
Invasiveness of Escherichia coli Is Associated with an IncFII Plasmid

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
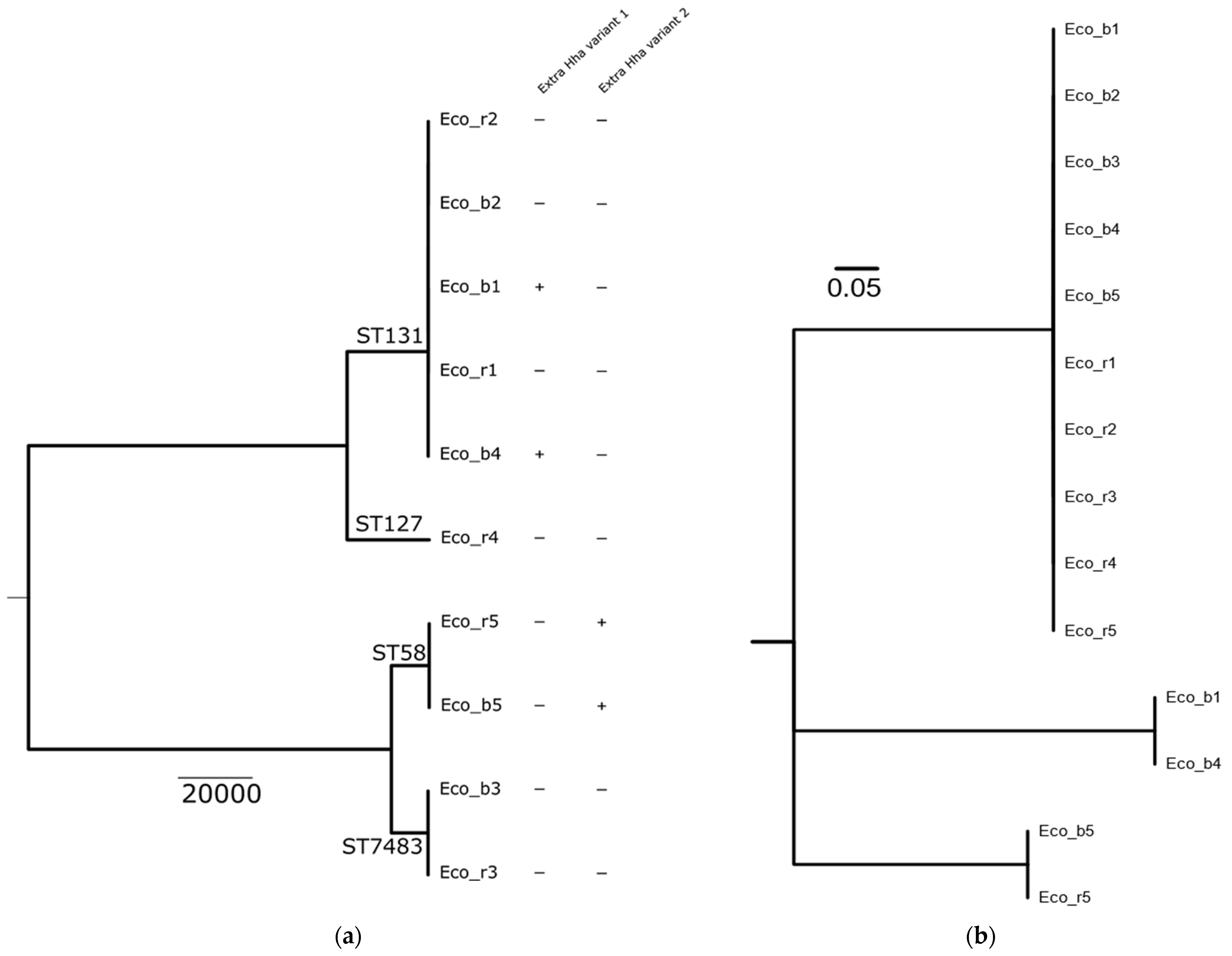
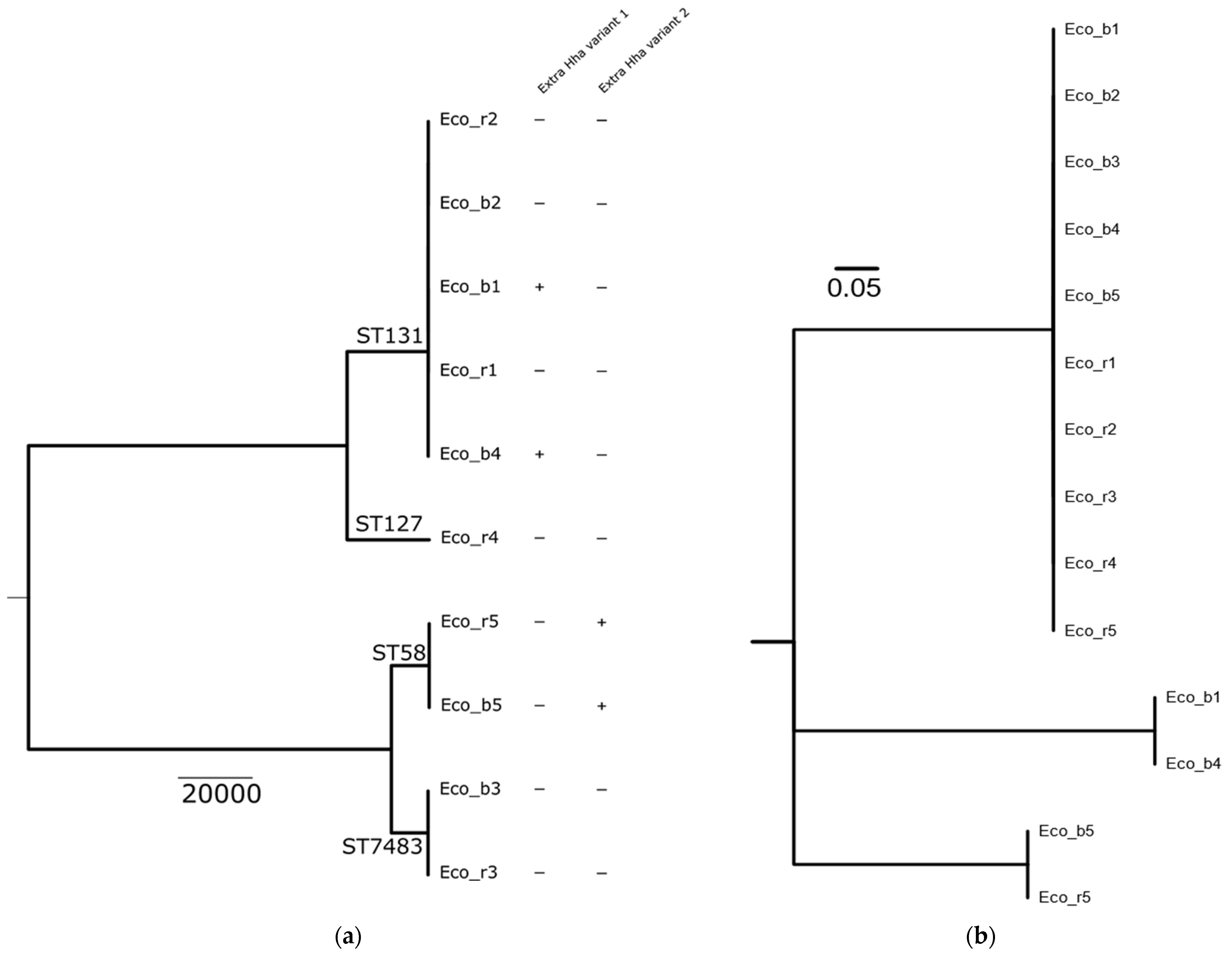
2.1. Clinical Isolates and Molecular Characterization
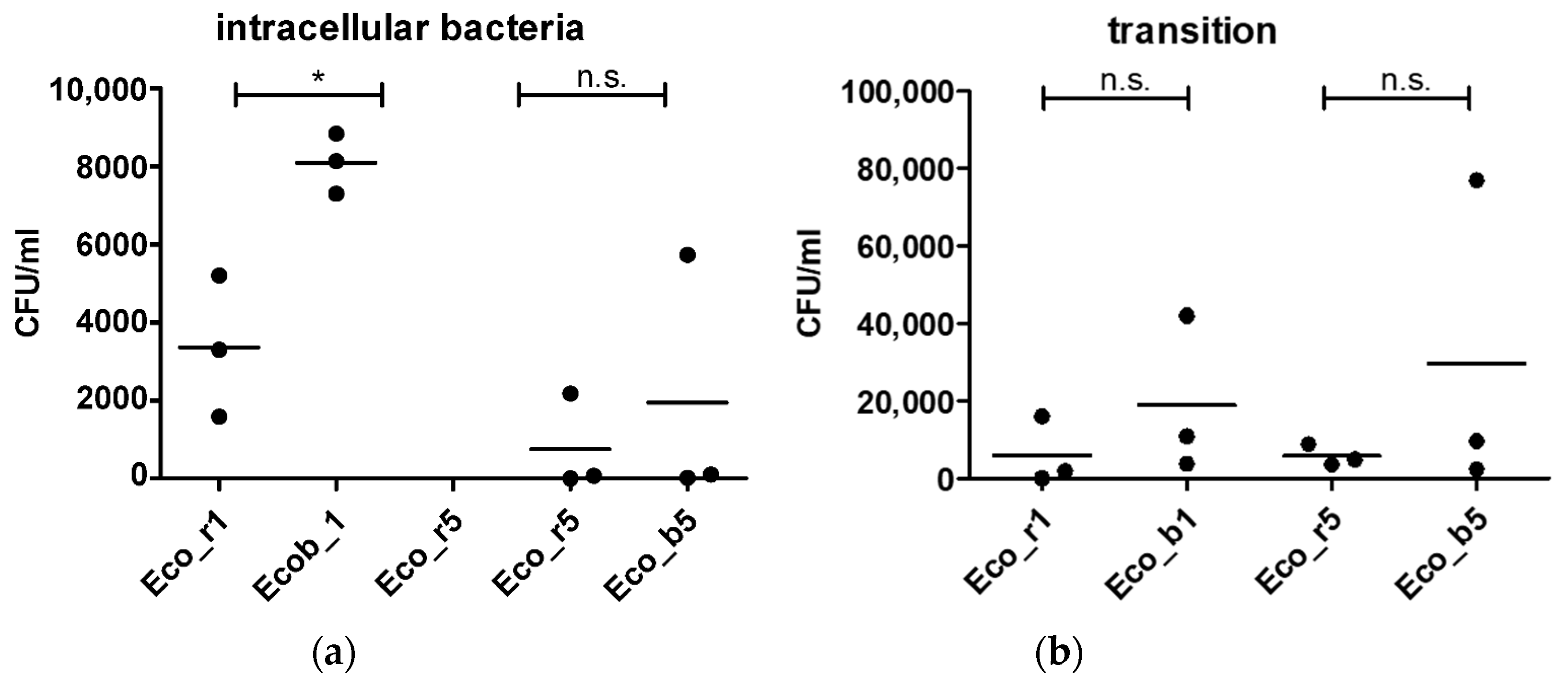
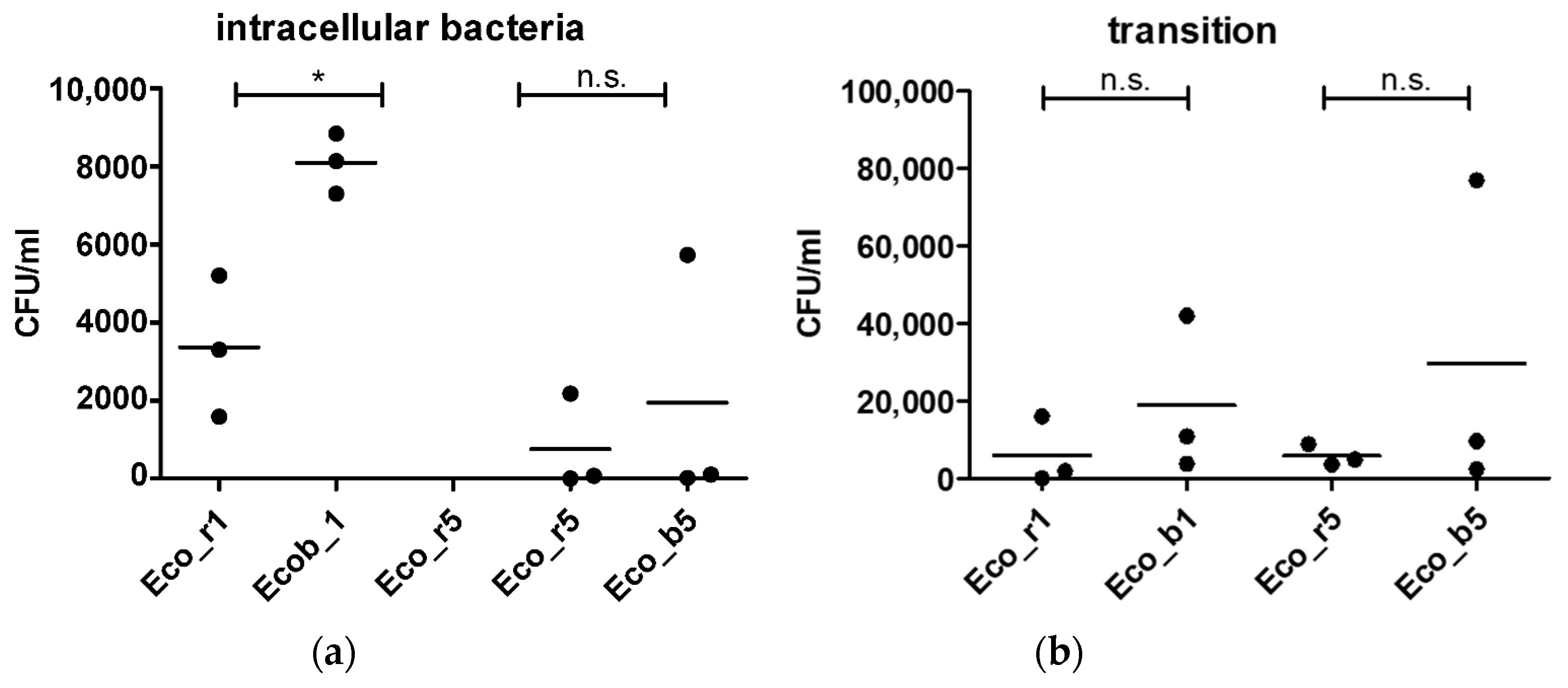
2.2. In Vitro Invasiveness of Clinical E. coli Isolates
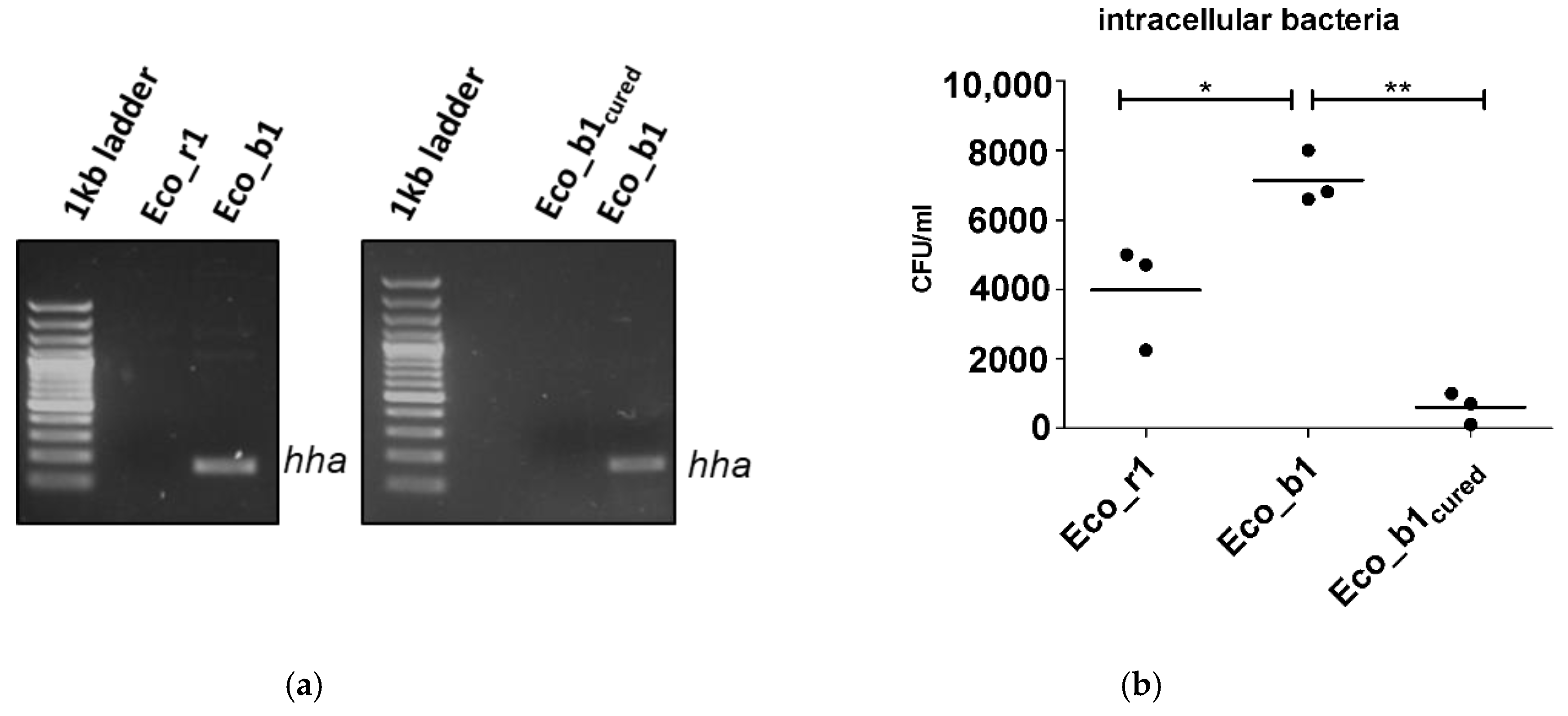
2.3. Loss of Function by Plasmid Curing
2.4. Confirmation of Findings in a Gut Organoid Model
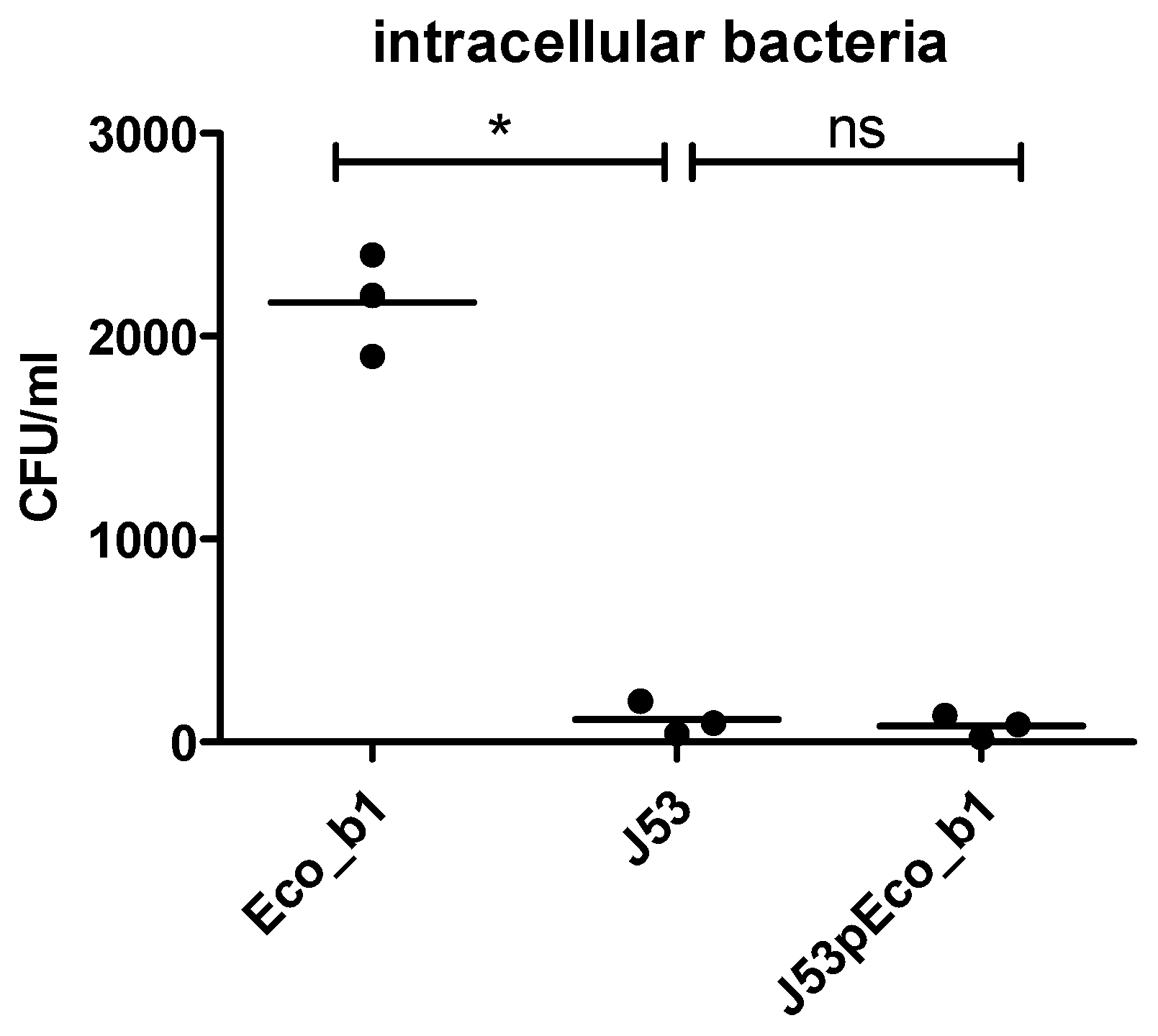
2.5. Plasmid Transfer into J53 E. coli
3. Discussion
4. Materials and Methods
4.1. Study Premise and Clinical E. coli Isolates
4.2. Whole Genome Sequencing and Data Analyses
4.3. Data Availability, Accession and Sequence Statistics
4.4. Invasion and Permeation Assay
4.5. Plasmid Curing and PCR for hha
4.6. Gut Organoid Model
4.7. Plasmid Transfer
4.8. Statistical Analysis and Data Presentation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Manges, A.R.; Geum, H.M.; Guo, A.; Edens, T.J.; Fibke, C.D.; Pitout, J.D.D. Global Extraintestinal Pathogenic Escherichia coli (ExPEC) Lineages. Clin. Microbiol. Rev. 2019, 32, e00135-18. [Google Scholar] [CrossRef] [PubMed]
- Berg, R.D. Bacterial translocation from the gastrointestinal tract. Trends Microbiol. 1995, 3, 149–154. [Google Scholar] [CrossRef]
- Rozwandowicz, M.; Brouwer, M.S.M.; Fischer, J.; Wagenaar, J.A.; Gonzalez-Zorn, B.; Guerra, B.; Mevius, D.J.; Hordijk, J. Plasmids carrying antimicrobial resistance genes in Enterobacteriaceae. J Antimicrob. Chemother. 2018, 73, 1121–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieto, J.M.; Carmona, M.; Bolland, S.; Jubete, Y.; de la Cruz, F.; Juarez, A. The hha gene modulates haemolysin expression in Escherichia coli. Mol. Microbiol. 1991, 5, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Navarre, W.W.; McClelland, M.; Libby, S.J.; Fang, F.C. Silencing of xenogeneic DNA by H-NS-facilitation of lateral gene transfer in bacteria by a defense system that recognizes foreign DNA. Genes Dev. 2007, 21, 1456–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, V.K.; Bearson, B.L. Hha controls Escherichia coli O157:H7 biofilm formation by differential regulation of global transcriptional regulators FlhDC and CsgD. Appl. Environ. Microbiol. 2013, 79, 2384–2396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, V.K.; Carlson, S.A.; Casey, T.A. Hyperadherence of an hha mutant of Escherichia coli O157:H7 is correlated with enhanced expression of LEE-encoded adherence genes. FEMS Microbiol. Lett. 2005, 243, 189–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazen, T.H.; Michalski, J.; Nagaraj, S.; Okeke, I.N.; Rasko, D.A. Characterization of a Large Antibiotic Resistance Plasmid Found in Enteropathogenic Escherichia coli Strain B171 and Its Relatedness to Plasmids of Diverse E. coli and Shigella Strains. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madrid, C.; Garcia, J.; Pons, M.; Juarez, A. Molecular evolution of the H-NS protein: Interaction with Hha-like proteins is restricted to enterobacteriaceae. J. Bacteriol. 2007, 189, 265–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madrid, C.; Balsalobre, C.; Garcia, J.; Juarez, A. The novel Hha/YmoA family of nucleoid-associated proteins: Use of structural mimicry to modulate the activity of the H-NS family of proteins. Mol. Microbiol. 2007, 63, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Garcillan-Barcia, M.P.; Francia, M.V.; de la Cruz, F. The diversity of conjugative relaxases and its application in plasmid classification. FEMS Microbiol. Rev. 2009, 33, 657–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solorzano, C.; Srikumar, S.; Canals, R.; Juarez, A.; Paytubi, S.; Madrid, C. Hha has a defined regulatory role that is not dependent upon H-NS or StpA. Front. Microbiol. 2015, 6, 773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrios, A.F.; Zuo, R.; Ren, D.; Wood, T.K. Hha, YbaJ, and OmpA regulate Escherichia coli K12 biofilm formation and conjugation plasmids abolish motility. Biotechnol. Bioeng. 2006, 93, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Bedzyk, L.A.; Thomas, S.M.; Ye, R.W.; Wood, T.K. Gene expression in Escherichia coli biofilms. Appl. Microbiol. Biotechnol. 2004, 64, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Prieto, A.; Urcola, I.; Blanco, J.; Dahbi, G.; Muniesa, M.; Quiros, P.; Falgenhauer, L.; Chakraborty, T.; Huttener, M.; Juarez, A. Tracking bacterial virulence: Global modulators as indicators. Sci. Rep. 2016, 6, 25973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahlen, T.F.; Wilson, R.L.; Boddicker, J.D.; Jones, B.D. Hha is a negative modulator of transcription of hilA, the Salmonella enterica serovar Typhimurium invasion gene transcriptional activator. J. Bacteriol. 2001, 183, 6620–6629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silphaduang, U.; Mascarenhas, M.; Karmali, M.; Coombes, B.K. Repression of intracellular virulence factors in Salmonella by the Hha and YdgT nucleoid-associated proteins. J. Bacteriol. 2007, 189, 3669–3673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocer, K.; Boutin, S.; Dalpke, A.H.; Heeg, K.; Mutters, N.T.; Nurjadi, D. Comparative genomic analysis reveals a high prevalence of inter-species in vivo transfer of carbapenem-resistance plasmids in patients with haematological malignancies. Clin. Microbiol. Infect 2020, 26, 780.e1–780.e8. [Google Scholar] [CrossRef] [PubMed]
- Isberg, R.; Falkow, S. A single genetic locus encoded by Yersinia pseudotuberculosis permits invasion of cultured animal cells by Escherichia coli K-12. Nature 1985, 317, 262–264. [Google Scholar] [CrossRef] [PubMed]
- Stanifer, M.L.; Mukenhirn, M.; Muenchau, S.; Pervolaraki, K.; Kanaya, T.; Albrecht, D.; Odendall, C.; Hielscher, T.; Haucke, V.; Kagan, J.C.; et al. Asymmetric distribution of TLR3 leads to a polarized immune response in human intestinal epithelial cells. Nat. Microbiol. 2020, 5, 181–191. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Accession | MLST | Coverage | #Contigs | Largest Contig | Total Length | GC(%) | N50 | N75 | L50 | L75 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Eco_b1 | SAMN20169821 | 131 | 70 | 81 | 596,711 | 5,127,344 | 50.76 | 218,234 | 124,147 | 8 | 15 |
| Eco_b1_cured | SAMN20169822 | 131 | 39 | 77 | 596,711 | 5,133,246 | 50.76 | 222,766 | 124,147 | 8 | 15 |
| Eco_b2 | SAMN20169823 | 131 | 54 | 59 | 623,297 | 5,055,866 | 50.75 | 240,470 | 124,868 | 7 | 14 |
| Eco_b3 | SAMN20169824 | 7483 | 33 | 68 | 413,140 | 4,918,104 | 50.81 | 192,174 | 102,049 | 10 | 18 |
| Eco_b4 | SAMN20169825 | 131 | 62 | 73 | 623,463 | 5,139,954 | 50.74 | 222,558 | 135,897 | 7 | 14 |
| Eco_b5 | SAMN20169826 | 58 | 28 | 91 | 345,561 | 4,863,584 | 50.71 | 132,372 | 61,122 | 13 | 25 |
| Eco_r1 | SAMN20169827 | 131 | 59 | 72 | 596,712 | 5,195,130 | 50.71 | 222,897 | 124,147 | 7 | 15 |
| Eco_r2 | SAMN20169828 | 131 | 60 | 61 | 623,297 | 5,098,506 | 50.75 | 240,470 | 124,868 | 7 | 14 |
| Eco_r3 | SAMN20169829 | 7483 | 38 | 63 | 381,423 | 4,918,014 | 50.81 | 213,742 | 119,893 | 9 | 17 |
| Eco_r4 | SAMN20169830 | 127 | 57 | 42 | 1,600,676 | 5,006,799 | 50.42 | 410,665 | 217,547 | 3 | 7 |
| Eco_r5 | SAMN20169831 | 58 | 31 | 94 | 326,419 | 4,904,693 | 50.68 | 132,372 | 61,122 | 13 | 26 |
| J53_Eco_b1 | SAMN20169832 | 10 | 91 | 90 | 414,105 | 4,766,686 | 50.77 | 125,945 | 64,036 | 13 | 26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krall, L.J.; Klein, S.; Boutin, S.; Wu, C.C.; Sähr, A.; Stanifer, M.L.; Boulant, S.; Heeg, K.; Nurjadi, D.; Hildebrand, D. Invasiveness of Escherichia coli Is Associated with an IncFII Plasmid. Pathogens 2021, 10, 1645. https://doi.org/10.3390/pathogens10121645
Krall LJ, Klein S, Boutin S, Wu CC, Sähr A, Stanifer ML, Boulant S, Heeg K, Nurjadi D, Hildebrand D. Invasiveness of Escherichia coli Is Associated with an IncFII Plasmid. Pathogens. 2021; 10(12):1645. https://doi.org/10.3390/pathogens10121645
Chicago/Turabian StyleKrall, Lars Johannes, Sabrina Klein, Sébastien Boutin, Chia Ching Wu, Aline Sähr, Megan L. Stanifer, Steeve Boulant, Klaus Heeg, Dennis Nurjadi, and Dagmar Hildebrand. 2021. "Invasiveness of Escherichia coli Is Associated with an IncFII Plasmid" Pathogens 10, no. 12: 1645. https://doi.org/10.3390/pathogens10121645
APA StyleKrall, L. J., Klein, S., Boutin, S., Wu, C. C., Sähr, A., Stanifer, M. L., Boulant, S., Heeg, K., Nurjadi, D., & Hildebrand, D. (2021). Invasiveness of Escherichia coli Is Associated with an IncFII Plasmid. Pathogens, 10(12), 1645. https://doi.org/10.3390/pathogens10121645