
Antibodies to Highly Pathogenic A/H5Nx (Clade 2.3.4.4) Influenza Viruses in the Sera of Vietnamese Residents

Abstract
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
Funding
Author Contributions
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Krammer, F.; Smith, G.J.D.; Fouchier, R.A.M.; Peiris, M.; Kedzierska, K.; Doherty, P.C.; Palese, P.; Shaw, M.L.; Treanor, J.; Webster, R.G.; et al. Influenza. Nat. Rev. Dis. Primers 2018, 4, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Russell, C.J.; Hu, M.; Okda, F.A. Influenza Hemagglutinin Protein Stability, Activation, and Pandemic Risk. Trends Microbiol. 2018, 26, 841–853. [Google Scholar] [CrossRef]
- Tong, S.; Li, Y.; Rivailler, P.; Conrardy, C.; Castillo, D.A.; Chen, L.M.; Recuenco, S.; Ellison, J.A.; Davis, C.T.; York, I.A.; et al. A distinct lineage of influenza A virus from bats. Proc. Natl. Acad. Sci. USA 2012, 109, 4269–4274. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Zhu, X.; Li, Y.; Shi, M.; Zhang, J.; Bourgeois, M.; Yang, H.; Chen, X.; Recuenco, S.; Gomez, J.; et al. New world bats harbor diverse influenza A viruses. PLoS Pathog. 2013, 9, e1003657. [Google Scholar] [CrossRef]
- Hay, A.J.; Gregory, V.; Douglas, A.R.; Yi, P.L. The evolution of human influenza viruses. Philos. Trans. R. Soc. B Biol. Sci. 2001, 356, 1861–1870. [Google Scholar] [CrossRef]
- Short, K.R.; Kedzierska, K.; van de Sandt, C.E. Back to the Future: Lessons Learned From the 1918 Influenza Pandemic. Front. Cell. Infect. Microbiol. 2018, 8, 343. [Google Scholar] [CrossRef]
- Scholtissek, C.; Rohde, W.; Von Hoyningen, V.; Rott, R. On the origin of the human influenza virus subtypes H2N2 and H3N2. Virology 1978, 87, 13–20. [Google Scholar] [CrossRef]
- Smith, G.J.D.; Vijaykrishna, D.; Bahl, J.; Lycett, S.J.; Worobey, M.; Pybus, O.G.; Ma, S.K.; Cheung, C.L.; Raghwani, J.; Bhatt, S.; et al. Origins and evolutionary genomics of the 2009 swineorigin H1N1 influenza A epidemic. Nature 2009, 459, 1122–1125. [Google Scholar] [CrossRef]
- Worobey, M.; Han, G.Z.; Rambaut, A. Genesis and pathogenesis of the 1918 pandemic H1N1 influenza A virus. Proc. Natl. Acad. Sci. USA 2014, 111, 8107–8112. [Google Scholar] [CrossRef]
- Morens, D.M.; Taubenberger, J.K. Influenza Cataclysm, 1918. N. Engl. J. Med. 2018, 379, 2285–2287. [Google Scholar] [CrossRef]
- WHO H5N1. Cumulative Number of Confirmed Human Cases of Avian Influenza A(H5N1) Reported to WHO. Available online: http://www.who.int/influenza/human_animal_interface/H5N1_cumulative_table_archives/en/ (accessed on 11 March 2021).
- Tsunekuni, R.; Sudo, K.; Nguyen, P.T.; Luu, B.D.; Phuong, T.D.; Tan, T.M.; Nguyen, T.; Mine, J.; Nakayama, M.; Tanikawa, T.; et al. Isolation of highly pathogenic H5N6 avian influenza virus in Southern Vietnam with genetic similarity to those infecting humans in China. Transbound. Emerg. Dis. 2019, 66, 2209–2217. [Google Scholar] [CrossRef] [PubMed]
- Thanh, H.D.; Tran, V.T.; Nguyen, D.T.; Hung, V.K.; Kim, W. Novel reassortant H5N6 highly pathogenic influenza A viruses in Vietnamese quail outbreaks. Comp. Immunol. Microbiol. Infect. Dis. 2018, 56, 45–57. [Google Scholar] [CrossRef]
- Bi, Y.; Chen, Q.; Wang, Q.; Chen, J.; Jin, T.; Wong, G.; Quan, C.; Liu, J.; Wu, J.; Yin, R.; et al. Genesis, Evolution and Prevalence of H5N6 Avian Influenza Viruses in China. Cell Host Microbe 2016, 20, 810–821. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Liu, H.; Xiong, C.; Di, L.; Shi, W.; Li, M.; Liu, S.; Chen, J.; Chen, G.; Li, Y.; et al. Novel avian influenza A (H5N6) viruses isolated in migratory waterfowl before the first human case reported in China, 2014. Sci. Rep. 2016, 6, 29888. [Google Scholar] [CrossRef]
- Takemae, N.; Tsunekuni, R.; Sharshov, K.; Tanikawa, T.; Uchida, Y.; Ito, H.; Soda, K.; Usui, T.; Sobolev, I.; Shestopalov, A.; et al. Five distinct reassortants of H5N6 highly pathogenic avian influenza A viruses affected Japan during the winter of 2016–2017. Virology 2017, 512, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhu, W.; Li, X.; Bo, H.; Zhang, Y.; Zou, S.; Gao, R.; Dong, J.; Zhao, X.; Chen, W.; et al. Genesis and Dissemination of Highly Pathogenic H5N6 Avian Influenza Viruses. J. Virol. 2017, 91. [Google Scholar] [CrossRef]
- Yang, H.; Carney, P.J.; Mishin, V.P.; Guo, Z.; Chang, J.C.; Wentworth, D.E.; Gubareva, L.V.; Stevens, J. Molecular Characterizations of Surface Proteins Hemagglutinin and Neuraminidase from Recent H5Nx Avian Influenza Viruses. J. Virol. 2016, 90, 5770–5784. [Google Scholar] [CrossRef]
- Dhingra, M.S.; Artois, J.; Robinson, T.P.; Linard, C.; Chaiban, C.; Xenarios, I.; Engler, R.; Liechti, R.; Kuznetsov, D.; Xiao, X.; et al. Global mapping of highly pathogenic avian influenza H5N1 and H5N× clade 2.3.4.4 viruses with spatial cross-validation. eLife 2016, 5, e19571. [Google Scholar] [CrossRef]
- Adlhoch, C.; Fusaro, A.; Kuiken, T.; Niqueux, É.; Terregino, C.; Staubach, C.; Muñoz Guajardo, I.; Baldinelli, F. Scientific report: Avian influenza overview February–May 2020. EFSA J. 2020, 18, e06194. [Google Scholar] [CrossRef]
- World Organization for Animal Health (OIE). Update on Highly Pathogenic Avian Influenza in Animals (Type h5 and h7); OIE: Paris, France, 2014; Available online: http://www.oie.int/en/animal-health-in-the-world/update-on-avian-influenza/2018/ (accessed on 11 March 2021).
- Kang, Y.; Liu, L.; Feng, M.; Yuan, R.; Huang, C.; Tan, Y.; Gao, P.; Xiang, D.; Zhao, X.; Li, Y.; et al. Highly pathogenic H5N6 influenza A viruses recovered from wild birds in Guangdong, southern China, 2014–2015. Sci. Rep. 2017, 7, 44410. [Google Scholar] [CrossRef]
- Schrauwen, E.J.; Fouchier, R.A. Host adaptation and transmission of influenza A viruses in mammals. Emerg. Microbes Infect. 2014, 3, e9. [Google Scholar] [CrossRef]
- FAO H7N9. Food and Agriculture Organization of the United Nations. H7N9 Situation Update. Available online: www.fao.org/ag/againfo/programmes/en/empres/H7N9/situation_update.html (accessed on 11 March 2021).
- Sutton, T.C. The Pandemic Threat of emerging H5 and H7 avian influenza viruses. Viruses 2018, 10, 461. [Google Scholar] [CrossRef]
- Caron, A.; Morand, S.; Garine-Wichatitsky, M.D. Epidemiological interaction at the wildlife/livestock/human interface: Can we anticipate emerging infectious diseases in their hotspots? A framework for understanding emerging diseases processes in their hot spots. New Front. Mol. Epi Infect. Dis. 2012, 2012, 311–332. [Google Scholar]
- Ilyicheva, T.N.; Durymanov, A.G.; Svyatchenko, S.V.; Marchenko, V.Y.; Sobolev, I.A.; Bakulina, A.Y.; Goncharova, N.I.; Kolosova, N.P.; Susloparov, I.M.; Pyankova, O.G.; et al. Humoral immunity to influenza in an at-risk population and severe influenza cases in Russia in 2016–2017. Arch. Virol. 2018, 163, 2675–2685. [Google Scholar] [CrossRef] [PubMed]
- Hoa, L.N.M.; Tuan, N.A.; My, P.H.; Huong, T.T.K.; Chi, N.T.Y.; Hau, T.T.T.; Carrique-Mas, J.; Duong, M.T.; Tho, N.D.; Hoang, N.D.; et al. Assessing evidence for avian-to-human transmission of influenza A/H9N2 virus in rural farming communities in northern Vietnam. J. Gen. Virol. 2017, 98, 2011–2016. [Google Scholar] [CrossRef] [PubMed]
- Bui, V.N.; Nguyen, T.T.; Nguyen-Viet, H.; Bui, A.N.; McCallion, K.A.; Lee, H.S.; Than, S.T.; Coleman, K.K.; Gray, G.C. Bioaerosol Sampling to Detect Avian Influenza Virus in Hanoi’s Largest Live Poultry Market. Clin. Infect. Dis. 2018. [Google Scholar] [CrossRef]
- Le, T.; Phan, L.; Nguyen, L.; Nguyen, H.; Ly, K.; Ho, T.N.; Trinh, T.; Nguyen, T.M. Fatal Avian Influenza A/H5N1 Infection in a 36-Week Pregnant Woman Survived by her Newborn—Soc Trang, Vietnam, 2012. Influenza Other Respir Viruses 2018. [Google Scholar] [CrossRef] [PubMed]
- Thuy, D.M.; Peacock, T.P.; Bich, V.T.N.; Fabrizio, T.; Hoang, D.N.; Tho, N.D.; Diep, N.T.; Nguyen, M.; Hoa, L.N.M.; Trang, H.T.T.; et al. Prevalence and diversity of H9N2 avian influenza in chickens of Northern Vietnam, 2014. Infect. Genet. Evol. 2016, 44, 530–540. [Google Scholar] [CrossRef]
- Mostafa, A.; Abdelwhab, E.M.; Mettenleiter, T.C.; Pleschka, S. Zoonotic potential of influenza A viruses: A comprehensive overview. Viruses 2018, 10, 497. [Google Scholar] [CrossRef]
- Lee, A.C.Y.; Zhu, H.; Zhang, A.J.X.; Li, C.; Wang, P.; Li, C.; Chen, H.; Hung, I.F.N.; To, K.K.W.; Yuen, K.-Y. Suboptimal humoral immune response against influenza A(H7N9) virus is related to its internal genes. Clin. Vaccine Immunol. 2015, 22, 1235–1243. [Google Scholar] [CrossRef]
- Kayali, G.; Setterquist, S.F.; Capuano, A.W.; Myers, K.P.; Gill, J.S.; Gray, G.C. Testing human sera for antibodies against avian influenza viruses: Horse RBC hemagglutination inhibition vs. microneutralization assays. J. Clin. Virol. 2008, 43, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Morens, D.V.; Daszak, P.; Taubenberger, J.K. Escaping Pandora’s Box—Another Novel Coronavirus. N. Engl. J. Med. 2020, 382, 1293–1295. [Google Scholar] [CrossRef] [PubMed]
- Taubenberger, J.K.; Kash, J.C.; Morens, D.M. The 1918 influenza pandemic: 100 years of questions answered and unanswered. Sci. Transl. Med. 2019, 11, eaau5485. [Google Scholar] [CrossRef]
- Thông Tin Về Tình Hình Dịch Cúm Gia Cầm, LMLM Và Tai Xanh Ngày 20/02/2017. Available online: http://www.cucthuy.gov.vn/Pages/thong-tin-ve-tinh-hinh-dich-cum-gia-cam-lmlm-va-tai-xanh-ngay-20-02-2017.aspx (accessed on 5 March 2021).
- Rowe, T.; Abernathy, R.A.; Hu-Primmer, J.; Thompson, W.W.; Lu, X.; Lim, W.; Fukuda, K.; Cox, N.J.; Katz, J.M. Detection of antibody to avian influenza A (H5N1) virus in human serum by using a combination of serologic assays. J. Clin. Microbiol. 1999, 37, 937–943. [Google Scholar] [CrossRef] [PubMed]
- Marchenko, V.; Goncharova, N.; Susloparov, I.; Kolosova, N.; Gudymo, A.; Svyatchenko, S.; Danilenko, A.; Durymanov, A.; Gavrilova, E.; Maksyutov, R.; et al. Isolation and characterization of H5Nx highly pathogenic avian influenza viruses of clade 2.3.4.4 in Russia. Virology 2018, 525, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Susloparov, I.M.; Goncharova, N.; Kolosova, N.; Danilenko, A.; Marchenko, V.; Onkhonova, G.; Evseenko, V.; Gavrilova, E.; Maksutov, R.A.; Ryzhikov, A. Genetic Characterization of Avian Influenza A(H5N6) Virus Clade 2.3.4.4, Russia, 2018. Emerg. Infect. Dis. 2019, 25, 2338–2339. [Google Scholar] [CrossRef]
- WHO. World Health Organization Surveillance Network: Manual for the Laboratory Diagnosis and Virological Surveillance of Influenza; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]


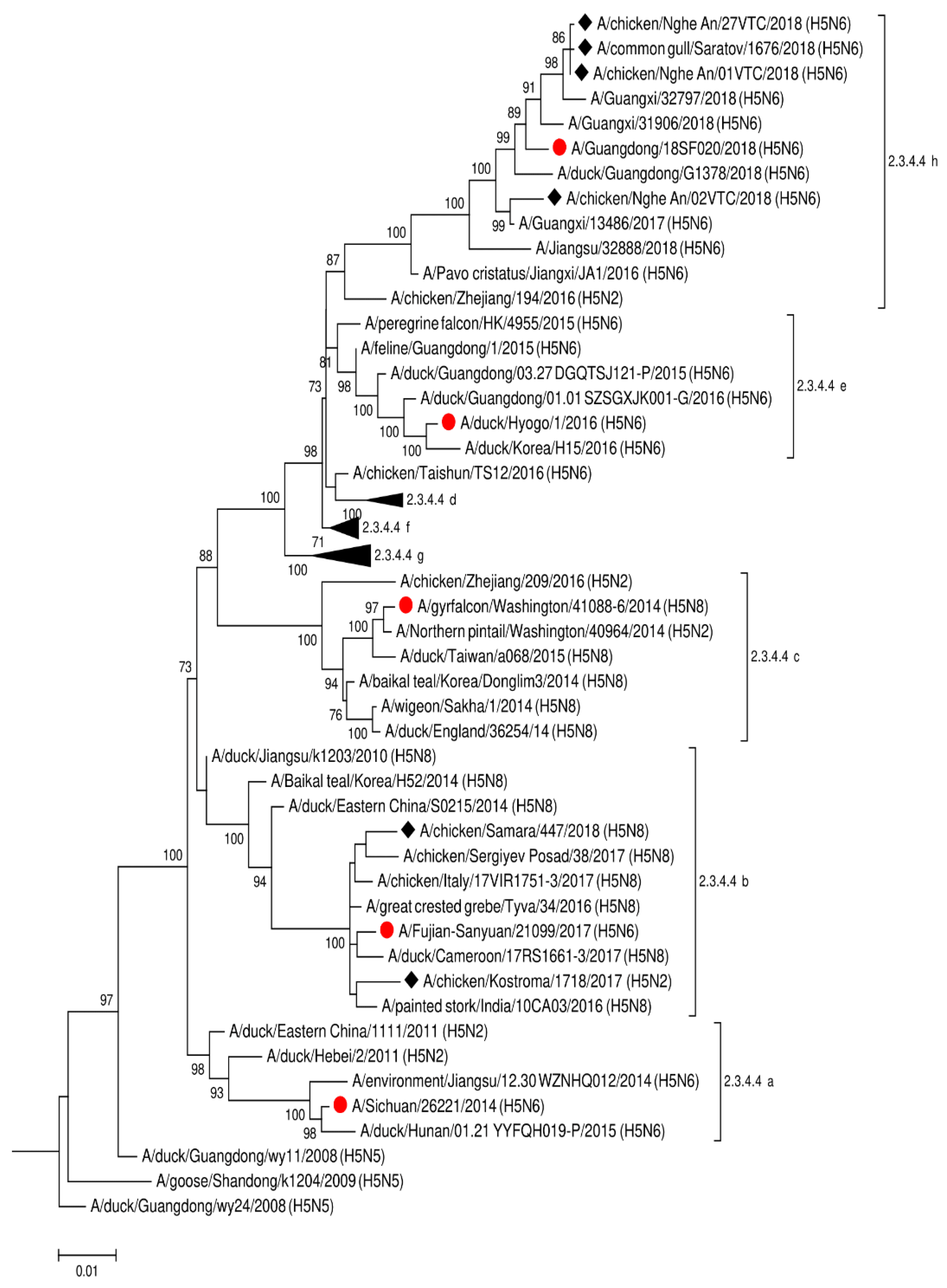{kind=link}
| Virus | Subtype | Clade | Reverse Titer with Antisera | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A/duck/England/36254/ 2014 | A/Northern Pintail/WA/40964/2014 | A/Sichuan/26221/2014 RG42A | A/gyrfalcon/WA/41088/2014 RG43A | A/great crested grebe/Tyva/34/2016 | A/wigeon/Sakha/1/2014 | A/chicken/SergiyevPosad/ 38/2017 | A/chicken/Kostroma/1718/ 2017 | A/common gull/Saratov/1676/2018 | A/chicken/Nghe An/27VTC/2018 | |||
| A/duck/England/36254/2014 | H5N8 | 2.3.4.4c | 640 | 5120 | 5120 | 640 | 1280 | 1280 | 1280 | 2560 | NA | NA |
| A/Northern Pintail/WA/40964/2014 | H5N2 | 2.3.4.4c | 320 | 5120 | 2560 | 2560 | 640 | 640 | 160 | 1280 | <20 | <20 |
| A/Sichuan/26221/2014 RG42A | H5N6 | 2.3.4.4a | 640 | 5120 | 5120 | 320 | 640 | 320 | 320 | 1280 | <20 | NA |
| A/gyrfalcon/WA/41088/2014 RG43A | H5N8 | 2.3.4.4c | 640 | 5120 | 1280 | 5120 | 1280 | 1280 | 320 | 1280 | <20 | <20 |
| A/great crested grebe/Tyva/34/2016 | H5N8 | 2.3.4.4b | 320 | 5120 | 2560 | 2560 | 1280 | 640 | 640 | 1280 | <20 | <20 |
| A/wigeon/Sakha/1/2014 | H5N8 | 2.3.4.4c | 640 | 10,240 | 2560 | 5120 | 640 | 640 | 640 | 2560 | <20 | <20 |
| A/chicken/SergiyevPosad/38/2017 | H5N8 | 2.3.4.4b | 160 | 2560 | 2560 | 2560 | 320 | 640 | 160 | 1280 | <20 | <20 |
| A/chicken/Kostroma/1718/2017 | H5N2 | 2.3.4.4b | 320 | 10,240 | 2560 | 2560 | 640 | 640 | 320 | 5120 | <20 | <20 |
| A/common gull/Saratov/1676/2018 | H5N6 | 2.3.4.4h | <20 | <20 | 80 | <20 | <20 | <20 | <20 | <20 | 320 | 80 |
| A/chicken/NgheAn/27VTC/2018 | H5N6 | 2.3.4.4h | <20 | 320 | 40 | 20 | <20 | <20 | <20 | <20 | 320 | 80 |
| A/chicken/NgheAn/01VTC/2018 | H5N6 | 2.3.4.4h | NA | 640 | 80 | <20 | <20 | <20 | <20 | <20 | 320 | 160 |
| A/chicken/Vietnam/NCVD-15A59/2015 | H5N6 | 2.3.4.4f | 320 | 10,240 | 2560 | 2560 | 640 | 640 | 160 | 640 | 40 | <20 |
| Human Serum Sample | Group | A/chicken/ NgheAn/01VTC/ 2018 (H5N6) 2.3.4.4 | A/chicken/ Kostroma/ 1718/ 2017 (H5N2) 2.3.4.4 | A/chicken/ NgheAn/27VTC/2018 (H5N6) 2.3.4.4 | A/common Gull/Saratov/1676/2018 (H5N6) 2.3.4.4 | ||||
|---|---|---|---|---|---|---|---|---|---|
| HI | MN | HI | MN | HI | MN | HI | MN | ||
| 20 | 1 | ≥160 | |||||||
| 56 | 2 | 40 | |||||||
| 60 | 2 | 40 | |||||||
| 82 | 2 | 40 | |||||||
| 100 | 2 | 40 | 40 | ||||||
| 105 | 2 | 80 | |||||||
| 128 | 2 | 40 | 40 | ||||||
| 134 | 2 | 40 | |||||||
| 200 | 3 | ≥160 | ≥160 | ≥160 | 160 | 160 | 80 | ||
| 203 | 3 | 80 | 80 | 160 | 160 | 80 | ≥160 | 80 | |
| 209 | 3 | ≥160 | 80 | 160 | ≥160 | 160 | ≥160 | 160 | |
| 212 | 3 | ≥160 | 160 | 80 | 80 | ≥160 | 160 | ||
| 213 | 3 | ≥160 | 160 | 160 | 80 | ≥160 | 80 | ||
| 214 | 3 | ≥160 | 160 | 160 | 80 | ≥160 | 80 | ||
| 221 | 3 | 80 | ≥160 | 80 | 80 | ≥160 | 160 | ||
| 222 | 3 | 40 | 160 | 40 | 80 | 80 | 80 | ||
| 231 | 3 | 40 | |||||||
| 232 | 3 | 40 | |||||||
| 235 | 3 | 80 | 160 | 160 | 320 | ≥160 | 80 | ||
| 237 | 3 | 80 | ≥160 | 160 | 320 | ≥160 | 160 | ||
| 239 | 3 | 80 | 160 | 40 | 80 | 160 | 160 | ||
| 240 | 3 | ≥160 | ≥160 | 160 | 80 | ≥160 | 320 | ||
| 241 | 3 | ≥160 | 160 | 160 | 80 | ≥160 | 320 | ||
| 250 | 3 | 80 | ≥160 | 80 | 80 | 160 | 160 | ||
| 251 | 3 | 40 | 160 | ||||||
| 255 | 3 | ≥160 | ≥160 | ≥160 | 80 | ≥160 | 320 | ||
| 257 | 3 | ≥160 | ≥160 | 80 | 80 | 80 | 160 | ||
| 258 | 3 | ≥160 | ≥160 | 160 | 80 | ≥160 | 160 | ||
| 259 | 3 | ≥160 | ≥160 | 160 | 160 | ≥160 | 160 | ||
| 268 | 3 | ≥160 | ≥160 | 80 | 160 | ≥160 | 80 | ||
| 270 | 3 | 80 | ≥160 | 40 | 80 | 80 | |||
| 271 | 3 | ≥160 | ≥160 | 80 | 160 | ≥160 | 80 | ||
| 272 | 3 | ≥160 | ≥160 | 80 | 160 | ≥160 | |||
| 273 | 3 | 80 | ≥160 | 80 | 80 | ≥160 | 320 | ||
| 274 | 3 | 80 | ≥160 | 80 | 80 | ≥160 | 80 | ||
| 275 | 3 | ≥160 | ≥160 | ≥160 | 160 | ≥160 | |||
| 277 | 3 | 40 | 40 | ||||||
| 278 | 3 | ≥160 | 160 | 80 | 160 | ≥160 | 80 | ||
| 281 | 3 | ≥160 | ≥160 | 80 | 160 | 160 | |||
| 283 | 3 | ≥160 | 80 | 80 | 80 | ≥160 | |||
| 284 | 3 | ≥160 | ≥160 | ≥160 | 160 | ≥160 | |||
| 285 | 3 | ≥160 | ≥160 | 320 | ≥160 | ||||
| 286 | 3 | ≥160 | ≥160 | 80 | 160 | ≥160 | |||
| 287 | 3 | ≥160 | 80 | 80 | 80 | ≥160 | |||
| 288 | 3 | 80 | 40 | ≥160 | |||||
| 289 | 3 | ≥160 | ≥160 | 160 | 160 | ≥160 | |||
| 290 | 3 | 80 | 80 | 80 | 80 | 80 | |||
| 291 | 3 | 80 | 160 | 160 | 80 | ≥160 | |||
| 292 | 3 | 80 | ≥160 | 160 | 80 | ≥160 | |||
| 293 | 3 | 80 | 160 | 80 | 80 | ≥160 | |||
| 294 | 3 | 80 | ≥160 | 80 | 80 | ≥160 | |||
| 295 | 3 | 80 | 160 | 80 | 80 | ≥160 | |||
| 296 | 3 | 80 | 160 | 40 | 40 | ||||
| 297 | 3 | 40 | 40 | 40 | |||||
| 298 | 3 | 40 | 80 | ||||||
| 299 | 3 | 80 | 80 | 80 | 160 | 80 | |||
| 300 | 3 | 80 | 80 | 80 | 160 | 80 | |||
| 303 | 3 | 40 | 80 | 40 | 80 | ||||
| 304 | 3 | 40 | 80 | 160 | |||||
| 307 | 3 | 40 | 80 | 40 | 160 | ||||
| 308 | 3 | 80 | 80 | 80 | 80 | 80 | 160 | ||
| 309 | 3 | 40 | ≥160 | 40 | 80 | 40 | |||
| 310 | 3 | 40 | 80 | 40 | 40 | ||||
| 334 | 3 | 80 | 80 | 80 | 80 | 160 | 40 | ||
| 335 | 3 | 80 | 80 | 80 | 160 | 80 | 80 | ||
| 338 | 3 | 80 | 80 | 80 | 160 | 40 | 80 | ||
| 339 | 3 | 80 | 80 | 80 | 160 | 80 | |||
| Total number of positive sera | 59 | 2 | 65 | 1 | 51 | 47 | 54 | 27 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ilyicheva, T.; Marchenko, V.; Pyankova, O.; Moiseeva, A.; Nhai, T.T.; Lan Anh, B.T.; Sau, T.K.; Kuznetsov, A.; Ryzhikov, A.; Maksyutov, R. Antibodies to Highly Pathogenic A/H5Nx (Clade 2.3.4.4) Influenza Viruses in the Sera of Vietnamese Residents. Pathogens 2021, 10, 394. https://doi.org/10.3390/pathogens10040394
Ilyicheva T, Marchenko V, Pyankova O, Moiseeva A, Nhai TT, Lan Anh BT, Sau TK, Kuznetsov A, Ryzhikov A, Maksyutov R. Antibodies to Highly Pathogenic A/H5Nx (Clade 2.3.4.4) Influenza Viruses in the Sera of Vietnamese Residents. Pathogens. 2021; 10(4):394. https://doi.org/10.3390/pathogens10040394
Chicago/Turabian StyleIlyicheva, Tatyana, Vasily Marchenko, Olga Pyankova, Anastasia Moiseeva, Tran Thi Nhai, Bui Thi Lan Anh, Trinh Khac Sau, Andrey Kuznetsov, Alexander Ryzhikov, and Rinat Maksyutov. 2021. "Antibodies to Highly Pathogenic A/H5Nx (Clade 2.3.4.4) Influenza Viruses in the Sera of Vietnamese Residents" Pathogens 10, no. 4: 394. https://doi.org/10.3390/pathogens10040394
APA StyleIlyicheva, T., Marchenko, V., Pyankova, O., Moiseeva, A., Nhai, T. T., Lan Anh, B. T., Sau, T. K., Kuznetsov, A., Ryzhikov, A., & Maksyutov, R. (2021). Antibodies to Highly Pathogenic A/H5Nx (Clade 2.3.4.4) Influenza Viruses in the Sera of Vietnamese Residents. Pathogens, 10(4), 394. https://doi.org/10.3390/pathogens10040394