
The Impact of Intraspecies Variability on Growth Rate and Cellular Metabolic Activity of Bacteria Exposed to Rotating Magnetic Field


 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results
2.1. Analysis of the Molecular Diversity between Strains of S. aureus
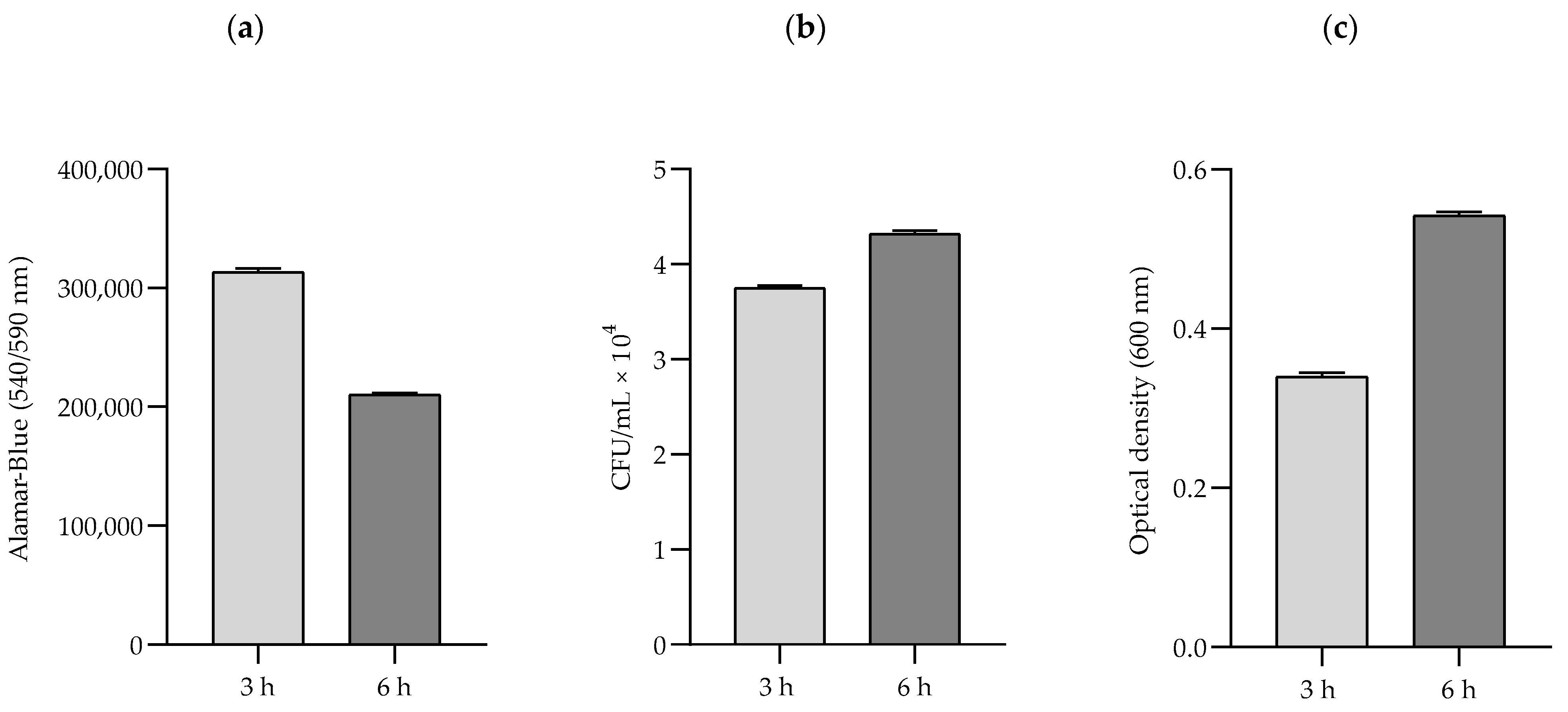
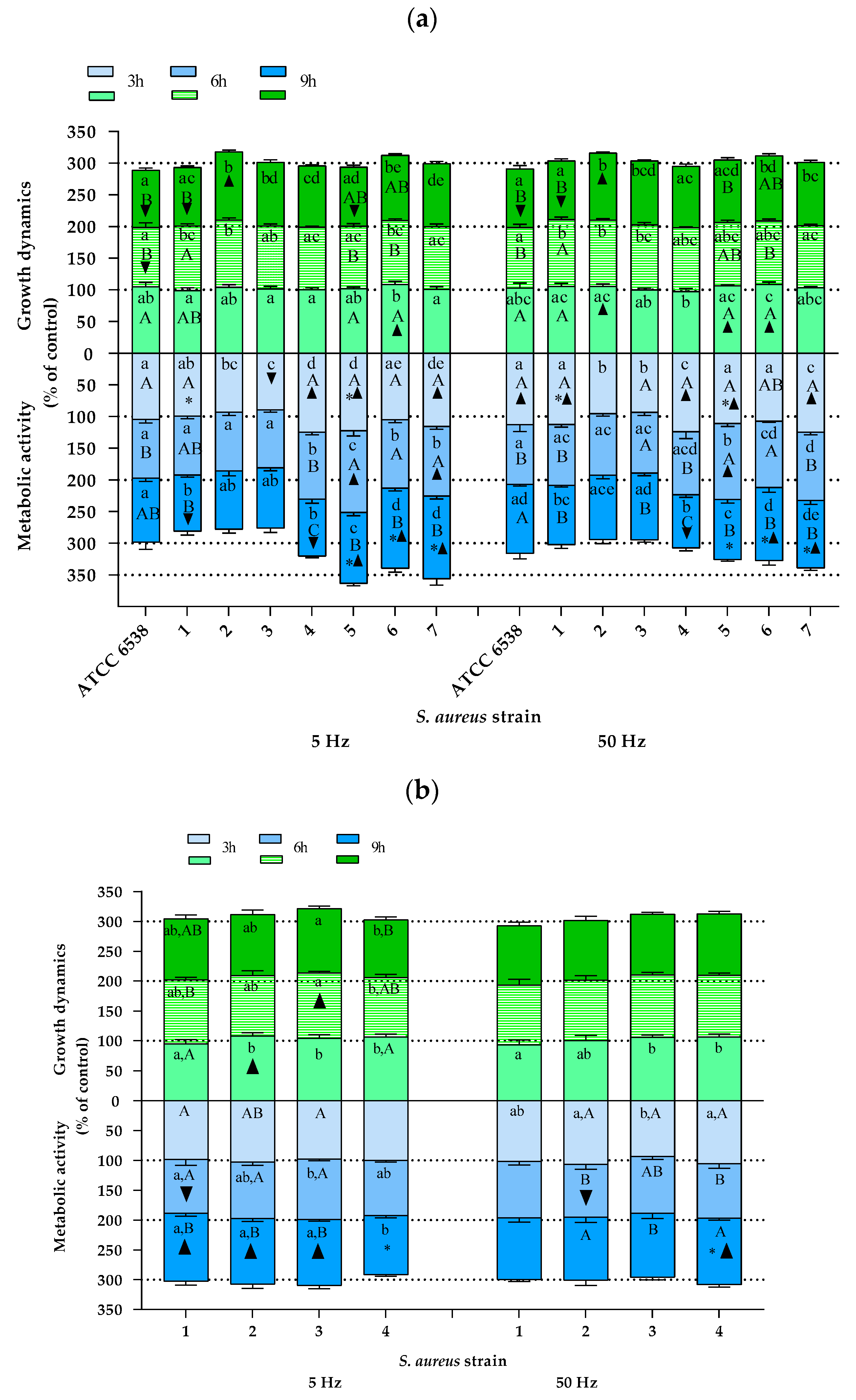
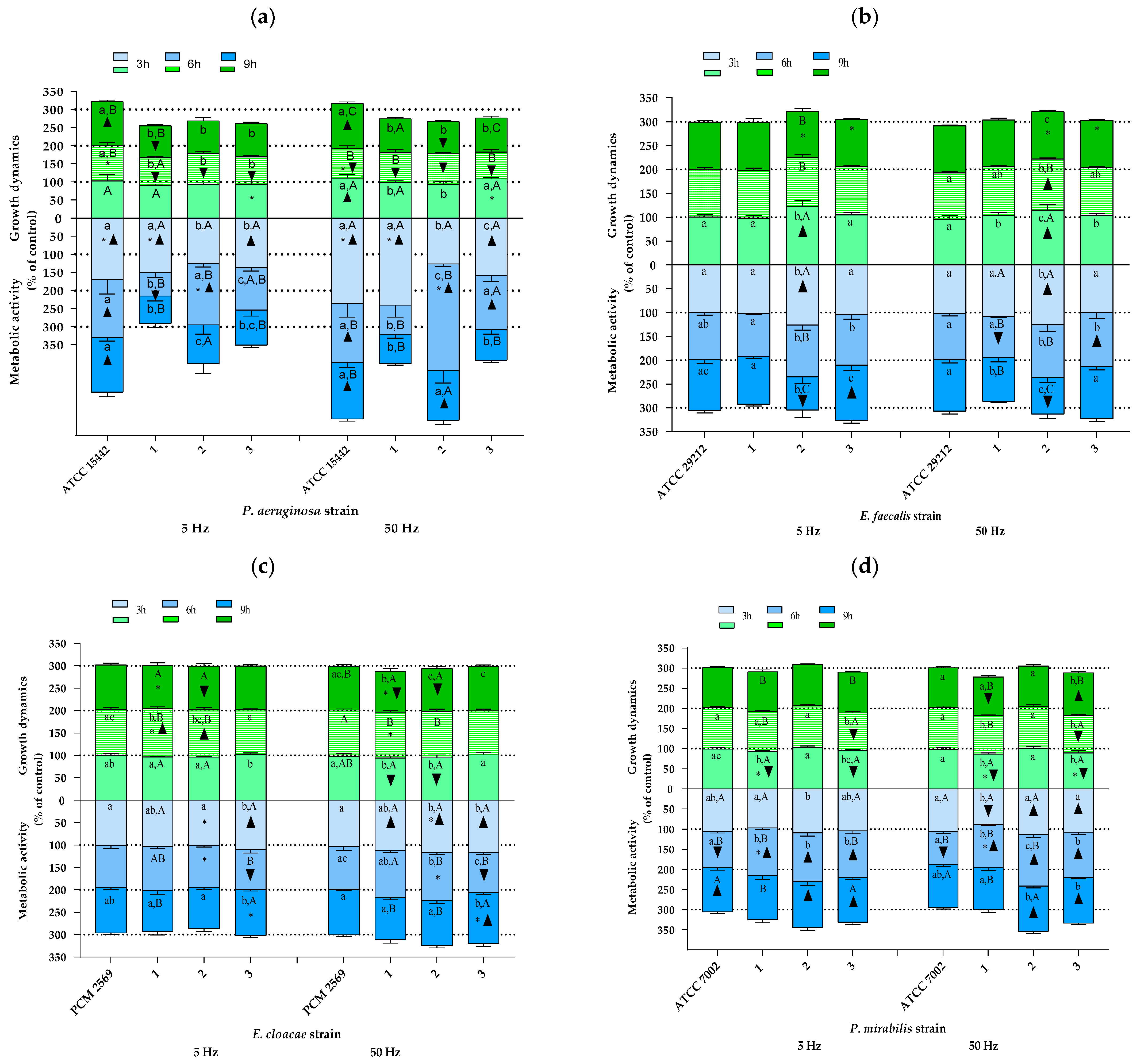
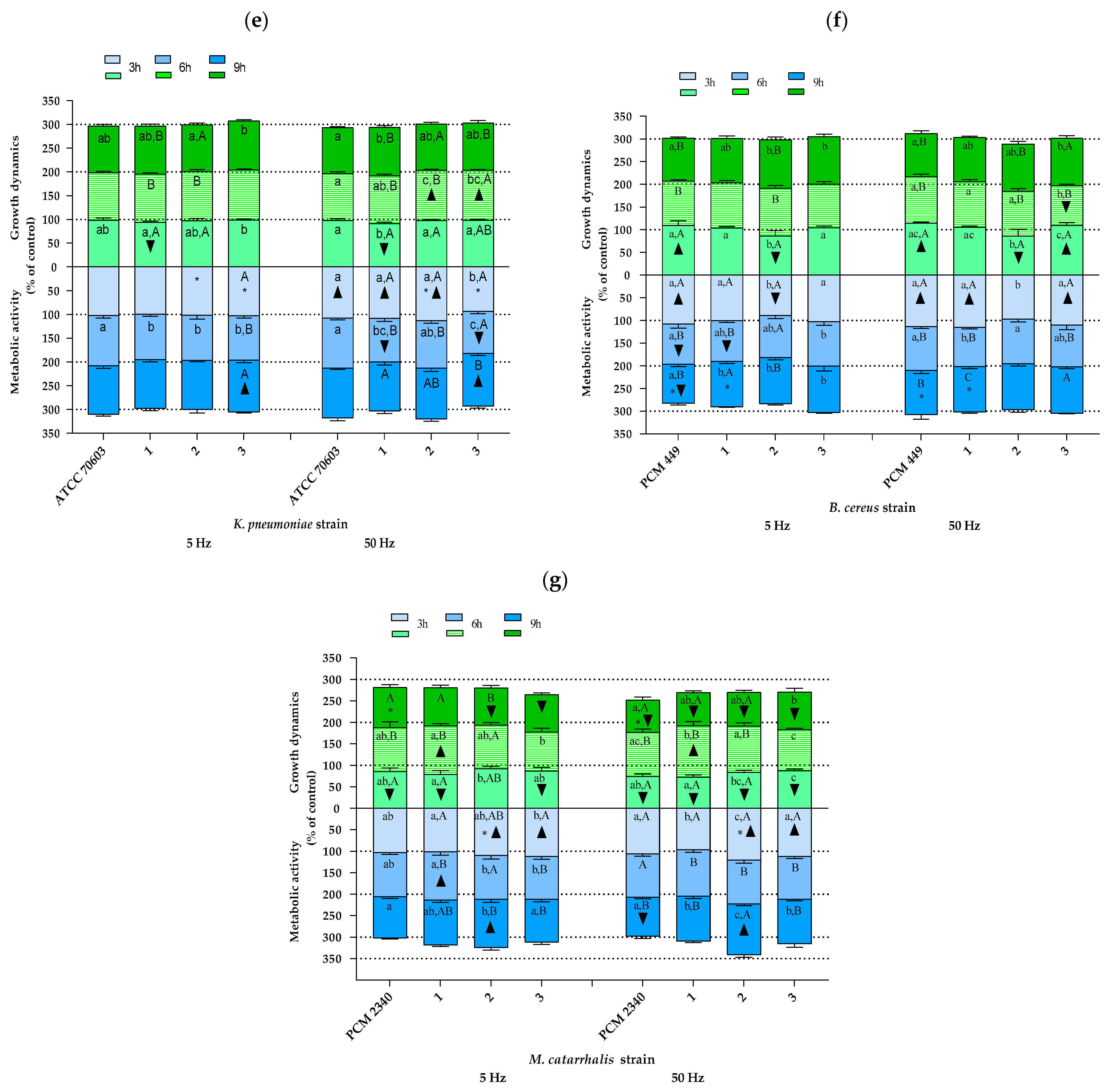
2.2. The Study of Growth Dynamics and Cellular Metabolic Activity
3. Discussion
4. Materials and Methods
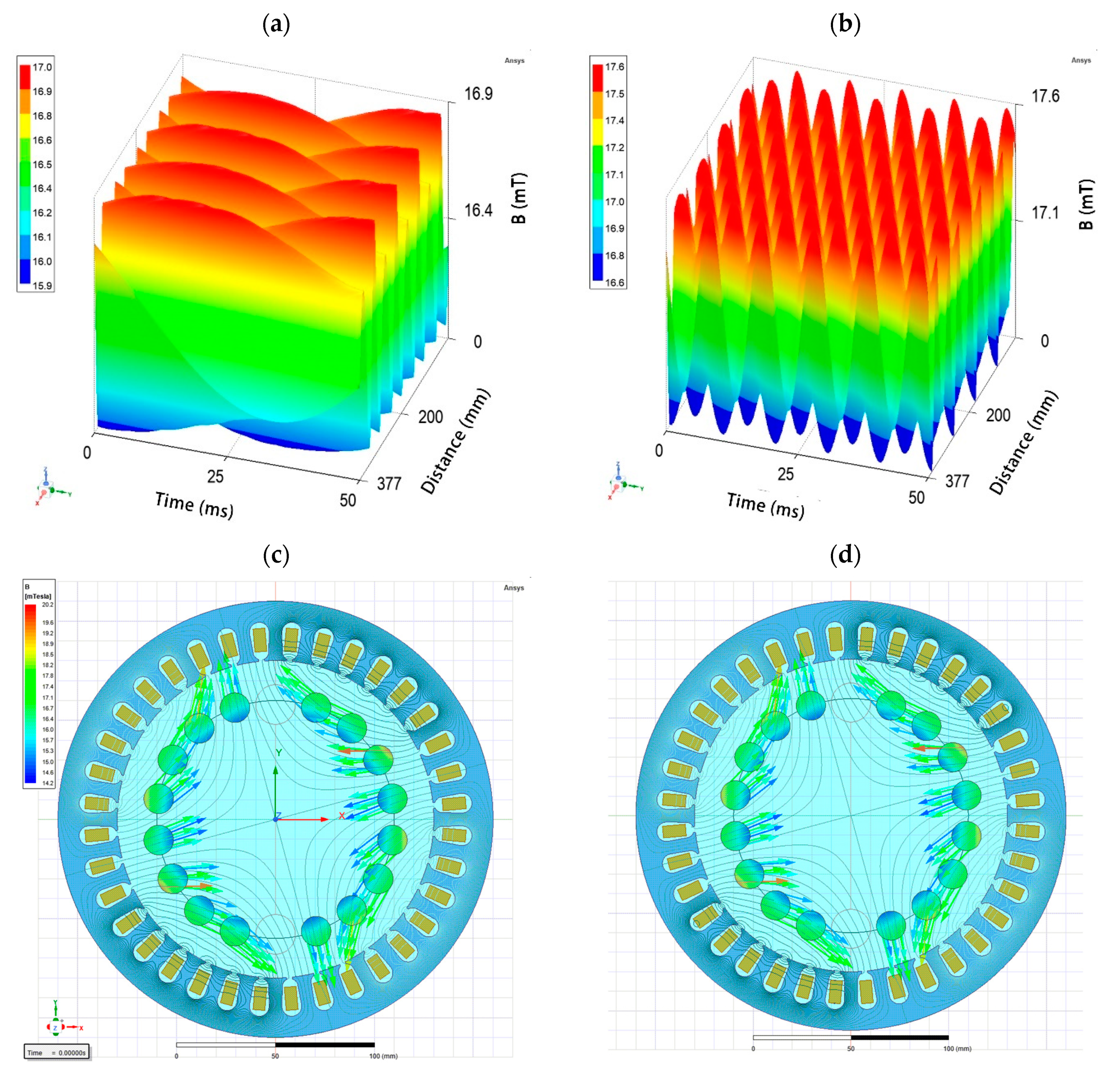
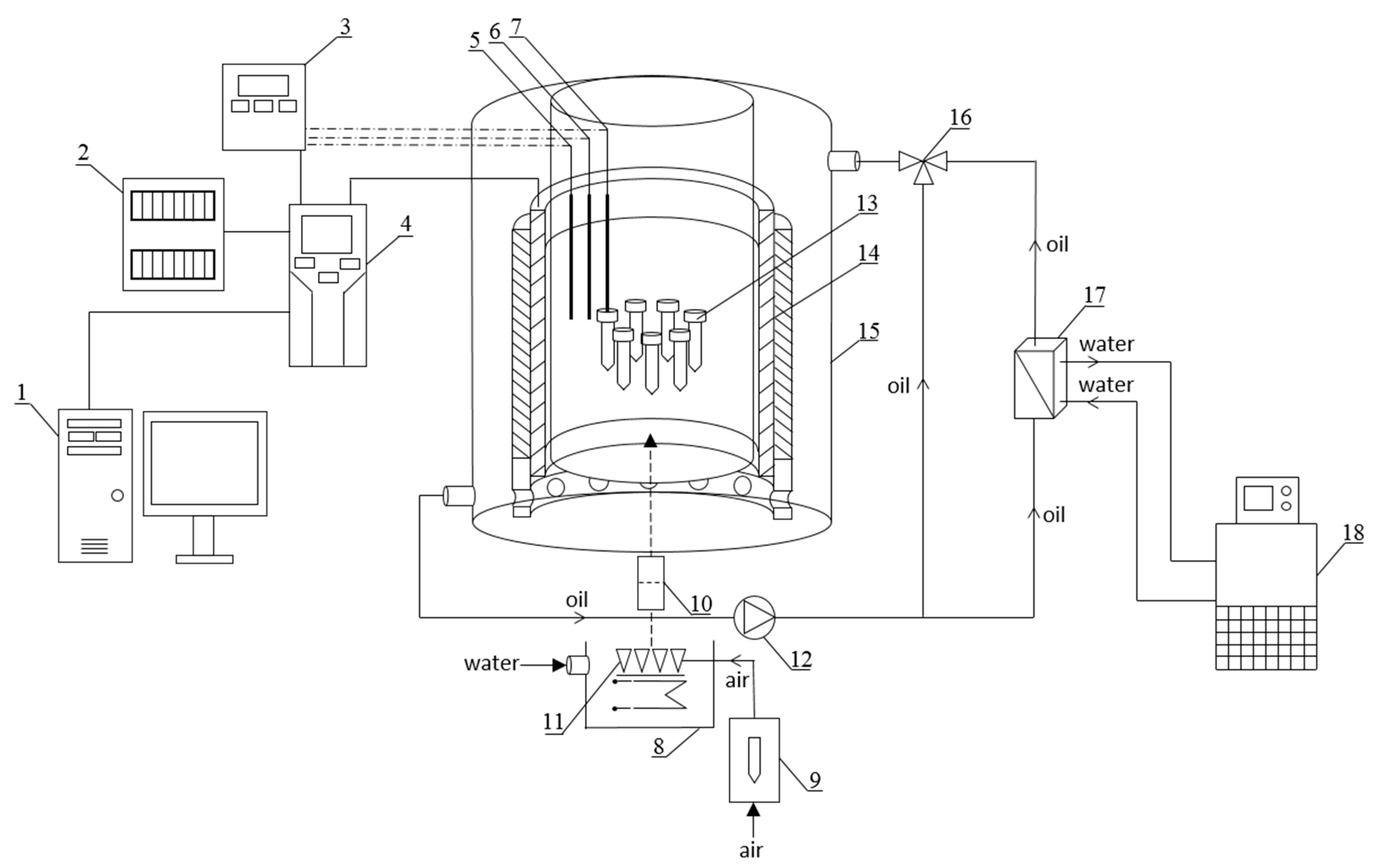
4.1. Experimental Setup
4.2. Microorganisms and Culture Conditions
4.3. Exposure of Bacterial Cultures to the RMF
4.4. Growth Dynamics and Metabolic Activity of Bacterial Cells
4.5. Analysis of Molecular Diversity between Strains of S. aureus
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Redlarski, G.; Lewczuk, B.; Żak, A.; Koncicki, A.; Krawczuk, M.; Piechocki, J.; Jakubiuk, K.; Tojza, P.; Jaworski, J.; Ambroziak, D.; et al. The influence of electromagnetic pollution on living organisms: Historical trends and forecasting changes. BioMed Res. Int. 2015, 2015, 234098. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.K.; Halvorson, H.O.; Sadoff, H.L. Electrolysis as a means of aerating submerged cultures of microorganisms. Appl. Microbiol. 1956, 4, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Sale, A.; Hamilton, W. Effects of high electric fields on microorganismsI. Killing of bacteria and yeasts. Biochim. Biophys. Acta (BBA)-Gen. Subj. 1967, 148, 781–788. [Google Scholar] [CrossRef]
- Sale, A.; Hamilton, W.A. Effects of high electric fields on micro-organisms. Biochim. Biophys. Acta (BBA)-Biomembr. 1968, 163, 37–43. [Google Scholar] [CrossRef]
- Grosman, Z.; Kolár, M.; Tesaríková, E. Effects of static magnetic field on some pathogenic microorganisms. Acta Univ. Palacki. Olomuc. Fac. Med. 1992, 134, 7–9. [Google Scholar] [PubMed]
- Kohno, M.; Yamazaki, M.; Kimura, I.; Wada, M. Effect of static magnetic fields on bacteria: Streptococcus mutans, Staphylococcus aureus, and Escherichia coli. Pathophysiology 2000, 7, 143–148. [Google Scholar] [CrossRef]
- Fojt, L.; Strašák, L.; Vetterl, V.; Šmarda, J. Comparison of the low-frequency magnetic field effects on bacteria Escherichia coli, Leclercia adecarboxylata and Staphylococcus aureus. Bioelectrochemistry 2004, 63, 337–341. [Google Scholar] [CrossRef]
- Potenza, L.; Ubaldi, L.; Sanctis, R.; de Bellis, R.; de Cucchiarini, L.; Dachà, M. Effects of a static magnetic field on cell growth and gene expression in Escherichia coli. Mutat. Res. 2004, 561, 53–62. [Google Scholar] [CrossRef]
- Kermanshahi, R.K.; Sailani, M.R. Effect of static electric field treatment on multiple antibiotic-resistant pathogenic strains of Escherichia coli and Staphylococcus aureus. J. Microbiol. Immunol. Infect. 2005, 38, 394–398. [Google Scholar]
- Strašák, L.; Vetterl, V.; Fojt, L. Effects of 50 Hz Magnetic Fields on the Viability of Different Bacterial Strains. Electrom. Biol. Med. 2005, 24, 293–300. [Google Scholar] [CrossRef]
- Obermeier, A.; Matl, F.D.; Friess, W.; Stemberger, A. Growth inhibition of Staphylococcus aureus induced by low-frequency electric and electromagnetic fields. Bioelectromagnetics 2009, 30, 270–279. [Google Scholar] [CrossRef] [PubMed]
- László, J.; Kutasi, J. Static magnetic field exposure fails to affect the viability of different bacteria strains. Bioelectromagnetics 2010, 31, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Inhan-Garip, A.; Aksu, B.; Akan, Z.; Akakin, D.; Ozaydin, A.N.; San, T. Effect of extremely low frequency electromagnetic fields on growth rate and morphology of bacteria. Int. J. Radiat. Biol. 2011, 87, 1155–1161. [Google Scholar] [CrossRef]
- Bajpai, I.; Saha, N.; Basu, B. Moderate intensity static magnetic field has bactericidal effect on E. coli and S. epidermidis on sintered hydroxyapatite. J. Biomed. Mater. Res. 2012, 100, 1206–1217. [Google Scholar] [CrossRef] [PubMed]
- Filipič, J.; Kraigher, B.; Tepuš, B.; Kokol, V.; Mandic-Mulec, I. Effects of low-density static magnetic fields on the growth and activities of wastewater bacteria Escherichia coli and Pseudomonas putida. Bioresour. Technol. 2012, 120, 225–232. [Google Scholar] [CrossRef]
- Segatore, B.; Setacci, D.; Bennato, F.; Cardigno, R.; Amicosante, G.; Iorio, R. Evaluations of the effects of extremely low-frequency electromagnetic fields on growth and antibiotic susceptibility of Escherichia coli and Pseudomonas aeruginosa. Int. J. Microbiol. 2012, 2012, 587293. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fijalkowski, K.; Nawrotek, P.; Struk, M.; Kordas, M.; Rakoczy, R. The effects of rotating magnetic field on growth rate, cell metabolic activity and biofilm formation by Staphylococcus aureus and Escherichia coli. J. Magn. 2013, 18, 289–296. [Google Scholar] [CrossRef]
- Nawrotek, P.; Fijałkowski, K.; Struk, M.; Kordas, M.; Rakoczy, R. Effects of 50 Hz rotating magnetic field on the viability of Escherichia coli and Staphylococcus aureus. Electrom. Biol. Med. 2014, 33, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Bayır, E.; Bilgi, E.; Şendemir-Ürkmez, A.; Hameş-Kocabaş, E.E. The effects of different intensities, frequencies and exposure times of extremely low-frequency electromagnetic fields on the growth of Staphylococcus aureus and Escherichia coli O157:H7. Electrom. Biol. Med. 2015, 34, 14–18. [Google Scholar] [CrossRef]
- Fijałkowski, K.; Nawrotek, P.; Struk, M.; Kordas, M.; Rakoczy, R. Effects of rotating magnetic field exposure on the functional parameters of different species of bacteria. Electrom. Biol. Med. 2015, 34, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Tessaro, L.W.E.; Murugan, N.J.; Persinger, M.A. Bacterial growth rates are influenced by cellular characteristics of individual species when immersed in electromagnetic fields. Microbiol. Res. 2015, 172, 26–33. [Google Scholar] [CrossRef]
- Mousavian-Roshanzamir, S.; Makhdoumi-Kakhki, A. The inhibitory effects of static magnetic field on Escherichia coli from two different sources at short exposure time. Rep. Biochem. Mol. Biol. 2017, 5, 112–116. [Google Scholar]
- Salmen, S.H.; Alharbi, S.A.; Faden, A.A.; Wainwright, M. Evaluation of effect of high frequency electromagnetic field on growth and antibiotic sensitivity of bacteria. Saudi J. Biol. Sci. 2018, 25, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Konopacki, M.; Rakoczy, R. The analysis of rotating magnetic field as a trigger of Gram-positive and Gram-negative bacteria growth. Biochem. Eng. J. 2019, 141, 259–267. [Google Scholar] [CrossRef]
- Masood, S.; Saleem, I.; Smith, D.; Chu, W.-K. Growth pattern of magnetic field-treated bacteria. Curr. Microbiol. 2020, 77, 194–203. [Google Scholar] [CrossRef]
- Piatti, E.; Cristina Albertini, M.; Baffone, W.; Fraternale, D.; Citterio, B.; Piera Piacentini, M.; Dachà, M.; Vetrano, F.; Accorsi, A. Antibacterial effect of a magnetic field on Serratia marcescens and related virulence to Hordeum vulgare and Rubus fruticosus callus cells. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2002, 132, 359–365. [Google Scholar] [CrossRef]
- Strašák, L.; Vetterl, V.; Šmarda, J. Effects of low-frequency magnetic fields on bacteria Escherichia coli. Bioelectrochemistry 2002, 55, 161–164. [Google Scholar] [CrossRef]
- Gao, W.; Liu, Y.; Zhou, J.; Pan, H. Effects of a strong static magnetic field on bacterium Shewanella oneidensis: An assessment by using whole genome microarray. Bioelectromagnetics 2005, 26, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Poortinga, A.T.; Bos, R.; Busscher, H.J. Lack of effect of an externally applied electric field on bacterial adhesion to glass. Colloids Surf. B Biointerfaces 2001, 20, 189–194. [Google Scholar] [CrossRef]
- Babushkina, I.V.; Borodulin, V.B.; Shmetkova, N.A.; Morrison, V.V.; Usanov, A.D.; Skripal’, A.V.; Usanov, D.A. The influence of alternating magnetic field on Escherichia coli bacterial cells. Pharm. Chem. J. 2005, 39, 398–400. [Google Scholar] [CrossRef]
- Justo, O.R.; Pérez, V.H.; Alvarez, D.C.; Alegre, R.M. Growth of Escherichia coli under extremely low-frequency electromagnetic fields. Appl. Biochem. Biotechnol. 2006, 134, 155–164. [Google Scholar] [CrossRef]
- Cellini, L.; Grande, R.; Di Campli, E.; Di Bartolomeo, S.; Di Giulio, M.; Robuffo, I.; Trubiani, O.; Mariggiò, M.A. Bacterial response to the exposure of 50 Hz electromagnetic fields. Bioelectromagnetics 2008, 29, 302–311. [Google Scholar] [CrossRef]
- Dunca, S.; Creanga, D.-E.; Ailiesei, O.; Nimitan, E. Microorganisms growth with magnetic fluids. J. Magn. Magn. Mater. 2005, 289, 445–447. [Google Scholar] [CrossRef]
- Hu, X.; Qiu, Z.; Wang, Y.; She, Z.; Qian, G.; Ren, Z. Effect of ultra-strong static magnetic field on bacteria: Application of Fourier-transform infrared spectroscopy combined with cluster analysis and deconvolution. Bioelectromagnetics 2009, 30, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Violle, C.; Enquist, B.J.; McGill, B.J.; Jiang, L.; Albert, C.H.; Hulshof, C.; Jung, V.; Messier, J. The return of the variance: Intraspecific variability in community ecology. Trends Ecol. Evol. 2012, 27, 244–252. [Google Scholar] [CrossRef]
- Dijkshoorn, L.; Ursing, B.M.; Ursing, J.B. Strain, clone and species: Comments on three basic concepts of bacteriology. J. Med Microbiol. 2000, 49, 397–401. [Google Scholar] [CrossRef]
- Tenover, F.C.; Arbeit, R.D.; Goering, R.V.; Mickelsen, P.A.; Murray, B.E.; Persing, D.H.; Swaminathan, B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: Criteria for bacterial strain typing. J. Clin. Microbiol. 1995, 33, 2233–2239. [Google Scholar] [CrossRef]
- Martínez-Carranza, E.; Barajas, H.; Alcaraz, L.-D.; Servín-González, L.; Ponce-Soto, G.-Y.; Soberón-Chávez, G. Variability of Bacterial Essential Genes Among Closely Related Bacteria: The case of Escherichia coli. Front. Microbiol. 2018, 9, 1059. [Google Scholar] [CrossRef]
- Nawrotek, P.; Czernomysy-Furowicz, D.; Borkowski, J.; Fijałkowski, K.; Pobucewicz, A. The effect of auto-vaccination therapy on the phenotypic variation of one clonal type of Staphylococcus aureus isolated from cows with mastitis. Vet. Microbiol. 2012, 155, 434–437. [Google Scholar] [CrossRef]
- Valle, A.; Zanardini, E.; Abbruscato, P.; Argenzio, P.; Lustrato, G.; Ranalli, G.; Sorlini, C. Effects of low electric current (LEC) treatment on pure bacterial cultures. J. Appl. Microbiol. 2007, 103, 1376–1385. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing. Available online: https://eucast.org/ (accessed on 28 February 2021).
- Clinical and Laboratory Standards Institute. Available online: https://clsi.org/ (accessed on 28 February 2021).
- Kong, H.; Yu, F.; Zhang, W.; Li, X.; Wang, H. Molecular epidemiology and antibiotic resistance profiles of methicillin-resistant Staphylococcus aureus strains in a tertiary hospital in China. Front. Microbiol. 2017, 8, 838. [Google Scholar] [CrossRef]
- Meroni, G.; Soares Filipe, J.F.; Drago, L.; Martino, P.A. Investigation on antibiotic-resistance, biofilm formation and virulence factors in multi drug resistant and non multi drug resistant Staphylococcus pseudintermedius. Microorganisms 2019, 7, 702. [Google Scholar] [CrossRef]
- Mulyukin, A.L.; Suzina, N.E.; Mel’nikov, V.G.; Gal’chenko, V.F.; El’-Registan, G.I. Dormant state and phenotypic variability of Staphylococcus aureus and Corynebacterium pseudodiphtheriticum. Microbiology 2014, 83, 149–159. [Google Scholar] [CrossRef]
- Libberton, B.; Coates, R.E.; Brockhurst, M.A.; Horsburgh, M.J. Evidence that intraspecific trait variation among nasal bacteria shapes the distribution of Staphylococcus aureus. Infect. Immun. 2014, 82, 3811–3815. [Google Scholar] [CrossRef]
- van Gestel, J.; Bareia, T.; Tenennbaum, B.; Dal Co, A.; Guler, P.; Aframian, N.; Puyesky, S.; Grinberg, I.; D’Souza, G.G.; Erez, Z.; et al. Short-range quorum sensing controls horizontal gene transfer at micron scale in bacterial communities. Nat. Commun. 2021, 12, 2324. [Google Scholar] [CrossRef]
- Suresh, S.; Alva, P.P.; Premanath, R. Modulation of quorum sensing-associated virulence in bacteria: Carbohydrate as a key factor. Arch. Microbiol. 2021, 203, 1881–1890. [Google Scholar] [CrossRef]
- Pisithkul, T.; Schroeder, J.W.; Trujillo, E.A.; Yeesin, P.; Stevenson, D.M.; Chaiamarit, T.; Coon, J.J.; Wang, J.D.; Amador-Noguez, D. Metabolic Remodeling during biofilm development of Bacillus subtilis. MBio 2019, 10, e00623-19. [Google Scholar] [CrossRef]
- Waite, A.J.; Frankel, N.W.; Emonet, T. Behavioral variability and phenotypic diversity in bacterial chemotaxis. Annu. Rev. Biophys. 2018, 47, 595–616. [Google Scholar] [CrossRef]
- Magana, M.; Sereti, C.; Ioannidis, A.; Mitchell, C.A.; Ball, A.R.; Magiorkinis, E.; Chatzipanagiotou, S.; Hamblin, M.R.; Hadjifrangiskou, M.; Tegos, G.P. Options and limitations in clinical investigation of bacterial biofilms. Clin. Microbiol. Rev. 2018, 31, e00084-16. [Google Scholar] [CrossRef]
- Luze, H.; Holzer, J.; Nischwitz, S.P.; Kamolz, L.-P. The importance of in vivo biofilm models for clinical practice. Wound Repair Regen. 2020, 28, 578–579. [Google Scholar] [CrossRef]
- Maillard, J.-Y.; Kampf, G.; Cooper, R. Antimicrobial stewardship of antiseptics that are pertinent to wounds: The need for a united approach. JAC Antimicrob. Resist. 2021, 3, dlab027. [Google Scholar] [CrossRef] [PubMed]
- Drozd, R.; Szymańska, M.; Żywicka, A.; Kowalska, U.; Rakoczy, R.; Kordas, M.; Konopacki, M.; Junka, A.F.; Fijałkowski, K. Exposure to non-continuous rotating magnetic field induces metabolic strain-specific response of Komagataeibacter xylinus. Biochem. Eng. J. 2021, 166, 107855. [Google Scholar] [CrossRef]
- Fijałkowski, K.; Masiuk, H.; Czernomysy-Furowicz, D.; Karakulska, J.; Nawrotek, P.; Paszkowska, A.; Struk, M.; Giedrys-Kalemba, S. Superantigen gene profiles, genetic relatedness and biological activity of exosecretions of Staphylococcus aureus isolates obtained from milk of cows with clinical mastitis. Microbiol. Immunol. 2013, 57, 674–683. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Available online: www.cdc.gov (accessed on 30 August 2021).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Species | Intensity | Frequency | Current | Time of Exposure | Field | Biological Effect/Author Conclusions | References |
|---|---|---|---|---|---|---|---|
| S. aureus E. coli | 0.5–4 T | - | - | 30–120 min | SMF | No influence on growth. | [5] |
| S. aureus * S. mutans * E. coli * | 30, 60, 80, 100 mT | - | - | Uncertain | SMF | Ferrite magnet caused strength-dependent inhibitory effect on the growth of S. mutans and S. aureus when cultured under anaerobic conditions. No growth effects were detected on E. coli cultures. | [6] |
| S. aureus FA 812 E. coli strain K12 L. adecarboxylata 2177 | 10 mT | 50 Hz | - | <30 min | LF-EMF | Decrease in the cell viability and inhibitory effect on the growth rate. | [7] |
| E. coli * | 300 mT | - | - | Up to 50 h | SMF | In the standard medium (LB), no differences between the control and exposed culture were observed. In the modified medium (LB + glutamic acid) after 25 h of cultivation, significant growth stimulation under field exposure occurred in comparison with the control. | [8] |
| S. aureus (38 strains) E. coli (38 strains) Strains were analyzed as a whole group | - | 50 Hz | - | 30–150 min | SEF | Inhibitory effect on the growth of Gram-negative E. coli was greater than Gram-positive S. aureus. | [9] |
| S. aureus FA 812 R. erythropolis E. coli strain K12 L. adecarboxylata 2177 P. denitrificans CCM 982 S. paucimobilis | 10 mT | 50 Hz | - | 24 min | MF | The MF caused a decrease in optical densities of bacterial cultures; the effect was higher for rodlike bacteria. | [10] |
| S. aureus ATCC 49230 | 5 mT | 20 Hz | 1.2 A | 24 h | LF-EMF | Decreased number of cells by 37.3% for electric field (E = 588 mV·cm−1) was observed. | [11] |
| S. aureus 52/03 B. circulans B 01115 M. luteus B 01072 P. fluorescens B 01102 S. enteritidis serovar Enteritidis 359/07 S. marcescens 15/2/7/2 E. coli ATCC 35218 | 159.2 mT | - | - | Up to 24 h | H-SMF | No influence on growth. | [12] |
| 477 mT, 12 mT, 2.8 mT | - | - | Up to 24 h | I-SMF | |||
| S. epidermidis ATCC 12228 S. aureus ATCC 25923 E. faecalis ATCC 29212 E. coli ATCC 25922 K. pneumoniae ATCC 4352 P. aeruginosa ATCC 27853 | 0.5 mT | 50 Hz | - | 6 h | ELF-EMF | Inhibitory effect on growth rate. | [13] |
| S. epidermidis ATCC 35984 E. coli ATCC 25922 | 100 mT | - | - | Up to 4 h | MI-SMF | Inhibitory effect on growth. | [14] |
| E. coli (MG1655, MG 1655rpoS:kan mutant, DH5α with pRPO22oBTcr) P. putida Dc27 | 5–50 mT | - | - | Up to 4 h | LD-SMF | Inhibitory effect on growth. | [15] |
| E. coli ATCC 25922 P. aeruginosa ATCC 27853 | 2 mT | 50 Hz | - | 4, 6, 8 h | ELF-EMF | No remarkable differences were found in the rate of bacteria growth comparing exposed groups with control groups. | [16] |
| S. aureus ATCC 43300 E. coli ATCC 8739 | 22–34 mT | 1–50 Hz | - | 60 min | RMF | Stimulation of the growth dynamics and cell metabolic activity. Higher proliferation rate and cell metabolic activity were found for E. coli. | [17] |
| S. aureus FRI 913 S. aureus ATCC 25923 S. aureus ATCC 43300 E. coli O157:H7 (two strains) E. coli E68II/0141 | 30 mT | 50 Hz | - | 150 min | RMF | Stimulatory effect on the growth and metabolic activity of E. coli and S. aureus. | [18] |
| S. aureus ATCC 6538 E. coli O157:H7 | 2–4 mT | 20, 40, 50 Hz | - | 1–6 h | ELF-EMF | Inhibitory effect on the growth rate in exposed cultures. | [19] |
| S. aureus ATCC 43300 S. mutans ATCC 35668 S. xylosus ATCC 29971 E. coli ATCC 8739 A. baumannii ATCC 19606 P. aeruginosa ATCC 10145 S. marcescens ATCC 274 C. sakazakii ATCC 29544 K. oxytoca PCM 2202 | 25–34 mT | 5-50 Hz | - | 60 min | RMF | Increase in the growth and metabolic activity except for A. baumannii and P. aeruginosa. | [20] |
| S. aureus ATCC 25923 S. epidermidis ATCC 14990 S. marcescens ATCC 264 E. coli ATCC 11303 | 250 μT | 6–25 Hz | - | 12 h | ELF-EM | Increased growth rate of S. epidermidis, S. aureus, and E. coli, inhibitory effect on the growth rate of S. marcescens. | [21] |
| E. coli (wild strains) E. coli ATCC 25922 | 2–20 mT | - | - | 0, 15, 30, 45, 60, 75, 90 min | SMF | Inhibitory effect on the growth rate in exposed cultures at 18 and 20 mT. | [22] |
| S. aureus ATCC 29213 S. epidermidis ATCC 25923 P. aeruginosa ATCC 27853 | 900/1800 MHz | - | 12 h | HF-EMF | Exposure of S. epidermidis and S. aureus to EMF decreased bacterial growth, except for S. aureus at 900 MHz at 12 h. Exposure of P. aeruginosa to EMF at 900 MHz reduced growth rate, while 1800 MHz had insignificant effect. | [23] | |
| S. aureus ATCC 43300 E. faecalis ATCC 29212 S. mutans ATCC 35668 E. coli ATCC 8739 S. marcescens ATCC 274 K. oxytoca PCM 2202 | up to 18 mT | 50 Hz | - | 8 h | RMF | Increased growth and metabolic activity of Gram-positive bacteria (up to 25%) and inhibited proliferation of Gram-negative bacteria (up to 17%) (with the exception of S. marcescens, no statistical differences were observed). | [24] |
| S. aureus 155554A S. epidermidis 155556A E. coli 155065A P. aeruginosa 155250A | 0.3–0.5 mT; 7.5 mT; 0.5 mT; 0.05–0.5 mT | - | - | 24–36 h | B-MF; RM-MF; U-MF; O-MF | Different MFs affect the growth pattern of bacteria differently, depending on the bacterial species. | [25] |
| Bacterial Species | Positive Effect (Stimulating Effect) | Negative Effect (Inhibitory Effect) | No Influence |
|---|---|---|---|
| Gram-positive cocci | |||
| E. faecalis | Konopacki and Rakoczy, 2019 [24] RMF | Inhan-Garip et al., 2011 [13] ELF-EMF | |
| S. aureus | Fijałkowski et al., 2013 [17] RMF Nawrotek et al., 2014 [18] RMF Fijałkowski et al., 2015 [20] RMF Tessaro et al., 2015 [21] ELF-EMF Konopacki and Rakoczy, 2019 [24] RMF | Kohno et al., 2000 [6] SMF Fojt et al., 2004 [7] LF-EMF Kermanshahi et al., 2005 [9] SEF Strašák et al., 2005 [10] MF Inhan-Garip et al., 2011 [13] ELF-EMF Bayir et al., 2015 [19] ELF-EMF Masood et al., 2020 [25] SMF/O-MF Obeimeier et al., 2009 [11] LF-EMF | Grosman et al., 1992 [5] SMF Lászlo and Kutasi, 2010 [12] SMF Salmen et al., 2018 [23] HF-EMF |
| S. epidermidis | Tessaro et al., 2015 [21] ELF-EMF | Inhan-Garip et al., 2011 [13] ELF-EMF Masood et al., 2020 [25] SMF/O-MF Bajpai et al., 2012 [14] MI-SMF | Salmen et al., 2018 [23] HF-EMF |
| S. mutans | Fijałkowski et al., 2015 [20] RMF Konopacki and Rakoczy, 2019 [24] RMF | Kohno et al., 2000 [6] SMF | |
| S. xylosus | Fijałkowski et al., 2015 [20] RMF | ||
| Gram-negative cocci | |||
| P. denitrificans | Strašák et al., 2005 [10] MF | ||
| Gram-positive rod | |||
| B. circulans | Lászlo and Kutasi, 2010 [12] SMF | ||
| Gram-negative rod | |||
| A. baumannii | Fijałkowski et al., 2015 [20] RMF | ||
| E. coli | Fijałkowski et al., 2013 [17] RMF Fijałkowski et al., 2015 [20] RMF Nawrotek et al., 2014 [18] RMF | Fojt et al., 2004 [7] LF-EMF Kermanshahi et al., 2005 [9] SEF Strašák et al., 2005 [10] MF Inhan-Garip et al., 2011 [13] ELF-EMF Bayir et al., 2015 [19] ELF-EMF Mousavian-Roshanzamir and Makhdoumi-Kakhki, 2017 [22] SMF Konopacki and Rakoczy, 2019 [24] RMF Masood et al., 2020 [25] SMF/O-MF Bajpai et al., 2012 [14] MI-SMF Filipič et al., 2012 [15] LD-SMF Tessaro et al., 2015 [21] ELF-EMF | Grosman et al., 1992 [5] SMF Kohno et al., 2000 [6] SMF Lászlo and Kutasi, 2010 [12] SMF Segatore et al., 2012 [16] ELF-EMF |
| K. oxytoca | Fijałkowski et al., 2015 [20] RMF | Konopacki and Rakoczy, 2019 [24] RMF | |
| K. pneumoniae | Inhan-Garip et al., 2011 [13] ELF-EMF | ||
| P. aeruginosa | Inhan-Garip et al., 2011 [13] ELF-EMF Fijałkowski et al., 2015 [20] RMF Salmen et al., 2018 [23] HF-EMF Masood et al., 2020 [25] SMF/O-MG | Segatore et al., 2012 [16] ELF-EMF | |
| P. fluorescens | Lászlo and Kutasi, 2010 [12] SMF | ||
| P. putida | Filipič et al., 2012 [15] LD-SMF | ||
| S. enteritidis | Lászlo and Kutasi, 2010 [12] SMF | ||
| S. marcescens | Fijałkowski et al., 2015 [20] RMF | Tessaro et al., 2015 [21] ELF-EMF | Lászlo and Kutasi, 2010 [12] SMF Konopacki and Rakoczy, 2019 [24] RMF |
| Phenotypic Pattern | Strain Number | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | ATCC 6538 | |
| dRIB | − | − | − | − | − | − | + | − |
| NOVO | − | + | − | − | − | − | + | − |
| ILATk | + | − | + | − | − | + | + | + |
| O129R | + | + | + | + | + | + | + | − |
| LAC | − | − | + | − | − | − | − | − |
| NAG | + | + | + | − | + | + | + | + |
| BGAL | + | − | + | − | + | − | − | − |
| AMAN | − | + | − | − | − | − | − | − |
| MBdG | + | − | − | + | + | + | + | − |
| AGLU | + | + | − | + | + | + | + | − |
| dGAL | + | + | + | − | + | − | − | + |
| ADH2s | + | − | − | − | − | − | − | + |
| Gram Staining | Shape | Spore | Capsule | Motility | Catalase/ Oxidase | Oxidative/ Fermentative | |
|---|---|---|---|---|---|---|---|
| S. aureus | +ve | cocci | −ve | −ve | −ve | +ve/−ve | F |
| P. aeruginosa | −ve | rod | −ve | −ve | +ve | +ve/+ve | O |
| P. mirabilis | −ve | rod | −ve | −ve | +ve | +ve/−ve | F |
| K. pneumoniae | −ve | rod | −ve | +ve | −ve | +ve/−ve | F |
| E. faecalis | +ve | cocci | −ve | −ve | −ve | −ve/−ve | F |
| E. cloacae | −ve | rod | −ve | −ve | +ve | +ve/−ve | F |
| M. catarrhalis | +ve | cocci | −ve | +ve | −ve | +ve/+ve | U |
| B. cereus | +ve | rod with square ends | +ve | −ve | +ve | +/−ve | O |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woroszyło, M.; Ciecholewska-Juśko, D.; Junka, A.; Pruss, A.; Kwiatkowski, P.; Wardach, M.; Fijałkowski, K. The Impact of Intraspecies Variability on Growth Rate and Cellular Metabolic Activity of Bacteria Exposed to Rotating Magnetic Field. Pathogens 2021, 10, 1427. https://doi.org/10.3390/pathogens10111427
Woroszyło M, Ciecholewska-Juśko D, Junka A, Pruss A, Kwiatkowski P, Wardach M, Fijałkowski K. The Impact of Intraspecies Variability on Growth Rate and Cellular Metabolic Activity of Bacteria Exposed to Rotating Magnetic Field. Pathogens. 2021; 10(11):1427. https://doi.org/10.3390/pathogens10111427
Chicago/Turabian StyleWoroszyło, Marta, Daria Ciecholewska-Juśko, Adam Junka, Agata Pruss, Paweł Kwiatkowski, Marcin Wardach, and Karol Fijałkowski. 2021. "The Impact of Intraspecies Variability on Growth Rate and Cellular Metabolic Activity of Bacteria Exposed to Rotating Magnetic Field" Pathogens 10, no. 11: 1427. https://doi.org/10.3390/pathogens10111427
APA StyleWoroszyło, M., Ciecholewska-Juśko, D., Junka, A., Pruss, A., Kwiatkowski, P., Wardach, M., & Fijałkowski, K. (2021). The Impact of Intraspecies Variability on Growth Rate and Cellular Metabolic Activity of Bacteria Exposed to Rotating Magnetic Field. Pathogens, 10(11), 1427. https://doi.org/10.3390/pathogens10111427