

The Fundamental Role of Nutrients for Metabolic Balance and Epigenome Integrity Maintenance



Abstract
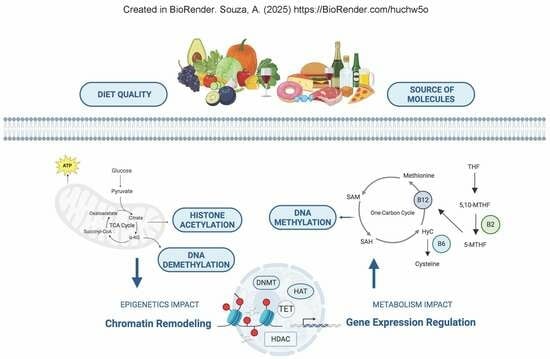
1. Introduction
2. One-Carbon Metabolism and Methyl Groups
3. The Nutritional Emphasis on MTHFR Gene Variants and Differentially Methylated Regions
4. Metabolism and Its Connection to DNA, RNA, and Histone Methylation
5. Importance of TCA Cycle and Generation of Acetyl Groups to Maintenance of Epigenetic Landscape
6. The Importance of Safeguarding Mitochondrial Metabolism and the Epigenome Against Oxidative Damage
7. Polyphenols: Powerful Antioxidants and Key Epigenetic Regulators for Health
8. Astaxanthin: A Multifaceted Carotenoid with Emerging Epigenetic Influence
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 3′ UTR | 3′ untranslated regions |
| 5,10-MTHF | 5,10-methylenetetrahydrofolate |
| ACC1 | Acetyl-coa Carboxylase 1 |
| Acetyl-CoA | Acetyl-coenzyme a |
| ALKBH5 | Alkb homolog 5 |
| AMPK | AMP-activated Protein Kinase |
| ARE | Antioxidant response element |
| ATP | Adenosine triphosphate |
| BHMT | Betaine-homocysteine methyltransferase |
| CBP | CREB-binding protein |
| CBS | Cystathionine β-synthase |
| CpG | Cytosine-phosphate-Guanine |
| DNMT | DNA methyltransferase |
| dTMP | Deoxythymidine 5′-monophosphate |
| dUMP | Deoxyuridine 5′-monophosphate |
| EGCG | Epigallocatechin-3-gallate |
| FADH2 | Flavin adenine dinucleotide |
| FOXO | Forkhead box O |
| FTO | Fat mass and obesity-associated protein |
| H3K18Ac | Histone h3 lysine 18 acetylation |
| HAT | Histone acetyltransferase |
| HCY | Homocysteine |
| HDAC | Histone deacetylase |
| HMT | Histone methyltransferases |
| JHDM | Jumonji C-domain-containing histone demethylases |
| LINE-1 | Long interspersed nuclear element-1 |
| m6A | N6-methyladenosine |
| METTL | Methyltransferase-like |
| MS | Methionine synthase |
| MTHFR | Methylenetetrahydrofolate reductase |
| mTOR | Mechanistic Target of Rapamycin |
| NAD+ | Nicotinamide adenine dinucleotide |
| NADH | Nicotinamide adenine dinucleotide |
| ncRNAs | Non-coding rnas |
| NF-kB | Nuclear factor kappa b |
| NMN | Nicotinamide mononucleotide |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| rDNA | Ribosomal DNA |
| ROS | Reactive oxygen species |
| rRNA | Ribosomal RNA |
| SAH | S-adenosylhomocysteine |
| SAM | S-adenosylmethionine |
| SIRT | Sirtuin |
| SNP | Single nucleotide polymorphism |
| TCA | Tricarboxylic acid cycle |
| TET | Ten-eleven Translocation |
References
- Jones, P.A. Functions of DNA methylation: Islands, start sites, gene bodies and beyond. Nat. Rev. Genet. 2012, 13, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Gómez de Cedrón, M.; Moreno Palomares, R.; Ramírez de Molina, A. Metabolo-epigenetic interplay provides targeted nutritional interventions in chronic diseases and ageing. Front. Oncol. 2023, 13, 1169168. [Google Scholar] [CrossRef] [PubMed]
- Kouzarides, T. Chromatin Modifications and Their Function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Duan, H. The role of m6A RNA methylation in cancer metabolism. Mol. Cancer 2022, 21, 14. [Google Scholar] [CrossRef]
- Allis, C.D.; Jenuwein, T. The molecular hallmarks of epigenetic control. Nat. Rev. Genet. 2016, 17, 487–500. [Google Scholar] [CrossRef]
- Paro, R.; Grossniklaus, U.; Santoro, R.; Wutz, A. Epigenetics and Metabolism. In Epigenetics; Grossniklaus, U., Santoro, R., Wutz, A., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 179–201. [Google Scholar] [CrossRef]
- Chen, C.; Wang, Z.; Qin, Y. Connections between metabolism and epigenetics: Mechanisms and novel anti-cancer strategy. Front. Pharmacol. 2022, 13, 935536. [Google Scholar] [CrossRef]
- Hardy, T.M.; Tollefsbol, T.O. Epigenetic diet: Impact on the epigenome and cancer. Epigenomics 2011, 3, 503–518. [Google Scholar] [CrossRef]
- Casari, G.; Albo, V.; Tini, S.; Sponchioni, M.; Rossi, D. Epigenetic Properties of Compounds Contained in Functional Foods Against Cancer. Biomolecules 2024, 15, 15. [Google Scholar] [CrossRef]
- Zhang, Y.; Kutateladze, T.G. Diet and the epigenome. Nat. Commun. 2018, 9, 3375. [Google Scholar] [CrossRef]
- Fila, M.; Chojnacki, C.; Chojnacki, J.; Blasiak, J. Is an ‘Epigenetic Diet’ for Migraines Justified? The Case of Folate and DNA Methylation. Nutrients 2019, 11, 2763. [Google Scholar] [CrossRef]
- van der Knaap, J.A.; Verrijzer, C.P. Undercover: Gene control by metabolites and metabolic enzymes. Genes Dev. 2016, 30, 2345–2369. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Egervari, G.; Wang, Y.; Berger, S.L.; Lu, Z. Regulation of chromatin and gene expression by metabolic enzymes and metabolites. Nat. Rev. Mol. Cell Biol. 2018, 19, 563–578. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, N.F.P.; Persuhn, D.C.; dos Santos, M.C.L.G. Can Global DNA Methylation Be Influenced by Polymorphisms in Genes Involved in Epigenetic Mechanisms? A Review. Genes 2024, 15, 1504. [Google Scholar] [CrossRef]
- Aissa, A.F.; Kageyama, J.; Sakaki, M.; Tanaka, Y.; Sato, R.; Nishimoto, M.; Ishihara, M. Epigenetic changes induced in mice liver by methionine-supplemented and methionine-deficient diets. Food Chem. Toxicol. 2022, 163, 112938. [Google Scholar] [CrossRef]
- Dai, W.; Chen, D.; Liu, K.; Li, J.; Zhao, C.; Fang, J.; Zhang, X.; Long, C.; Song, Y.; Wang, J.; et al. Epigenetics-targeted drugs: Current paradigms and future challenges. Signal Transduct. Target. Ther. 2024, 9, 332. [Google Scholar] [CrossRef]
- Yaskolka Meir, A.; Tordjman, M.; Tsaban, G.; Levy, S.; Zuk, C.; Bashi, N.; Zilberman-Schapira, L.; Shahar, D.R.; Corem, M.; Tirosh, D.; et al. The effect of polyphenols on DNA methylation-assessed biological age attenuation: The DIRECT PLUS randomized controlled trial. BMC Med. 2023, 21, 364. [Google Scholar] [CrossRef]
- Prasanth, M.I.; Chelikam, V.; Nambiar, N.; Shaik, A.B.; Nair, S.S.; Ponnusamy, S.; Cheruvath, A.; Krishnan, G.; Soman, S.S.; Mohandas, A. Role of Epigenetic Modulation in Neurodegenerative Diseases: Implications of Phytochemical Interventions. Antioxidants 2024, 13, 606. [Google Scholar] [CrossRef]
- Rajendran, P.; Singh, V.K.; Sharma, N.; Singh, A.; Soni, V.; Shukla, Y. Polyphenols as Potent Epigenetics Agents for Cancer. Int. J. Mol. Sci. 2022, 23, 11712. [Google Scholar] [CrossRef]
- Clare, C.E.; Itagaki, R.E.; Fike, M.K.; Rempel, L.A. One-Carbon Metabolism: Linking Nutritional Biochemistry to Epigenetic Programming of Long-Term Development. Annu. Rev. Anim. Biosci. 2019, 7, 263–287. [Google Scholar] [CrossRef]
- van Vliet, M.M.; Vrijkotte, T.G.M.; van Eijsden, M.; de Lau, L.M.; van der Knaap, E.A. One-carbon metabolism as an underlying pathway for placental DNA methylation—A systematic review. Epigenetics 2024, 19, 2318516. [Google Scholar] [CrossRef]
- Choi, S.-W.; Friso, S. Modulation of DNA methylation by one-carbon metabolism: A milestone for healthy aging. Nutr. Res. Pract. 2023, 17, 597. [Google Scholar] [CrossRef]
- Fardous, A.M.; Heydari, A.R. Uncovering the Hidden Dangers and Molecular Mechanisms of Excess Folate: A Narrative Review. Nutrients 2023, 15, 4699. [Google Scholar] [CrossRef]
- Shahzad, K.; Parveen, T.; Fatima, T.; Al-Ghamdi, K.M.; Al-Balawi, S.H.; Al-Ghamdi, H.A.; Ashraf, M.; Elasbali, A.M. A Structured-based Model for the Decreased Activity of Ala222Val and Glu429Ala Methylenetetrahydrofolate Reductase (MTHFR) Mutants. Bioinformation 2013, 9, 929–936. [Google Scholar] [CrossRef]
- Serefidou, M.; Venkatasubramani, A.V.; Imhof, A. The Impact of One Carbon Metabolism on Histone Methylation. Front. Genet. 2019, 10, 764. [Google Scholar] [CrossRef]
- Ducker, G.S.; Rabinowitz, J.D. One-Carbon Metabolism in Health and Disease. Cell Metab. 2017, 25, 27–42. [Google Scholar] [CrossRef]
- Škovierová, H.; Vidomanová, E.; Galfiová, B.; Galfi, P.; Kurilová, K.; Ďuračková, Z.; Šalmíková, A. The Molecular and Cellular Effect of Homocysteine Metabolism Imbalance on Human Health. Int. J. Mol. Sci. 2016, 17, 1733. [Google Scholar] [CrossRef]
- Zarembska, E.; Ślusarczyk, K.; Wrzosek, M. The Implication of a Polymorphism in the Methylenetetrahydrofolate Reductase Gene in Homocysteine Metabolism and Related Civilisation Diseases. Int. J. Mol. Sci. 2023, 25, 193. [Google Scholar] [CrossRef]
- Graydon, J.S.; Lussier, E.; DeSimone, S.P.; Saldivar, J.C. Ethnogeographic Prevalence and Implications of the 677C>T and 1298A>C MTHFR Polymorphisms in US Primary Care Populations. Biomark. Med. 2019, 13, 649–661. [Google Scholar] [CrossRef]
- Pawlik, P.; Chojnowska, E.; Szuflińska, J.; Konopka, W.; Karczmarz, P.; Skoneczny, M.; Cygan, E.; Kosiorek, E.; Błasiak, J.; Moczulska, M.; et al. Common Variants in One-Carbon Metabolism Genes (MTHFR, MTR, MTHFD1) and Depression in Gynecologic Cancers. Int. J. Mol. Sci. 2023, 24, 12574. [Google Scholar] [CrossRef] [PubMed]
- Weiner, A.S.; Khankin, E.; Anpilogova, I.I.; Anpilogov, K.V.; Shvarts, D.A. Methylenetetrahydrofolate reductase C677T and methionine synthase A2756G polymorphisms influence on leukocyte genomic DNA methylation level. Gene 2014, 533, 168–172. [Google Scholar] [CrossRef]
- Burghardt, K.J.; Hultman, C.M.; Karlsson, R.; Öhman, M.; Schalling, M.; Ekström, T.J. DNA Methylation in Schizophrenia Subjects: Gender and MTHFR 677C/T Genotype Differences. Epigenomics 2012, 4, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Wei, J.; Zhang, D.; Liu, S.; Huang, Z.; Zeng, Y.; Wang, J.; Liu, M. Effect of MTHFR Gene Polymorphism Impact on Atherosclerosis via Genome-Wide Methylation. Med. Sci. Monit. 2016, 22, 341–345. [Google Scholar] [CrossRef]
- Mendes, C.C.; Silva, K.A.O.; Lima, M.F.V.; Miranda-Vilela, D.L.; Silva, D.D.; Ribeiro, N.B.; Gualberto, A.F.; Silva, L.D.; Reis, R.C.M.S.; Tavares, J.G.C.R.; et al. One-carbon metabolism and global DNA methylation in mothers of individuals with Down syndrome. Hum. Cell 2021, 34, 1671–1681. [Google Scholar] [CrossRef] [PubMed]
- Mattei, A.L.; Bailly, N.; Meissner, A. DNA methylation: A historical perspective. Trends Genet. 2022, 38, 676–707. [Google Scholar] [CrossRef] [PubMed]
- Tajima, S.; Saitoh, M.; Ueno, K. Domain Structure of the Dnmt1, Dnmt3a, and Dnmt3b DNA Methyltransferases. In DNA Methylation, Volume 2: Transcriptional Regulation and DNA Methylation; Tajima, S., Saitoh, M., Ueno, K., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 45–68. [Google Scholar] [CrossRef]
- Kumari, S.; Singh, S.; Yadav, R.L.; Gupta, G. Chromatin Organization, Epigenetics and Differentiation: An Evolutionary Perspective. In Epigenetics and Chromatin; Springer: Berlin/Heidelberg, Germany, 2013; pp. 3–35. [Google Scholar] [CrossRef]
- Jani, K.S.; Min, J.; Zheng, Y. Histone H3 tail binds a unique sensing pocket in EZH2 to activate the PRC2 methyltransferase. Proc. Natl. Acad. Sci. USA 2019, 116, 8295–8300. [Google Scholar] [CrossRef]
- Briggs, S.D.; Bryk, M.; Strahl, K.B.; Cheung, W.L.; Davie, K.; Allis, C.D.; Winston, F.; Waddell, J. Histone H3 lysine 4 methylation is mediated by Set1 and required for cell growth and rDNA silencing in Saccharomyces cerevisiae. Genes Dev. 2001, 15, 3286–3295. [Google Scholar] [CrossRef]
- Zhao, H.; Chen, T. Tet family of 5-methylcytosine dioxygenases in mammalian development. J. Hum. Genet. 2013, 58, 421–427. [Google Scholar] [CrossRef]
- Ma, C.; Zhang, T.; Li, Z.; Fu, C.; Yang, L.; Luo, J. Ten-eleven translocation proteins (TETs): Tumor suppressors or tumor enhancers? Front. Biosci. 2021, 26, 895–915. [Google Scholar] [CrossRef]
- Chen, Y.; Li, D.; Wang, T.; Luo, T.; Deng, S.; Zhang, H.; Zhang, Y.; Li, L.; Wang, X. Reading the m6A-encoded epitranscriptomic information in development and diseases. Cell Biosci. 2024, 14, 124. [Google Scholar] [CrossRef]
- Corbeski, I.; Eiteneuer, M.; Hauer, D.R.; Meinhart, A. The catalytic mechanism of the RNA methyltransferase METTL3. Elife 2024, 12, e92537. [Google Scholar] [CrossRef]
- Li, P.; Zhang, P.; Wei, F.; Li, Y.; Liu, Q. Epigenetic regulation in female reproduction: The impact of m6A on maternal-fetal health. Cell Death Discov. 2025, 11, 43. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.-Y.; Cao, J.; Li, Y.-F.; Sun, G.-Z.; Zhu, M. Main N6-Methyladenosine Readers: YTH Family Proteins in Cancers. Front. Oncol. 2021, 11, 635329. [Google Scholar] [CrossRef]
- Liu, H.; Wang, Q.; Lin, L.; Liu, K.; Han, M.; Lu, X.; Luo, M.; Li, P.; Luo, H.; Peng, S.; et al. Energy metabolism in health and diseases. Signal Transduct. Target. Ther. 2025, 10, 69. [Google Scholar] [CrossRef]
- Peng, M.; Yin, N.; Chockley, K.; Habowski, A.N.; Delgoffe, G.M.; Magnuson, D.J.; Rabinowitz, J.D.; Edinger, A.L. Aerobic glycolysis promotes T helper 1 cell differentiation through an epigenetic mechanism. Science 2016, 354, 481–484. [Google Scholar] [CrossRef]
- Gagnidze, K.; Pfaff, D.W. Epigenetic Mechanisms: DNA Methylation and Histone Protein Modification. In Neuroscience in the 21st Century; Springer International Publishing: Cham, Switzerland, 2022; pp. 2677–2716. [Google Scholar] [CrossRef]
- Molina-Serrano, D.; Kyriakou, D.; Kirmizis, A. Histone Modifications as an Intersection Between Diet and Longevity. Front. Genet. 2019, 10, 192. [Google Scholar] [CrossRef] [PubMed]
- Lei, I.; Pan, H.; Sun, X.; Shen, J.; Song, R.; Ma, J.; Lu, R.; Zheng, Y.; Yan, C.; Fu, X.; et al. Acetyl-CoA production by specific metabolites promotes cardiac repair after myocardial infarction via histone acetylation. Elife 2021, 10, e60311. [Google Scholar] [CrossRef]
- Bishop, T.R.; Ng, C.; Miller, J.J.; Zhang, Z.; Cvetesic, N.; Huang, C.; Kaelin, W.G.; Chodera, J.D.; Zheng, G.G.; Allis, C.D. Acetyl-CoA biosynthesis drives resistance to histone acetyltransferase inhibition. Nat. Chem. Biol. 2023, 19, 1215–1222. [Google Scholar] [CrossRef] [PubMed]
- Stüve, P.; Zhang, Y.; Giering, K.; Janke, K.; Geffert, H.; Grusche, M.; Munder, M.; Schildberg, F.A.; Freese, C.; Lütjohann, D.; et al. ACC1 is a dual metabolic-epigenetic regulator of Treg stability and immune tolerance. Mol. Metab. 2025, 94, 102111. [Google Scholar] [CrossRef]
- Gnaiger, E. Mitochondrial Pathways and Respiratory Control. An Introduction to OXPHOS Analysis, 5th ed.; Bioenerg Commun: Innsbruck, Austria, 2020. [Google Scholar] [CrossRef]
- Hong, Y.; Zhao, C.; Li, Y.; Liu, C.; Luo, T.; Zhang, Y.; Li, L.; Wang, X. Reactive Oxygen Species Signaling and Oxidative Stress: Transcriptional Regulation and Evolution. Antioxidants 2024, 13, 312. [Google Scholar] [CrossRef]
- Denu, J.M. The Sir2 family of protein deacetylases. Curr. Opin. Chem. Biol. 2005, 9, 431–440. [Google Scholar] [CrossRef]
- Yu, J.; Auwerx, J. The Role of Sirtuins in the Control of Metabolic Homeostasis. Ann. N. Y. Acad. Sci. 2009, 1173, s1. [Google Scholar] [CrossRef]
- Braidy, N.; Villalva, M.G.; van der Kooy, F.; Nazareth, J.; Chang, D.; Jugder, B.E.; Liu, X.; Yi, Z.; Wirth, J.; Sachdev, P. Age Related Changes in NAD+ Metabolism Oxidative Stress and Sirt1 Activity in Wistar Rats. PLoS ONE 2011, 6, e19194. [Google Scholar] [CrossRef] [PubMed]
- Chini, C.C.S.; Sastry, K.S.; Braidy, N.; Chini, E.N. NAD metabolism: Role in senescence regulation and aging. Aging Cell 2024, 23, e13920. [Google Scholar] [CrossRef]
- Bouyahya, A.; Et-Takafi, N.; El Omari, N.; Al-Aboudi, A.R.; El Mskini, A.; El Koraichi, N.; El Ghouizi, A.; Aboulaghras, S.; Bakrim, S.; Jafri, A. The Role of Epigenetic Modifications in Human Cancers and the Use of Natural Compounds as Epidrugs: Mechanistic Pathways and Pharmacodynamic Actions. Biomolecules 2022, 12, 367. [Google Scholar] [CrossRef] [PubMed]
- Bouyahya, A.; Et-Takafi, N.; El Omari, N.; Al-Aboudi, A.R.; El Mskini, A.; El Ghouizi, A.; Aboulaghras, S.; Bakrim, S.; Jafri, A. Bioactive compounds from nature: Antioxidants targeting cellular transformation in response to epigenetic perturbations induced by oxidative stress. Biomed. Pharmacother. 2024, 174, 116432. [Google Scholar] [CrossRef]
- Jomova, K.; Horakova, L. Flavonoids and their role in oxidative stress, inflammation, and human diseases. Chem. Biol. Interact. 2025, 413, 111489. [Google Scholar] [CrossRef]
- Hu, L.; Zhang, S.; Li, X.; Chen, S.; Yang, Y.; Chen, J.; Yang, Z.; Zhou, M.; Yang, C.; Xu, C. Botanical Flavonoids: Efficacy, Absorption, Metabolism and Advanced Pharmaceutical Technology for Improving Bioavailability. Molecules 2025, 30, 1184. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, E.; Bettuzzi, S.; Naponelli, V. The Potential of Epigallocatechin Gallate (EGCG) in Targeting Autophagy for Cancer Treatment: A Narrative Review. Int. J. Mol. Sci. 2022, 23, 6075. [Google Scholar] [CrossRef]
- Huang, C.; Huang, F.; Li, M.; Liu, W.; Pan, S.; Zheng, X. EGCG inhibits protein synthesis, lipogenesis, and cell cycle progression through activation of AMPK in p53 positive and negative human hepatoma cells. Mol. Nutr. Food Res. 2009, 53, 1156–1165. [Google Scholar] [CrossRef]
- Holczer, M.; Szeman, B.; Szabo, A.; Kiss, A.; Zsuga, J.; Szabados, E.; Zvara, A.; Fekete, K.; Csanadi, A.; Szabados, T.; et al. Epigallocatechin-3-Gallate (EGCG) Promotes Autophagy-Dependent Survival via Influencing the Balance of mTOR-AMPK Pathways upon Endoplasmic Reticulum Stress. Oxid. Med. Cell. Longev. 2018, 2018, 6721530. [Google Scholar] [CrossRef]
- Dong, X.; Liu, S.; Sun, Y.; Zhang, R.; Liu, Q.; Wu, X. Epigallocatechin-Gallate: Unraveling Its Protective Mechanisms and Therapeutic Potential. Cell Biochem. Funct. 2025, 43, e70056. [Google Scholar] [CrossRef] [PubMed]
- Vahid, F.; Mohammadshahi, M.; Majidi, M.; Mohammadi-Asl, J. The role dietary of bioactive compounds on the regulation of histone acetylases and deacetylases: A review. Gene 2015, 562, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.K.; Srivastava, R.K.; Sethi, G.; Shandilya, A.; Aggarwal, B.B. Resveratrol Inhibits Growth of Orthotopic Pancreatic Tumors through Activation of FOXO Transcription Factors. PLoS ONE 2011, 6, e25166. [Google Scholar] [CrossRef] [PubMed]
- Poulose, N.; Raju, R. Sirtuin regulation in aging and injury. Biochim. Biophys. Acta Mol. Basis Dis. 2015, 1852, 2442–2455. [Google Scholar] [CrossRef]
- Houston, R.; Li, L.; Wang, S.; Kim, B.; Ragan, M.; Zhang, Y.; Deng, Z.; Chen, S.; Shen, X.; Yang, L.; et al. Acetylation-mediated remodeling of the nucleolus regulates cellular acetyl-CoA responses. PLoS Biol. 2020, 18, e3000981. [Google Scholar] [CrossRef]
- Tang, M.; Zhang, R.; Cao, Y.; Tang, Z. SIRT7: A sentinel of genome stability. Open Biol. 2021, 11, 210047. [Google Scholar] [CrossRef]
- Bosch-Presegué, L.; Vaquero, A. Sirtuin-dependent epigenetic regulation in the maintenance of genome integrity. FEBS J. 2015, 282, 1745–1767. [Google Scholar] [CrossRef]
- You, Y.; Liang, W. SIRT1 and SIRT6: The role in aging-related diseases. Biochim. Biophys. Acta Mol. Basis Dis. 2023, 1869, 166815. [Google Scholar] [CrossRef]
- Sundaram, M.K.; Warrier, A.; Bava, S.V.; Balaram, P.; Singh, V.P.; Hariharan, M. Quercetin modifies 5′CpG promoter methylation and reactivates various tumor suppressor genes by modulating epigenetic marks in human cervical cancer cells. J. Cell. Biochem. 2019, 120, 18357–18369. [Google Scholar] [CrossRef]
- Kashyap, D.; Sharma, A.; Tuli, H.S.; Sak, K.; Kumar, M.; George, S.; Singh, S.; Sharma, U.; Das, K.; Kumar, A. Kaempferol—A dietary anticancer molecule with multiple mechanisms of action: Recent trends and advancements. J. Funct. Foods 2017, 30, 203–219. [Google Scholar] [CrossRef]
- Crespy, V.; Williamson, G. A Review of the Health Effects of Green Tea Catechins in In Vivo Animal Models. J. Nutr. 2004, 134, 3431S–3440S. [Google Scholar] [CrossRef]
- Isemura, M. Catechin in Human Health and Disease. Molecules 2019, 24, 528. [Google Scholar] [CrossRef]
- Diaconeasa, Z.; Sidor, L.C.; Rusu, A.; Leabu, M.; Rugină, D. Anthocyanins, Vibrant Color Pigments, and Their Role in Skin Cancer Prevention. Biomedicines 2020, 8, 336. [Google Scholar] [CrossRef]
- Najjar Khalilabad, S.; Hosseini, S.A.; Tabrizi, A.; Vahid, F. How hesperidin and Hesperetin, as promising food Supplements, combat cardiovascular Diseases: A systematic review from bench to bed. J. Funct. Foods 2024, 120, 106358. [Google Scholar] [CrossRef]
- Pearson, K.J.; Baur, J.A.; Lewis, K.N.; Anderson, R.G.; Bartke, T.A.; Bradford, M.R.; Conklin, B.W.; Csiszar, T.; Dixit, K.B.; Dollins, D.E.; et al. Resveratrol Delays Age-Related Deterioration and Mimics Transcriptional Aspects of Dietary Restriction without Extending Life Span. Cell Metab. 2008, 8, 157–168. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Sundaram, C.; Malani, N.; Ichikawa, H. Curcumin: The indian solid gold. In The Molecular Targets and Therapeutic Uses of Curcumin in Health and Disease; Aggarwal, B.B., Surh, Y.-J., Shishodia, S., Eds.; Springer: Boston, MA, USA, 2007; pp. 1–75. [Google Scholar] [CrossRef]
- Boumezough, K.; Chahdi, H.; Akssira, A.; El Ghouizi, A. The potential health benefits of olive oil polyphenols in metabolic disorders management. PharmaNutrition 2025, 31, 100428. [Google Scholar] [CrossRef]
- Weng, Y.-P.; Lin, H.-Y.; Lo, Y.-L.; Chang, C.-Y.; Chiu, C.-Y.; Pan, H.-L. The inhibitory activity of gallic acid against DNA methylation: Application of gallic acid on epigenetic therapy of human cancers. Oncotarget 2018, 9, 361–374. [Google Scholar] [CrossRef]
- Bayazid, A.B.; Lim, B.O. Therapeutic Effects of Plant Anthocyanin against Alzheimer’s Disease and Modulate Gut Health, Short-Chain Fatty Acids. Nutrients 2024, 16, 1554. [Google Scholar] [CrossRef]
- Dang, Y.; Li, Z.; Yu, F. Recent Advances in Astaxanthin as an Antioxidant in Food Applications. Antioxidants 2024, 13, 879. [Google Scholar] [CrossRef]
- Medoro, A.; Davinelli, S.; Milella, L.; Willcox, B.J.; Allsopp, R.C.; Scapagnini, G.; Willcox, D.C. Dietary Astaxanthin: A Promising Antioxidant and Anti-Inflammatory Agent for Brain Aging and Adult Neurogenesis. Mar. Drugs 2023, 21, 643. [Google Scholar] [CrossRef]
- Shokri-Mashhadi, N.; Tahmasebi, M.; Mohammadi-Asl, J.; Zakerkish, M.; Mohammadshahi, M. The Antioxidant and Anti-Inflammatory Effects of Astaxanthin Supplementation on the Expression of miR-146a and miR-126 in Patients with Type 2 Diabetes Mellitus: A Randomised, Double-Blind, Placebo-Controlled Clinical Trial. Int. J. Clin. Pract. 2021, 75, e14022. [Google Scholar] [CrossRef] [PubMed]
- Chaboksafar, M.; Fakhr, L.; Kheirouri, S.; Alizadeh, M. The Effects of Astaxanthin Supplementation on Expression of MicroRNAs Involved in Cardiovascular Diseases: A Systematic Review of Current Evidence. Int. J. Food Sci. Nutr. 2022, 73, 1019–1029. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Class | Polyphenol | Diet Source | Function in Metabolism and Epigenetics | References |
|---|---|---|---|---|
| Flavonoid | Quercetin | Apples, onions, capers, berries | Quercetin has been implicated in the regulation of DNA methylation, influencing various gene expression profiles linked to inflammatory and cellular stress responses. This flavonoid also exhibits potential to modulate epigenetic enzymes, promoting a favorable genomic environment. | [73] |
| Flavonoid | Kaempferol | Kale, spinach, broccoli, leeks, beans | This flavonoid is known to induce significant changes in DNA methylation patterns and histone acetylation, thus altering the expression of genes involved in cell differentiation and apoptosis, with implications for cancer prevention. | [74] |
| Flavonoid | Catechins | Green tea, black tea, dark chocolate | Catechins have been shown to modulate various histone modifications, particularly enhancing acetylation, which can lead to the upregulation of genes associated with antioxidant activity and downregulation of pro-inflammatory pathways. | [75,76] |
| Flavonoid | Anthocyanins | Berries (blueberries, blackberries), red cabbage, eggplant | These potent antioxidants are thought to induce epigenetic modifications that can counteract oxidative stress and inflammation, facilitating a protective response through modulation of gene expression related to cellular health. | [77] |
| Flavonoid | Hesperidin | Citrus fruits (oranges, lemons), peppermint, berries | Hesperidin plays a significant role in modulating gene expression related to oxidative stress response, potentially influencing signaling pathways crucial for maintaining cellular homeostasis through epigenetic mechanisms. | [78] |
| Non-Flavonoid | Resveratrol | Red wine, grapes, peanuts, berries | Resveratrol exhibits a remarkable capacity to influence histone acetylation, which in turn regulates the expression of genes involved in longevity and metabolic health, highlighting its potential in disease prevention strategies. | [79] |
| Non-Flavonoid | Curcumin | Turmeric, ginger, curry powder, mustard | Known for its multifaceted actions, curcumin can trigger significant epigenetic alterations in histone modification, impacting the expression of genes associated with inflammation, cancer progression, and neuroprotection. | [80] |
| Non-Flavonoid | Oleuropein | Extra virgin olive oil, olives, olive leaf extract | This polyphenol is recognized for its role in modulating key epigenetic factors involved in metabolic pathways, potentially influencing lipid metabolism and inflammatory responses, thus promoting overall health. | [81] |
| Non-Flavonoid | Gallic acid | Black tea, walnuts, grapes, pomegranates | Gallic acid is believed to affect DNA methylation dynamics, particularly in genes related to immune responses and cancer susceptibility, promoting a balanced epigenetic landscape. | [82,83] |
| Non-Flavonoid | Anthocyanidins | Grapes, apples, cocoa, berries | These compounds exhibit the capacity to influence epigenetic regulation concerning antioxidant defense systems, thereby mediating protective effects against oxidative damage and chronic diseases. | [84] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souza, A.P.d.; Marinho, V.; Marques, M.R. The Fundamental Role of Nutrients for Metabolic Balance and Epigenome Integrity Maintenance. Epigenomes 2025, 9, 23. https://doi.org/10.3390/epigenomes9030023
Souza APd, Marinho V, Marques MR. The Fundamental Role of Nutrients for Metabolic Balance and Epigenome Integrity Maintenance. Epigenomes. 2025; 9(3):23. https://doi.org/10.3390/epigenomes9030023
Chicago/Turabian StyleSouza, Ana Paula de, Vitor Marinho, and Marcelo Rocha Marques. 2025. "The Fundamental Role of Nutrients for Metabolic Balance and Epigenome Integrity Maintenance" Epigenomes 9, no. 3: 23. https://doi.org/10.3390/epigenomes9030023
APA StyleSouza, A. P. d., Marinho, V., & Marques, M. R. (2025). The Fundamental Role of Nutrients for Metabolic Balance and Epigenome Integrity Maintenance. Epigenomes, 9(3), 23. https://doi.org/10.3390/epigenomes9030023