
The Absence of Abdominal Pigmentation in Livestock Associated Culicoides following Artificial Blood Feeding and the Epidemiological Implication for Arbovirus Surveillance

 , , and
, , and
Abstract
:1. Introduction
2. Results
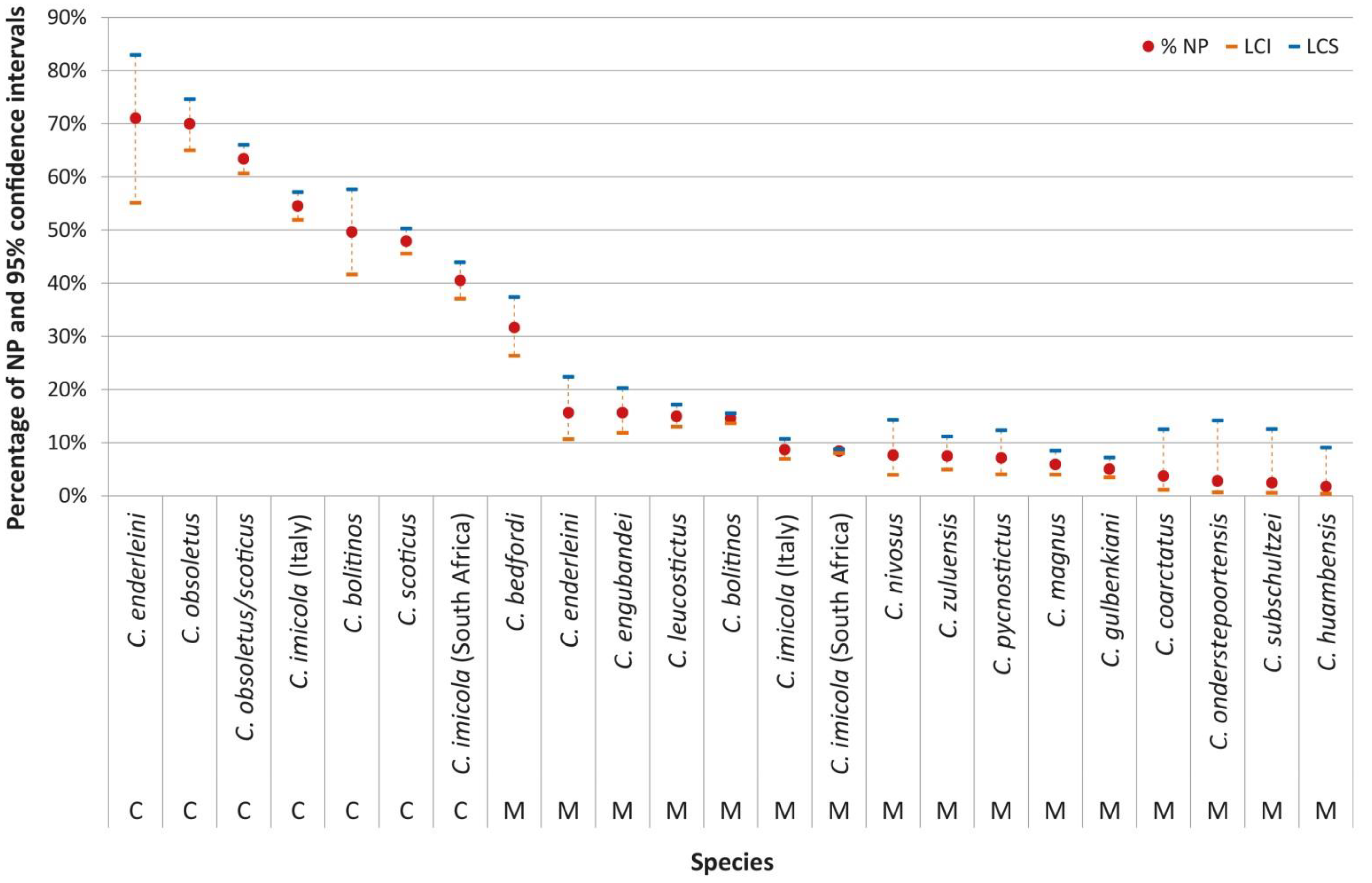
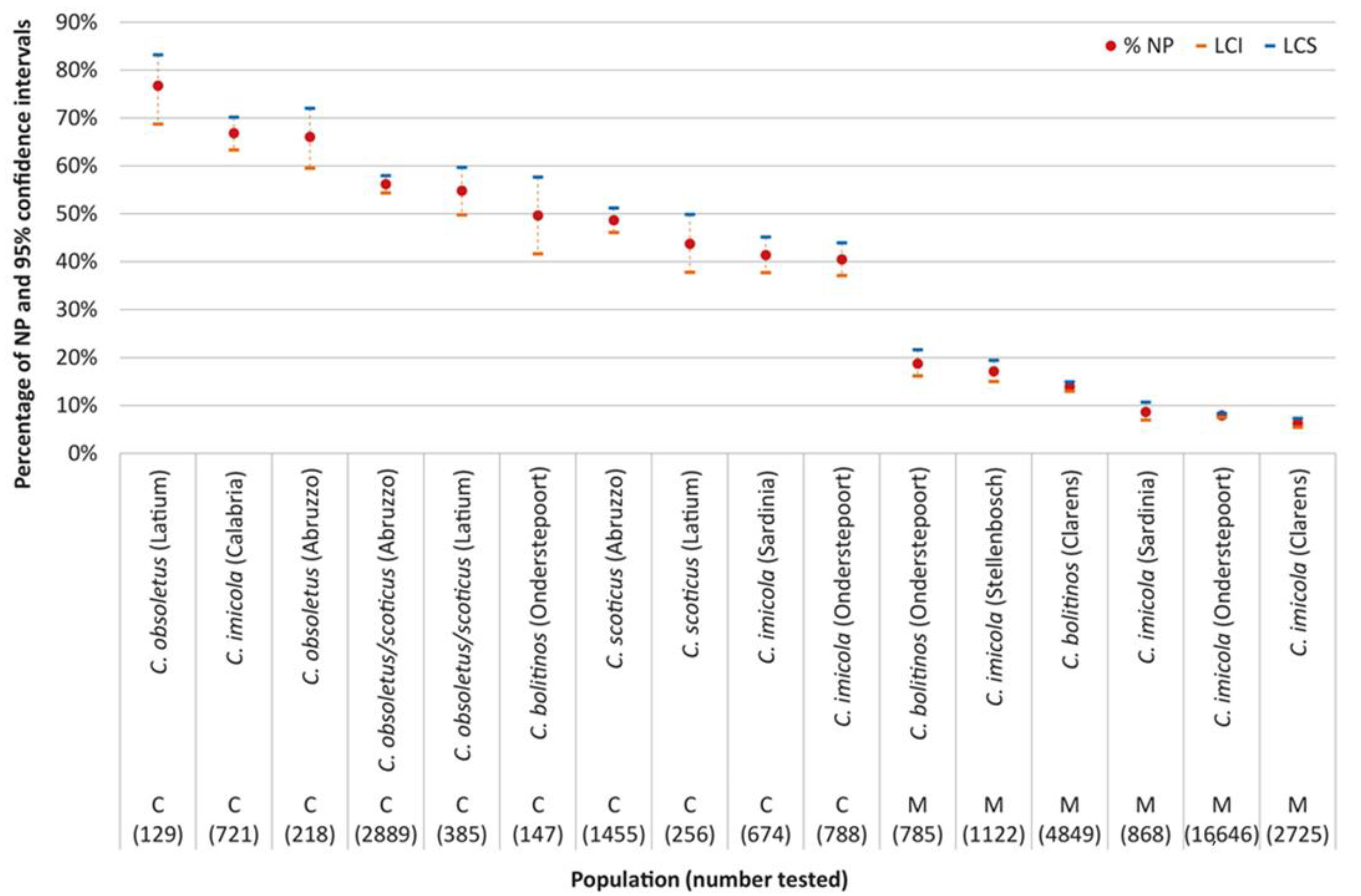
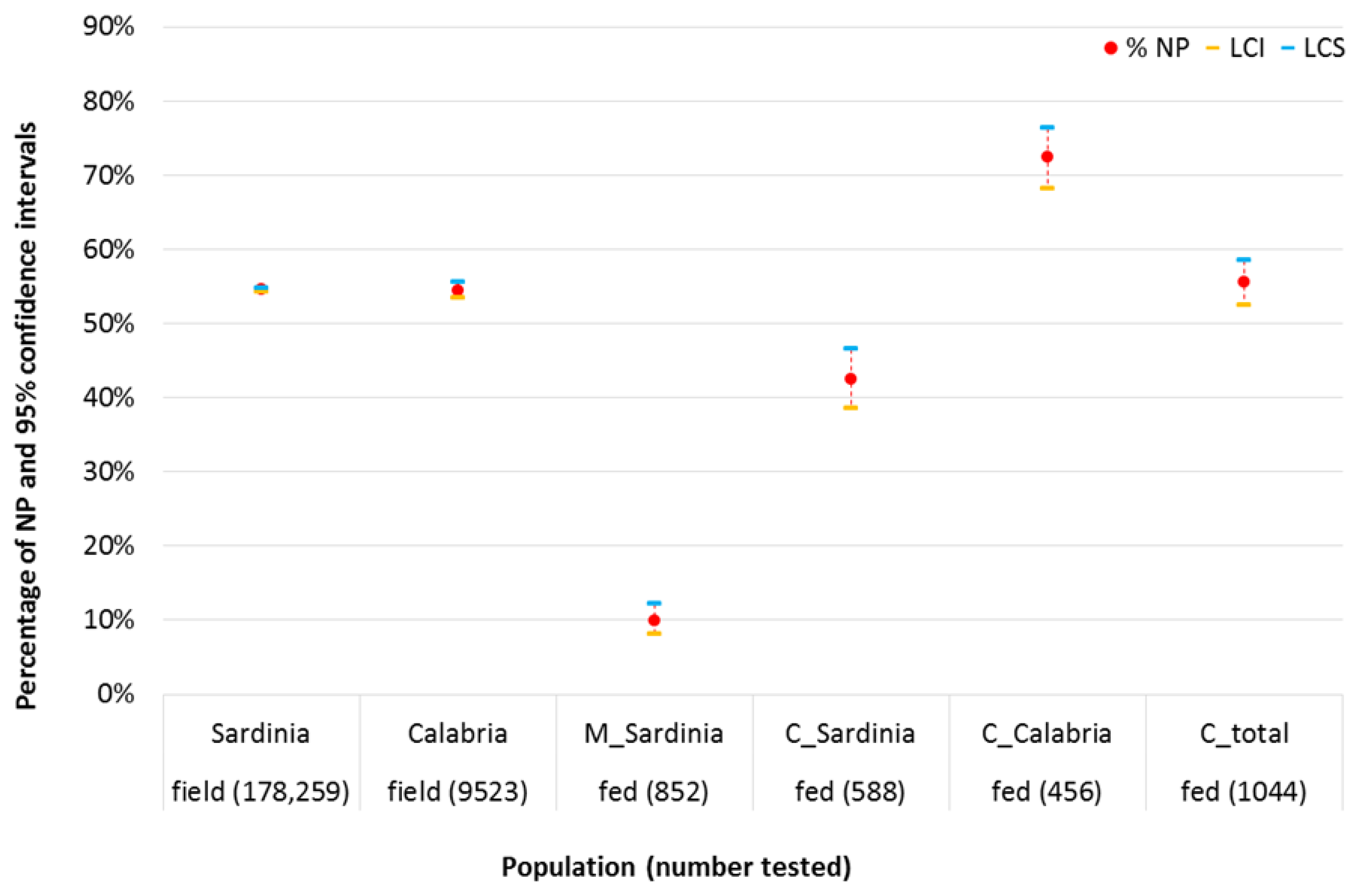
2.1. Comparison between Laboratory Feeding Results and Field Data
2.2. Repeated Feeding
2.3. Virus Detection
3. Discussion
4. Materials and Methods
4.1. Specimen Collection and Artificial Feeding
4.2. Comparison between Laboratory Feeding Results and Field Data
4.3. Repeated Feeding
4.4. Virus Detection
4.5. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Wilson, A.; Darpel, K.; Mellor, P.S. Where does bluetongue virus sleep in the winter? PLoS Biol. 2008, 6, e210. [Google Scholar] [CrossRef] [Green Version]
- Conte, A.; Goffredo, M.; Candeloro, L.; Calistri, P.; Curci, G.; Colaiuda, V.; Quaglia, M.; Mancini, G.; Santilli, A.; Di Lorenzo, A.; et al. Analysis of climatic factors involved in the BTV-1 incursion in Central Italy in 2014. Vet. Ital. 2016, 52, 223–229. [Google Scholar]
- Rodríguez-Sánchez, B.; Iglesias-Martín, I.; Martínez-Avilés, M.; Sánchez-Vizcaíno, J.M. Orbiviruses in the Mediterranean basin: Updated epidemiological situation of bluetongue and new methods for the detection of BTV serotype 4. Transbound. Emerg. Dis. 2008, 55, 205–214. [Google Scholar] [CrossRef]
- Saegerman, C.; Berkvens, D.; Mellor, P.S. Bluetongue epidemiology in the European Union. Emerg. Infect. Dis. 2008, 14, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Conraths, F.J.; Gethmann, J.M.; Staubach, C.; Mettenleiter, T.C.; Beer, M.; Hoffmann, B. Epidemiology of bluetongue virus serotype 8, Germany. Emerg. Infect. Dis. 2009, 15, 433–435. [Google Scholar] [CrossRef]
- Conte, A.; Gilbert, M.; Goffredo, M. Eight years of entomological surveillance in Italy show no evidence of Culicoides imicola geographical range expansion. J. Appl. Ecol. 2009, 46, 1332–1339. [Google Scholar]
- Falconi, C.; López-Olvera, J.R.; Gortázar, C. BTV infection in wild ruminants, with emphasis on red deer: A review. Vet. Microbiol. 2011, 151, 209–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, B.; Scheuch, M.; Höper, D.; Jungblut, R.; Holsteg, M.; Schirrmeier, H.; Eschbaumer, M.; Goller, K.V.; Wernike, K.; Fischer, M.; et al. Novel Orthobunyavirus in cattle, Europe, 2011. Emerg. Infect. Dis. 2012, 18, 469–472. [Google Scholar] [CrossRef] [PubMed]
- Goffredo, M.; Monaco, F.; Capelli, G.; Quaglia, M.; Federici, V.; Catalani, M.; Montarsi, F.; Polci, A.; Pinoni, C.; Calistri, P.; et al. Schmallenberg virus in Italy: A retrospective survey in Culicoides stored during the bluetongue Italian surveillance program. Prev. Vet. Med. 2013, 111, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Balenghien, T.; Pagès, N.; Goffredo, M.; Carpenter, S.; Augot, D.; Jacquier, E.; Talavera, S.; Monaco, F.; Depaquit, J.; Grillet, C.; et al. The emergence of Schmallenberg virus across Culicoides communities and ecosystems in Europe. Prev. Vet. Med. 2014, 116, 360–369. [Google Scholar] [CrossRef]
- Savini, G.; Afonso, A.; Mellor, P.; Aradaib, I.A.O.; Yadin, H.; Sanaa, M.; Wilson, W.; Monaco, F.; Domingo, M. Epizootic haemorragic disease. Res. Vet. Sci. 2011, 91, 1–17. [Google Scholar] [CrossRef]
- Sick, F.; Beer, M.; Kampen, H.; Wernike, K. Culicoides biting midges—Underestimated vectors for arboviruses of public health and veterinary importance. Viruses 2019, 11, 376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, S.; Rajko-Nenow, P.; Ashby, M.; Frost, L.; Carpenter, S.; Batten, C. Outbreak of African horse sickness in Thailand, 2020. Transbound. Emerg. Dis. 2020, 67, 1764–1767. [Google Scholar] [CrossRef]
- Mellor, P.S. The replication of bluetongue virus in Culicoides vectors. Curr. Top. Microbiol. Immunol. 1990, 162, 143–161. [Google Scholar]
- Allingham, P.; Standfast, H. An investigation of transovarial transmission of Akabane virus in Culicoides brevitarsis. Aust. Vet. J. 1990, 67, 273–274. [Google Scholar] [CrossRef]
- Nunamaker, R.A.; Sieburth, P.J.; Dean, V.C.; Wigington, J.G.; Nunamaker, C.E.; Mecham, J.O. Absence of transovarial transmission of bluetongue virus in Culicoides variipennis: Immunogold labeling of bluetongue virus antigen in developing oocytes from Culicoides variipennis (Coquillett). Comp. Biochem. Physiol. A Comp. Physiol. 1990, 96, 19–31. [Google Scholar] [CrossRef]
- Osborne, C.J.; Mayo, C.E.; Mullens, B.A.; McDermott, E.G.; Gerry, A.C.; Reisen, W.K.; MacLachlan, N.J. Lack of evidence for laboratory and natural vertical transmission of bluetongue virus in Culicoides sonorensis (Diptera: Ceratopogonidae). J. Med. Entomol. 2015, 52, 274–277. [Google Scholar] [CrossRef] [Green Version]
- Venter, G.J.; Koekemoer, J.J.O.; Paweska, J.T. Investigations on outbreaks of African horse sickness in the surveillance zone of South Africa. Rev. Sci. Tech. 2006, 25, 1097–1109. [Google Scholar] [CrossRef] [Green Version]
- Goffredo, M.; Catalani, M.; Federici, V.; Portanti, O.; Marini, V.; Mancini, G.; Quaglia, M.; Santilli, A.; Teodori, L.; Savini, G. Vector species of Culicoides midges implicated in the 2012–2014 Bluetongue epidemics in Italy. Vet. Ital. 2015, 51, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Meiswinkel, R.; Venter, G.J.; Nevill, E.M. Vectors: Culicoides spp. In Infectious Diseases of Livestock, 2nd ed.; Coetzer, J.A.W., Tustin, R.C., Eds.; Oxford University Press: Cape Town, South Africa, 2004; Volume 1, pp. 93–136. [Google Scholar]
- Venter, G.J.; Nevill, E.M.; Van der Linde, T.C. Seasonal abundance and parity of stock-associated Culicoides species (Diptera: Ceratopogonidae) in different climatic regions in southern Africa in relation to their vector potential. Onderstepoort. J. Vet. Res. 1997, 64, 259–271. [Google Scholar]
- Mayo, C.E.; Osborne, C.J.; Mullens, B.A.; Gerry, A.C.; Gardner, I.A.; Reisen, W.K.; Barker, C.M.; MacLachlan, N.J. Seasonal variation and impact of waste-water lagoons as larval habitat on the population dynamics of Culicoides sonorensis (Diptera: Ceratopogonidae) at two dairy farms in Northern California. PLoS ONE 2014, 9, e89633. [Google Scholar] [CrossRef]
- Eurlex. Available online: https://eur-lex.europa.eu/legal-content/en/TXT/?uri=CELEX%3A32020R0689 (accessed on 30 November 2021).
- Lysyk, T.J. Seasonal abundance, parity, and survival of adult Culicoides sonorensis (Diptera: Ceratopogonidae) in Southern Alberta, Canada. J. Med. Entomol. 2007, 44, 959–969. [Google Scholar] [CrossRef]
- Rádrová, J.; Mračková, M.; Galková, Z.; Lamka, J.; Račka, K.; Barták, P.; Votýpka, J. Seasonal dynamics, parity rate, and composition of Culicoides (Diptera: Ceratopogonidae) occurring in the vicinity of wild and domestic ruminants in the Czech Republic. J. Med. Entomol. 2015, 53, 416–424. [Google Scholar] [CrossRef]
- Foxi, C.; Delrio, G.; Falchi, G.; Marche, M.G.; Satta, G.; Ruiu, L. Role of different Culicoides vectors (Diptera: Ceratopogonidae) in bluetongue virus transmission and overwintering in Sardinia (Italy). Parasites Vectors 2016, 9, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyce, A.L. The recognition of nulliparous and parous Culicoides (Diptera: Ceratopogonidae) without dissection. J. Aust. Entomol. Soc. 1969, 8, 11–15. [Google Scholar] [CrossRef]
- Akey, D.H.; Potter, H.W. Pigmentation Associated with Oogenesis in the Biting Fly Culicoides variipennis (Diptera: Ceratopogonidae): Determination of Parity. J. Med. Entomol. 1979, 16, 67–70. [Google Scholar] [CrossRef]
- Linley, J.R.; Braverman, Y. The lateral abdominal pigmentation in Culicoides variipennis and Culicoides furens (Diptera: Ceratopogonidae): Quantitative measurement of its relationship to age and oogenesis. J. Med. Entomol. 1986, 23, 51–63. [Google Scholar] [CrossRef]
- Venter, G.J.; Hill, E.; Pajor, I.T.P.; Nevill, E.M. The use of a membrane feeding technique to determine the infection rate of Culicoides imicola (Diptera, Ceratopogonidae) for 2 bluetongue virus serotypes in South Africa. Onderstepoort. J. Vet. Res. 1991, 58, 5–9. [Google Scholar] [PubMed]
- Goffredo, M.; Romeo, G.; Monaco, F.; Di Gennaro, A.; Savini, G. Laboratory survival and blood feeding response of wild-caught Culicoides obsoletus Complex (Diptera: Ceratopogonidae) through natural and artificial membranes. Vet. Ital. 2004, 40, 282–285. [Google Scholar]
- Venter, G.J.; Paweska, J.T.; Lunt, H.; Mellor, P.S.; Carpenter, S. An alternative method of blood-feeding Culicoides imicola and other haematophagous Culicoides species for vector competence studies. Vet. Parasitol. 2005, 131, 331–335. [Google Scholar] [CrossRef]
- Carpenter, S.; Lunt, H.L.; Arav, D.; Venter, G.J.; Mellor, P.S. Oral susceptibility to bluetongue virus of Culicoides (Diptera: Ceratopogonidae) from the United Kingdom. J. Med. Entomol. 2006, 43, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Veronesi, E.; Henstock, M.; Gubbins, S.; Batten, C.; Manley, R.; Barber, J.; Carpenter, S. Implicating Culicoides biting midges as vectors of Schmallenberg virus using semi-quantitative RT-PCR. PLoS ONE 2013, 8, e57747. [Google Scholar] [CrossRef] [Green Version]
- Federici, V.; Ippoliti, C.; Goffredo, M.; Catalani, M.; Di Provvido, A.; Santilli, A.; Quaglia, M.; Mancini, G.; Di Nicola, F.; Di Gennaro, A.; et al. Epizootic haemorrhagic disease in Italy: Vector competence of indigenous Culicoides species and spatial multicriteria evaluation of vulnerability. Vet. Ital. 2016, 52, 271–279. [Google Scholar] [PubMed]
- Van Der Saag, M.R.; Ward, M.P.; Kirkland, D. Application of an embryonated chicken egg model to assess the vector competence of Australian Culicoides midges for bluetongue viruses: Vector competence model for Culicoides. Med. Vet. Entomol. 2017, 31, 263–271. [Google Scholar] [CrossRef]
- Paslaru, A.I.; Mathis, A.; Torgerson, P.; Veronesi, E. Vector competence of pre-alpine Culicoides (Diptera: Ceratopogonidae) for bluetongue virus serotypes 1, 4 and 8. Parasites Vectors 2018, 11, 466. [Google Scholar] [CrossRef]
- Federici, V.; Goffredo, M.; Mancini, G.; Quaglia, M.; Santilli, A.; Di Nicola, F.; De Ascentis, M.; Cabras, P.; Volpicelli, C.; De Liberato, C.; et al. Vector competence of Italian populations of Culicoides for some bluetongue virus strains responsible for recent northern African and European outbreaks. Viruses 2019, 11, 941. [Google Scholar] [CrossRef] [Green Version]
- Venter, G.J. Culicoides spp. (Diptera: Ceratopogonidae) as vectors of bluetongue virus in South Africa—A review. Vet. Ital. 2016, 51, 325–333. [Google Scholar] [CrossRef]
- Venter, G.J. Culicoides (Diptera: Ceratopogonidae) as vectors of African horse sickness virus in South Africa. In Abstracts of the XXV International Congress of Entomology, Proceedings of XXV International Congress of Entomology, Orlando, FL, USA, 25–30 September2016; Entomological Society of America: Annapolis, MD, USA, 2016. [Google Scholar] [CrossRef]
- Goffredo, M.; Meiswinkel, R.; Federici, V.; Di Nicola, F.; Mancini, G.; Ippoliti, C.; Di Lorenzo, A.; Quaglia, M.; Santilli, A.; Conte, A.; et al. The ‘Culicoides obsoletus group’ in Italy: Relative abundance, geographic range, and role as vector for Bluetongue virus. Vet. Ital. 2016, 52, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Savini, G.; Goffredo, M.; Monaco, F.; Di Gennaro, A.; Cafiero, M.A.; Baldi, L.; De Santis, P.; Meiswinkel, R.; Caporale, V. Bluetongue virus isolations from midges belonging to the Obsoletus complex (Culicoides, Diptera: Ceratopogonidae) in Italy. Vet. Rec. 2005, 157, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Kettle, D.S. The bionomics and control of Culicoides and Leptoconops (Diptera, Ceratopogonidae = Heleidae). Annu. Rev. Entomol. 1962, 7, 401–418. [Google Scholar] [CrossRef]
- Walker, A.R.; Boreham, P.F.L. Blood feeding of Culicoides (Diptera, Ceratopogonidae) in Kenya in relation to the epidemiology of bluetongue and ephemeral fever. Bull. Entomol. Res. 1976, 66, 181–188. [Google Scholar] [CrossRef]
- De Beer, C.J.; Boikanyo, S.N.B.; Venter, G.J. Assessment of the Hemotek® system for the in vitro feeding of field-collected Culicoides imicola (Diptera: Ceratopogonidae) in South Africa. Med. Vet. Entomol. 2021, 35, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Veronesi, E.; Venter, G.J.; Labuschagne, K.; Mellor, P.S.; Carpenter, S. Life-history parameters of Culicoides (Avaritia) imicola Kieffer in the laboratory at different rearing temperatures. Vet. Parasitol. 2009, 163, 370–373. [Google Scholar] [CrossRef] [PubMed]
- Nevill, E.M. Biological Studies on Some South African Culicoides Species (Diptera: Ceratopogonidae) and the Morphology of Their Immature Stages. Master’s Thesis, University of Pretoria, Pretoria, South Africa, 1967. [Google Scholar]
- Braverman, Y.; Linley, J.R. Fecundity and proportions of gravid females in populations of the bluetongue vector Culicoides imicola (Diptera: Ceratopogonidae) and several other species in Israel. J. Med. Entomol. 1994, 31, 838–843. [Google Scholar] [CrossRef]
- Barcelò, C.; Miranda, M.A. Bionomics of livestock-associated Culicoides (biting midge) bluetongue virus vectors under laboratory conditions. Med. Vet. Entomol. 2018, 32, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Birley, M.H.; Boorman, J.P.T. Estimating the survival and biting rates of haematophagous insects, with particular reference to the Culicoides obsoletus group (Diptera, Ceratopogonidae) in southern England. J. Anim. Ecol. 1982, 51, 135–148. [Google Scholar] [CrossRef]
- Van der Saag, M.; Nicholas, A.; Ward, M.; Kirkland, P. Evaluation of in vitro methods for assessment of infection of Australian Culicoides spp. with Bluetongue viruses. Vet. Ital. 2015, 51, 335–343. [Google Scholar] [CrossRef]
- Bonneau, K.R.; DeMaula, C.D.; Mullens, B.A.; MacLachlan, N.J. Duration of viraemia infectious to Culicoides sonorensis in bluetongue virus-infected cattle and sheep. Vet. Microbiol. 2002, 88, 115–125. [Google Scholar] [CrossRef]
- Lehane, M.J. The Biology of Blood-Sucking in Insects; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Friend, W.G.; Smith, J.J.B. Factors affecting feeding by bloodsucking insects. Annu. Rev. Entomol. 1977, 22, 309–331. [Google Scholar] [CrossRef]
- Davis, E.L.; Butler, J.F.; Roberts, R.H.; Reinert, J.F.; Kline, D.L. Laboratory blood feeding of Culicoides mississippiensis (Diptera: Ceratopogonidae) through a reinforced silicone membrane. J. Med. Entomol. 1983, 20, 177–182. [Google Scholar] [CrossRef]
- Larska, M.; Lechowski, L.; Grochowsk, M.; Żmudziński, J.F. Detection of the Schmallenberg virus in nulliparous Culicoides obsoletus/scoticus complex and C. punctatus—The possibility of transovarial virus transmission in the midge population and of a new vector. Vet. Microbiol. 2013, 166, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Kęsik-Maliszewska, J.; Larska, M.; Collins, Á.B.; Rola, J. Post-epidemic distribution of Schmallenberg Virus in Culicoides arbovirus vectors in Poland. Viruses 2019, 11, 447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozo-Lopez, P.; Londono-Renteria, B.; Drolet, B.S. Venereal transmission of vesicular stomatitis virus by Culicoides sonorensis Midges. Pathogens 2020, 9, 316. [Google Scholar] [CrossRef]
- Braverman, Y.; Mumcuoglu, K. Newly emerged nulliparous Culicoides imicola Kieffer (Diptera: Ceratopogonidae) with pigmented abdomen. Vet. Parasitol. 2009, 160, 356–358. [Google Scholar] [CrossRef]
- Harrup, L.E.; Purse, B.V.; Golding, N.; Mellor, P.S.; Carpenter, S. Larval development and emergence sites of farm-associated Culicoides in the United Kingdom. Med. Vet. Entomol. 2013, 27, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Magliano, A.; Scaramozzino, P.; Ravagnan, S.; Montarsi, F.; Da Rold, G.; Cincinelli, G.; Moni, A.; Slvestri, P.; Carvelli, A.; De Liberato, C. Indoor and outdoor winter activity of Culicoides biting midges, vectors of bluetongue virus, in Italy. Med. Vet. Entomol. 2018, 32, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Meiswinkel, R.; Goffredo, M.; Dijkstra, E.G.M.; van der Ven, I.J.K.; Baldet, T.; Elbers, A. Endophily in Culicoides associated with BTV-Infected cattle in the province of Limburg, South-Eastern Netherlands, 2006. Prev. Vet. Med. 2008, 87, 182–195. [Google Scholar] [CrossRef]
- Carpenter, S.; McArthur, C.; Selby, R.; Ward, R.; Nolan, D.V.; Mordue Luntz, A.J.; Dallas, J.F.; Tripet, F.; Mellor, P.S. Experimental infection studies of UK Culicoides species midges with bluetongue virus serotypes 8 and 9. Vet. Rec. 2008, 163, 589–592. [Google Scholar] [CrossRef]
- Venter, G.J.; Wright, I.M.; Del Rio, R.; Lucientes, J.; Miranda, M.A. The susceptibility of Culicoides imicola and other South African livestock-associated Culicoides species to infection with bluetongue virus serotype 8. Med. Vet. Entomol. 2011, 25, 320–326. [Google Scholar] [CrossRef]
- Del Rio López, R.; Miranda, M.A.; Paredes-Esquivel, C.; Lucientes, J.; Calvete, C.; Estrada, R.; Venter, G.J. Recovery rates of bluetongue virus serotypes 1, 2, 4 and 8 Spanish strains from orally infected Culicoides imicola in South Africa. Med. Vet. Entomol. 2012, 26, 162–167. [Google Scholar] [CrossRef] [Green Version]
- Ruder, M.G.; Howerth, E.W.; Stallknecht, D.E.; Allison, A.B.; Carter, D.L.; Drolet, B.S.; Klement, E.; Mead, D.G. Vector competence of Culicoides sonorensis (Diptera: Ceratopogonidae) to epizootic hemorrhagic disease virus serotype 7. Parasites Vectors 2012, 5, 236. [Google Scholar] [CrossRef] [Green Version]
- Venter, G.J.; Paweska, J.T.; Van Dijk, A.A.; Mellor, P.S.; Tabachnick, W.J. Vector competence of Culicoides bolitinos and C. imicola for South African bluetongue virus serotypes 1, 3 and 4. Med Vet. Entomol. 1998, 12, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Meiswinkel, R. Preliminary wing Picture Atlas of Afrotropical Culicoides. Onderstepoort; Veterinary Institute: Pretoria, South Africa, 1994. [Google Scholar]
- Akey, D.H.; Barnard, D.R. Parity in airborne populations of the biting gnat Culicoides variipennis (Diptera: Ceratopogonidae) in Northeastern Colorado. Environ. Entomol. 1983, 12, 91–95. [Google Scholar] [CrossRef]
- Goffredo, M.; Meiswinkel, R. Entomological surveillance of bluetongue in Italy: Methods of capture, catch analysis and identification of Culicoides biting midges. Vet. Ital. 2004, 40, 260–265. [Google Scholar] [PubMed]
- Sivia, D.S.; Skilling, J. Data Analysis. A Bayesian Tutorial, 2nd ed.; Oxford University Press: Oxford, UK, 1996; p. 241. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | M | |||
|---|---|---|---|---|
| Species | No. of Tested | No. of NP * (%) | No. of Tested | No. of NP * (%) |
| Italy: | ||||
| C. scoticus | 1711 | 820 (47.9) | - | - |
| C. imicola | 1395 | 761 (54.6) | 868 | 75 (8.6) |
| C. obsoletus/scoticus | 1216 | 771 (63.4) | - | - |
| C. obsoletus | 347 | 243 (70.0) | - | - |
| C. circumscriptus | 6 | 0 | - | - |
| C. pulicaris | 16 | 4 (25.0) | - | - |
| C. newsteadi | 5 | 4 (80.0) | - | - |
| C. punctatus | 4 | 1 (25.0) | - | - |
| C. montanus | 3 | 2 (66.7) | - | - |
| Total | 4703 | 2606 (55.4) | 868 | 75 (8.6) |
| South Africa: | ||||
| C. imicola | 788 | 319 (40.5) | 20,617 | 1735 (8.4) |
| C. bolitinos | 147 | 73 (49.7) | 5668 | 826 (14.6) |
| C. leucostictus | 7 | 1 (14.3) | 1136 | 170 (15.0) |
| C. gulbenkiani | - | - | 537 | 27 (5.0) |
| C. magnus | - | - | 427 | 25 (5.9) |
| C. engubandei | - | - | 288 | 45 (15.6) |
| C. zuluensis | 2 | 0 | 280 | 21 (7.5) |
| C. bedfordi | 3 | 1 (33.3) | 269 | 85 (31.6) |
| C. pycnostictus | 12 | 1 (8.3) | 154 | 11 (7.1) |
| C. enderleini | 38 | 27 (71.1) | 147 | 23 (15.6) |
| C. nivosus | 18 | 0 | 105 | 8 (7.6) |
| C. huambensis | - | - | 58 | 1 (1.7) |
| C. coarctatus | - | - | 54 | 2 (3.7) |
| C. subschultzei | - | - | 41 | 1 (2.4) |
| C. onderstepoortensis | - | - | 36 | 1(2.8) |
| C. milnei | - | - | 29 | 0 |
| C. dutoiti | - | - | 26 | 4 (15.4) |
| C. nevilli | - | - | 14 | 2 (14.3) |
| C. schultzei | 1 | 0 | 7 | 0 |
| C. neavei | - | - | 5 | 0 |
| C. expectator | - | - | 4 | 0 |
| C. angolensis | - | - | 1 | 0 |
| C. brucei | - | - | 1 | 0 |
| C. cornutus | - | - | 1 | 0 |
| C. nigripennis group | - | - | 1 | 0 |
| C. similis | - | - | 1 | 0 |
| C. tropicalis | 1 | 0 | - | - |
| Total | 1017 | 422 (41.5) | 29,907 | 2987 (10.0) |
| Total Italy and SA | 5720 | 3028 (52.9) | 30,775 | 3062 (9.9) |
| Feeding | No. of NP * Females Tested | No. of Engorged Females (Feeding Rate %) | No. of Females Surviving 10 Days after Feeding | |
|---|---|---|---|---|
| NP * | P ** | |||
| 1st | 2168 | 767 (35.4) | 468 | 52 |
| 2nd | 468 | 38 (8.1) | 29 | 0 |
| 3rd | 29 | 8 (27.6) | 5 | 0 |
| African Horse Sickness Virus (AHSV) | Bluetongue Virus (BTV) | Epizootic Haemorrhagic Disease Virus (EHDV) | ||||
|---|---|---|---|---|---|---|
| Species | C | M | C | M | C | Total |
| C. imicola | 12 | 49 | 47 | 108 | ||
| C. scoticus | 70 | 18 | 5 | 93 | ||
| C. bolitinos | 7 | 3 | 13 | 23 | ||
| C. obsoletus | 12 | 7 | 1 | 20 | ||
| C. leucostictus | 2 | 2 | ||||
| C. magnus | 1 | 1 | ||||
| Total | 82 | 22 | 77 | 60 | 6 | 247 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goffredo, M.; Quaglia, M.; De Ascentis, M.; d’Alessio, S.G.; Federici, V.; Conte, A.; Venter, G.J. The Absence of Abdominal Pigmentation in Livestock Associated Culicoides following Artificial Blood Feeding and the Epidemiological Implication for Arbovirus Surveillance. Pathogens 2021, 10, 1571. https://doi.org/10.3390/pathogens10121571
Goffredo M, Quaglia M, De Ascentis M, d’Alessio SG, Federici V, Conte A, Venter GJ. The Absence of Abdominal Pigmentation in Livestock Associated Culicoides following Artificial Blood Feeding and the Epidemiological Implication for Arbovirus Surveillance. Pathogens. 2021; 10(12):1571. https://doi.org/10.3390/pathogens10121571
Chicago/Turabian StyleGoffredo, Maria, Michela Quaglia, Matteo De Ascentis, Silvio Gerardo d’Alessio, Valentina Federici, Annamaria Conte, and Gert Johannes Venter. 2021. "The Absence of Abdominal Pigmentation in Livestock Associated Culicoides following Artificial Blood Feeding and the Epidemiological Implication for Arbovirus Surveillance" Pathogens 10, no. 12: 1571. https://doi.org/10.3390/pathogens10121571
APA StyleGoffredo, M., Quaglia, M., De Ascentis, M., d’Alessio, S. G., Federici, V., Conte, A., & Venter, G. J. (2021). The Absence of Abdominal Pigmentation in Livestock Associated Culicoides following Artificial Blood Feeding and the Epidemiological Implication for Arbovirus Surveillance. Pathogens, 10(12), 1571. https://doi.org/10.3390/pathogens10121571