Comegna, L.; Cristalli, P. Anti-Infective, Anti-Inflammatory, and Immunomodulatory Properties of Breast Milk Factors for the Protection of Infants in the Pandemic From COVID-19. Front. Public Health 2021, 8, 589736. [Google Scholar] [CrossRef] [PubMed]
- Demers-Mathieu, V.; DaPra, C.; Mathijssen, G.; Sela, D.A.; Järvinen, K.M.; Seppo, A.; Fels, S.; Medo, E. Human milk antibodies against S1 and S2 Subunits from SARS-CoV-2, HCoV-OC43, and HCoV-229E in mothers with a confirmed COVID-19 PCR, viral symptoms, and unexposed mothers. Int. J. Mol. Sci. 2021, 22, 1749. [Google Scholar] [CrossRef] [PubMed]
- Bobik, T.V.; Kostin, N.N.; Skryabin, G.A.; Tsabai, P.N.; Simonova, M.A.; Knorre, V.D.; Stratienko, O.N.; Aleshenko, N.L.; Vorobiev, I.I.; Khurs, E.N.; et al. COVID-19 in Russia: Clinical and Immunological Features of the First-Wave Patients. Acta Nat. 2021, 48, 102–115. [Google Scholar] [CrossRef]
- Burbelo, P.D.; Riedo, F.X.; Morishima, C.; Rawlings, S.; Smith, D.; Das, S.; Strich, J.R.; Chertow, D.S.; Davey, R.T.; Cohen, J. Sensitivity in Detection of Antibodies to Nucleocapsid and Spike Proteins of Severe Acute Respiratory Syndrome Coronavirus 2 in Patients With Coronavirus Disease. J. Infect. Dis. 2020, 222, 206–213. [Google Scholar] [CrossRef]
- Durova, O.M.; Vorobiev, I.I.; Smirnov, I.V.; Reshetnyak, A.V.; Telegin, G.B.; Shamborant, O.G.; Orlova, N.A.; Genkin, D.D.; Bacon, A.; Ponomarenko, N.A.; et al. Strategies for induction of catalytic antibodies toward HIV-1 glycoprotein gp120 in autoimmune prone mice. Mol. Immunol. 2009, 47, 87–95. [Google Scholar] [CrossRef]
- Sinegubova, M.V.; Orlova, N.A.; Kovnir, S.V.; Dayanova, L.K.; Vorobiev, I.I. High-level expression of the monomeric SARS-CoV-2 S protein RBD 320-537 in stably transfected CHO cells by the EEF1A1-based plasmid vector. PLoS ONE 2021, 16, e0242890. [Google Scholar] [CrossRef]
- Stepanov, A.V.; Belogurov, A.A., Jr.; Ponomarenko, N.A.; Stremovskiy, O.A.; Kozlov, L.V.; Bichucher, A.M.; Dmitriev, S.E.; Smirnov, I.V.; Shamborant, O.G.; Balabashin, D.S.; et al. Design of targeted B cell killing agents. PLoS ONE 2011, 6, e20991. [Google Scholar] [CrossRef]
- Joyner, M.J.; Carter, R.E.; Senefeld, J.W.; Klassen, S.A.; Mills, J.R.; Johnson, P.W.; Theel, E.S.; Wiggins, C.C.; Bruno, K.A.; Klompas, A.M.; et al. Convalescent Plasma Antibody Levels and the Risk of Death from Covid-19. N. Engl. J. Med. 2021, 11, 1015–1027. [Google Scholar] [CrossRef] [PubMed]
- Rappazzo, C.G.; Tse, L.V.; Kaku, C.I.; Wrapp, D.; Sakharkar, M.; Huang, D.; Deveau, L.M.; Yockachonis, T.J.; Herbert, A.S.; Battles, M.B.; et al. Broad and potent activity against SARS-like viruses by an engineered human monoclonal antibody. Science 2021, 371, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Huang, L.; Zhang, G.; Yao, Y.; Zhou, H.; Shen, S.; Shen, B.; Li, B.; Li, X.; Chen, M.; et al. A SARS-CoV-2 neutralizing antibody with exceptional spike binding coverage and optimized therapeutic potentials. Nat. Commun. 2021, in press. [Google Scholar] [CrossRef]
- Weinreich, D.M.; Sivapalasingam, S.; Norton, T.; Ali, S.; Gao, H.; Bhore, R.; Musser, B.J.; Soo, Y.; Rofail, D.; Im, J.; et al. REGN-COV2, A Neutralizing Antibody Cocktail, in Outpatients with Covid-19. N. Engl. J. Med. 2021, 384, 238–251. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liu, Z.; Li, S.; Xu, W.; Zhang, Q.; Silva, I.T.; Li, C.; Wu, Y.; Jiang, Q.; Liu, Z.; et al. Enhancement versus neutralization by SARS-CoV-2 antibodies from a convalescent donor associates with distinct epitopes on the RBD. Cell Rep. 2021, 34, 108699. [Google Scholar] [CrossRef] [PubMed]
- Cabinian, A.; Sinsimer, D.; Tang, M.; Zumba, O.; Mehta, H.; Toma, A.; Sant’Angelo, D.; Laouar, Y.; Laouar, A. Transfer of Maternal Immune Cells by Breastfeeding: Maternal Cytotoxic T Lymphocytes Present in Breast Milk Localize in the Peyer’s Patches of the Nursed Infant. PLoS ONE 2016, 11, e0156762. [Google Scholar] [CrossRef] [PubMed]
- Laouar, A. Maternal Leukocytes and Infant Immune Programming during Breastfeeding. Trends Immunol. 2020, 41, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Ziganshin, R.H.; Ivanova, O.M.; Lomakin, Y.A.; Belogurov, A.A., Jr.; Kovalchuk, S.I.; Azarkin, I.V.; Arapidi, G.P.; Anikanov, N.A.; Shender, V.O.; Piradov, M.A.; et al. The Pathogenesis of the Demyelinating Form of Guillain-Barre Syndrome (GBS): Proteo-peptidomic and Immunological Profiling of Physiological Fluids. Mol. Cell. Proteom. 2016, 7, 2366–2378. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Indicator | Group 1 (Trimester I of Pregnancy) n = 3 | Group 2 (Trimester II of Pregnancy) n = 21 | Group 3 (Trimester III of Pregnancy) n = 17 | p-Value |
|---|---|---|---|---|
| Age, years (median (min; max)) | 27 (24; 36) | 33 (23; 42) | 29 (24; 41) | p = 0.159 |
| Mild COVID-19 symptoms, n (%) | 2 (67%) | 16 (76%) | 12 (71%) | p = 0.779 |
| Moderate COVID-19 symptoms, n (%) | 0 (0%) | 4 (19%) | 4 (23%) | p = 0.191 |
| Severe COVID-19 symptoms, n (%) | 1 (33%) | 1 (5%) | 1 (6%) | p = 0.011 |
| Premature birth, n (%) | 1 (33%) multiple pregnancy | 2 (9%) 1—ingrowth of the placenta; 1—intrauterine infection, generally contracted pelvis, 2nd degree | 3 (18%) 1—ingrowth of the placenta 2—delivery | p = 0.134 |
| Birth at term, n (%) | 2 (67%) | 19 (91%) | 14 (82%) | p = 0.771 |
| Newborn body weight, g, median (min; max) | 2966 (2220; 3878) | 3444 (1800; 4222) | 3540 (2770; 4340) | p = 0.709 |
| Newborn body length, cm, median (min; max) | 49.0 (44.0; 55.0) | 53.0 (41.0; 58.0) | 53.0 (49.0; 56.0) | p = 0.300 |
| Apgar score at 1 min after birth, median (min; max) | 7.5 (4; 8) | 8 (6; 8) | 8 (7; 8) | p = 0.040 |
| Apgar score at 5 min after birth, median (min; max) | 8 (8; 9) | 9 (7; 9) | 9 (8; 9) | p = 0.090 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bobik, T.V.; Kostin, N.N.; Skryabin, G.A.; Tsabai, P.N.; Simonova, M.A.; Knorre, V.D.; Mokrushina, Y.A.; Smirnov, I.V.; Kosolapova, J.A.; Vtorushina, V.V.; et al. Epitope-Specific Response of Human Milk Immunoglobulins in COVID-19 Recovered Women. Pathogens 2021, 10, 705. https://doi.org/10.3390/pathogens10060705
Bobik TV, Kostin NN, Skryabin GA, Tsabai PN, Simonova MA, Knorre VD, Mokrushina YA, Smirnov IV, Kosolapova JA, Vtorushina VV, et al. Epitope-Specific Response of Human Milk Immunoglobulins in COVID-19 Recovered Women. Pathogens. 2021; 10(6):705. https://doi.org/10.3390/pathogens10060705
Chicago/Turabian StyleBobik, Tatyana V., Nikita N. Kostin, George A. Skryabin, Polina N. Tsabai, Maria A. Simonova, Vera D. Knorre, Yuliana A. Mokrushina, Ivan V. Smirnov, Julia A. Kosolapova, Valentina V. Vtorushina, and et al. 2021. "Epitope-Specific Response of Human Milk Immunoglobulins in COVID-19 Recovered Women" Pathogens 10, no. 6: 705. https://doi.org/10.3390/pathogens10060705
APA StyleBobik, T. V., Kostin, N. N., Skryabin, G. A., Tsabai, P. N., Simonova, M. A., Knorre, V. D., Mokrushina, Y. A., Smirnov, I. V., Kosolapova, J. A., Vtorushina, V. V., Inviyaeva, E. V., Polushkina, E., Petrova, U. L., Levadnaya, A. V., Krechetova, L. V., Shmakov, R. G., Sukhikh, G. T., & Gabibov, A. G. (2021). Epitope-Specific Response of Human Milk Immunoglobulins in COVID-19 Recovered Women. Pathogens, 10(6), 705. https://doi.org/10.3390/pathogens10060705