
Critical Considerations for Investigating MicroRNAs during Tumorigenesis: A Case Study in Conceptual and Contextual Nuances of miR-211-5p in Melanoma

 and
and
Abstract
1. Introduction
2. The Role of MicroRNAs in Stabilizing Transcriptional Programs
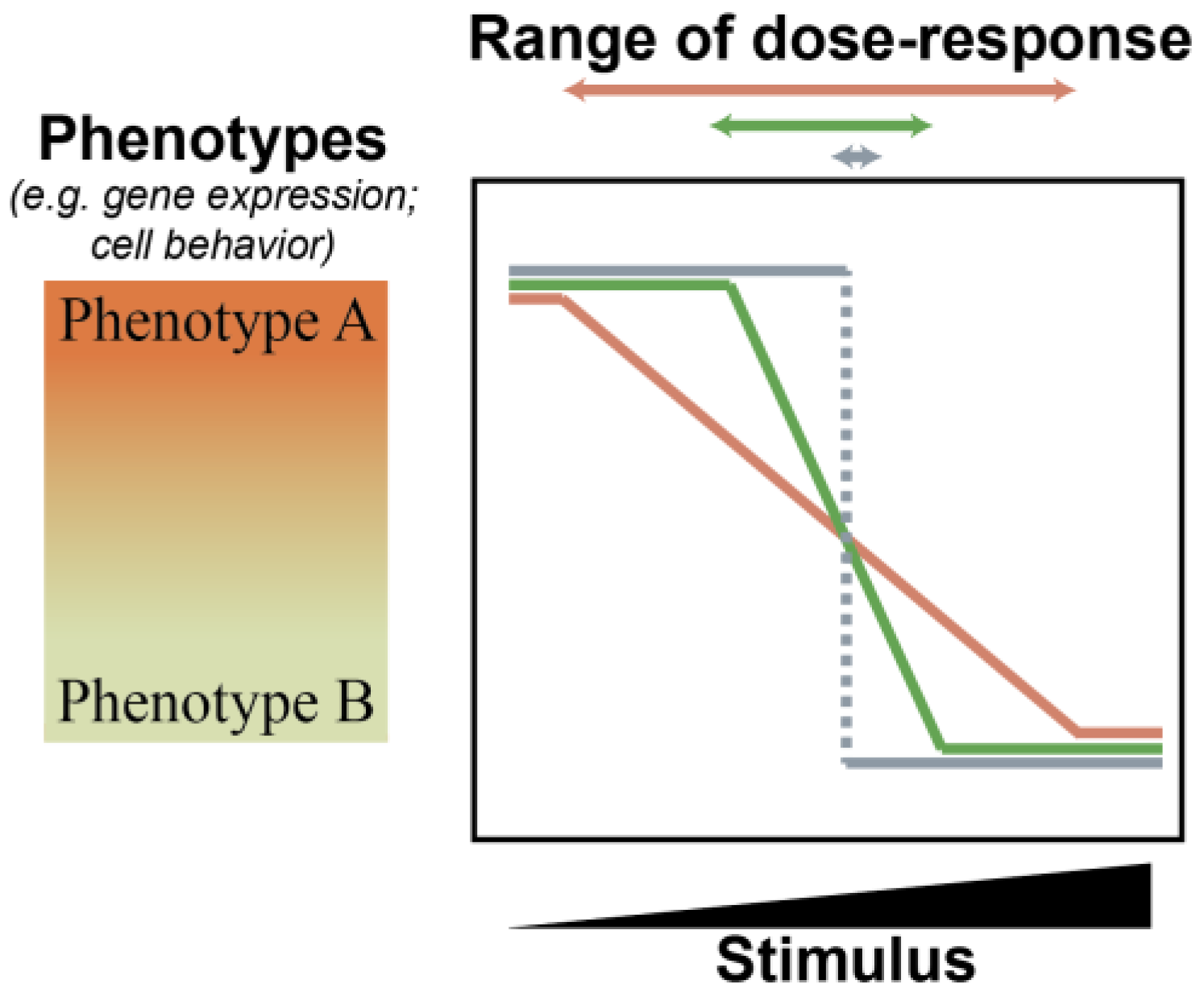
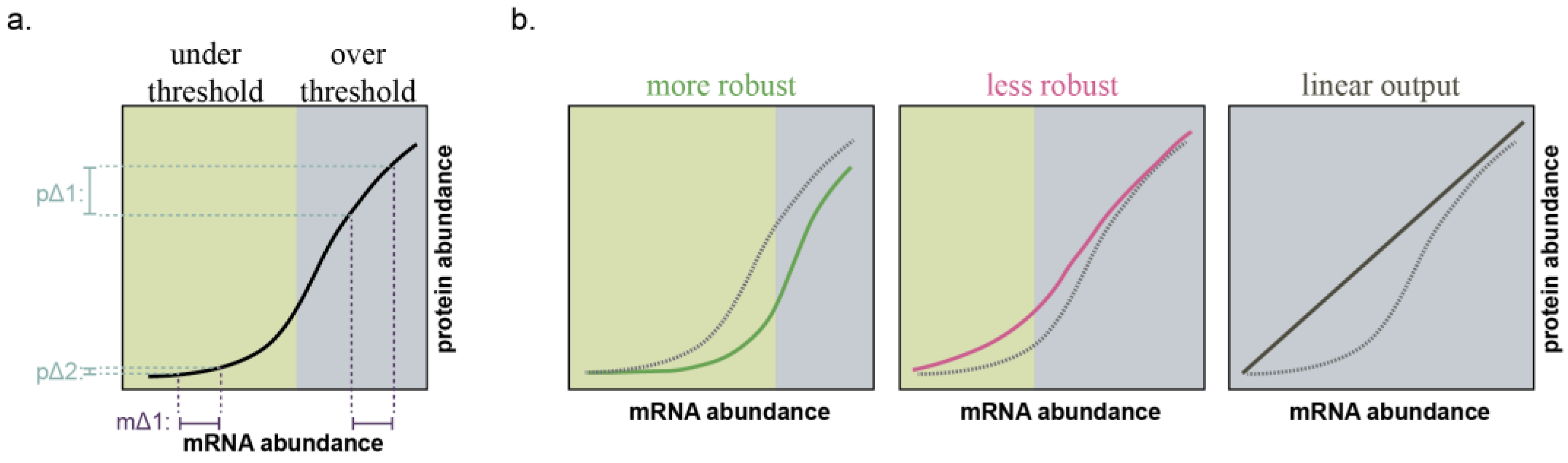
2.1. MicroRNAs Bolster Cell State Robustness
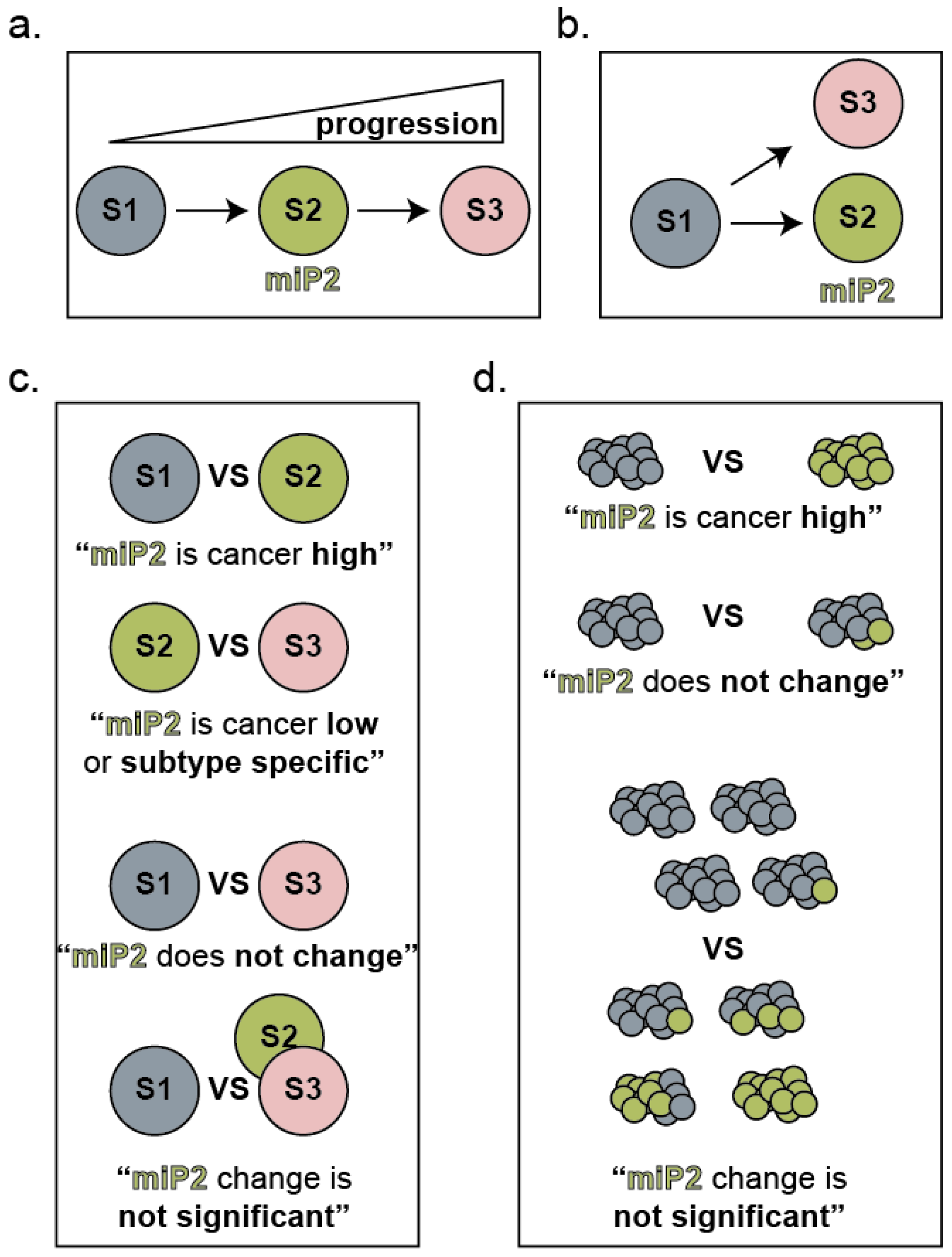
2.2. Considerations for the Role of miRNAs during Tumor Progression
3. MicroRNAs Consistently Associated with Melanoma Progression
3.1. Assessment of Melanoma Cohort Assembly in MicroRNA Profiling Studies
3.2. Cross-Study Concordance of MicroRNAs Associated with Melanocytic Nevi
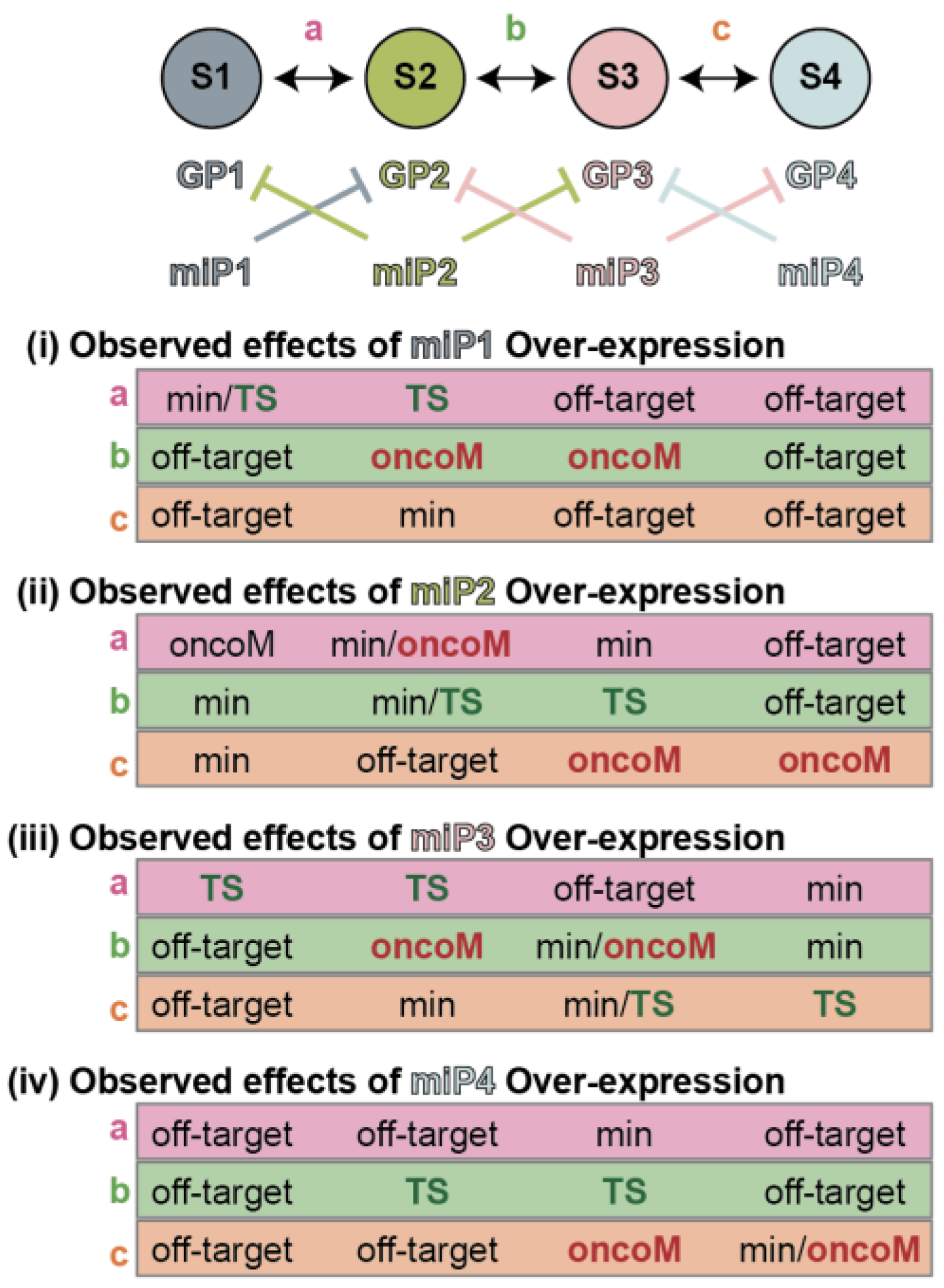
3.3. Increased Variability of miRNA Expression Is Associated with Melanoma Progression
4. miR-211-5p Expression Confers Robustness to Pigmented Cells
4.1. miR-211-5p Is a Consistent Nevus-Associated miRNA
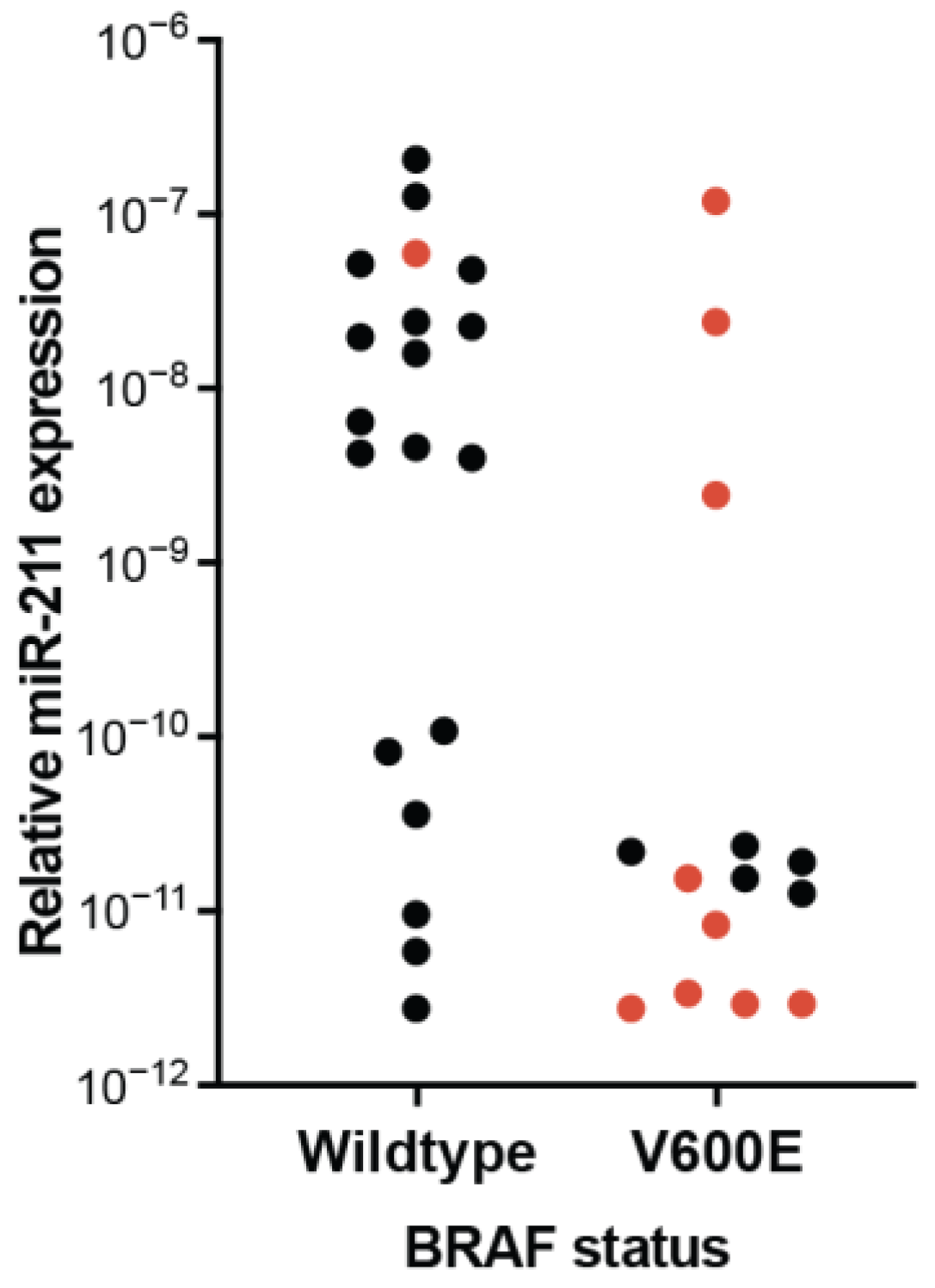
4.2. Experimental Conditions Influence Observed miR-211 Expression and Function
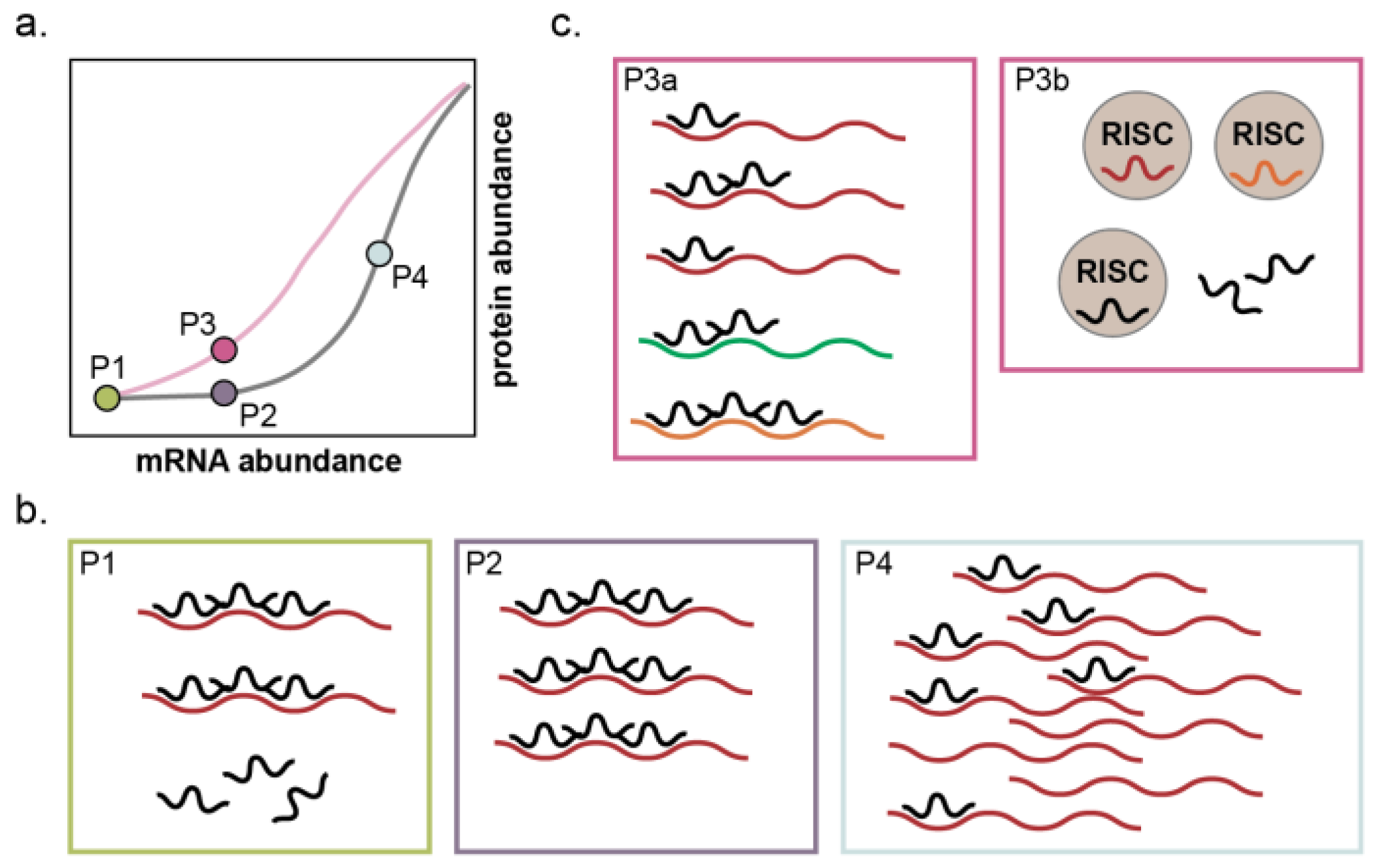
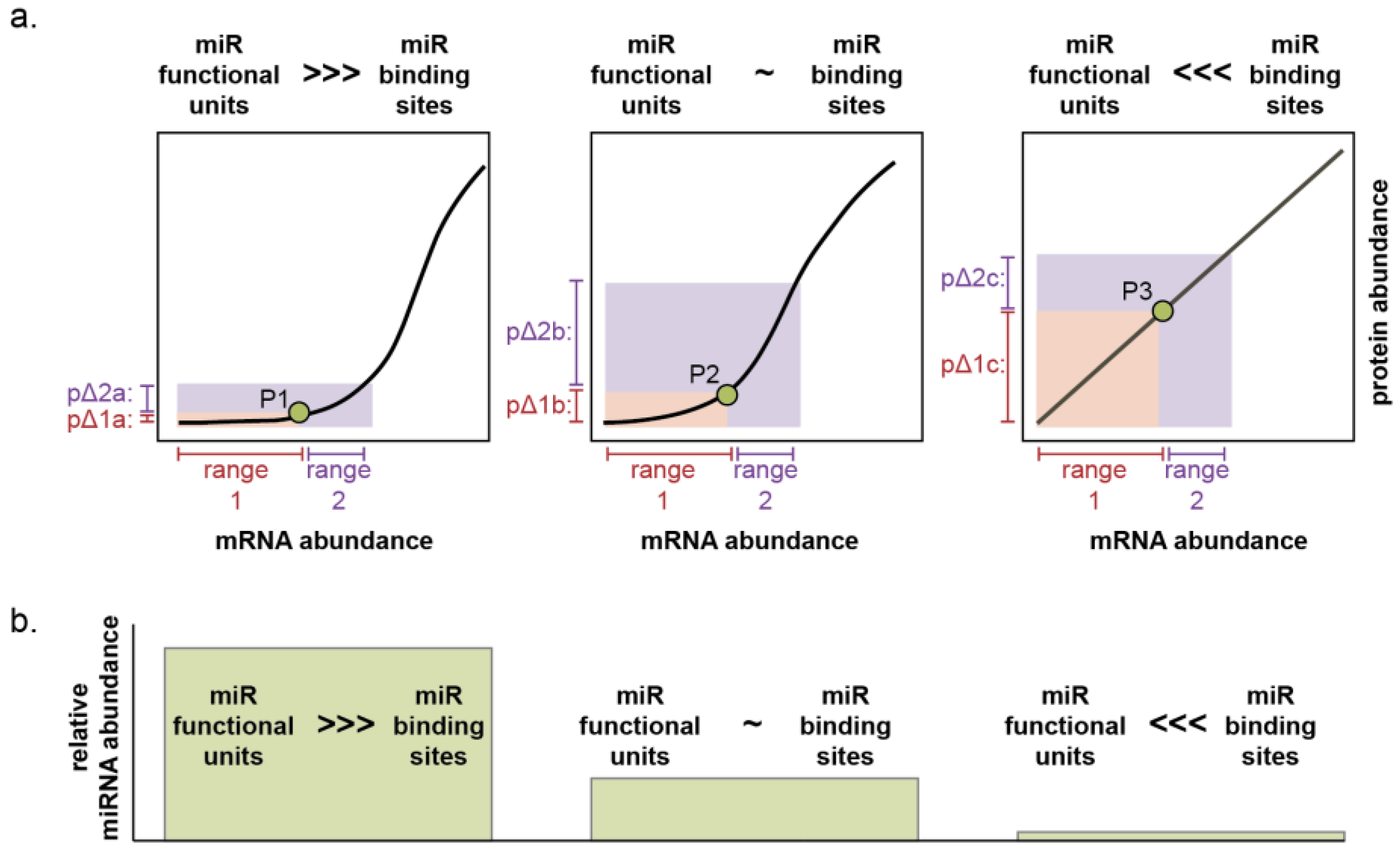
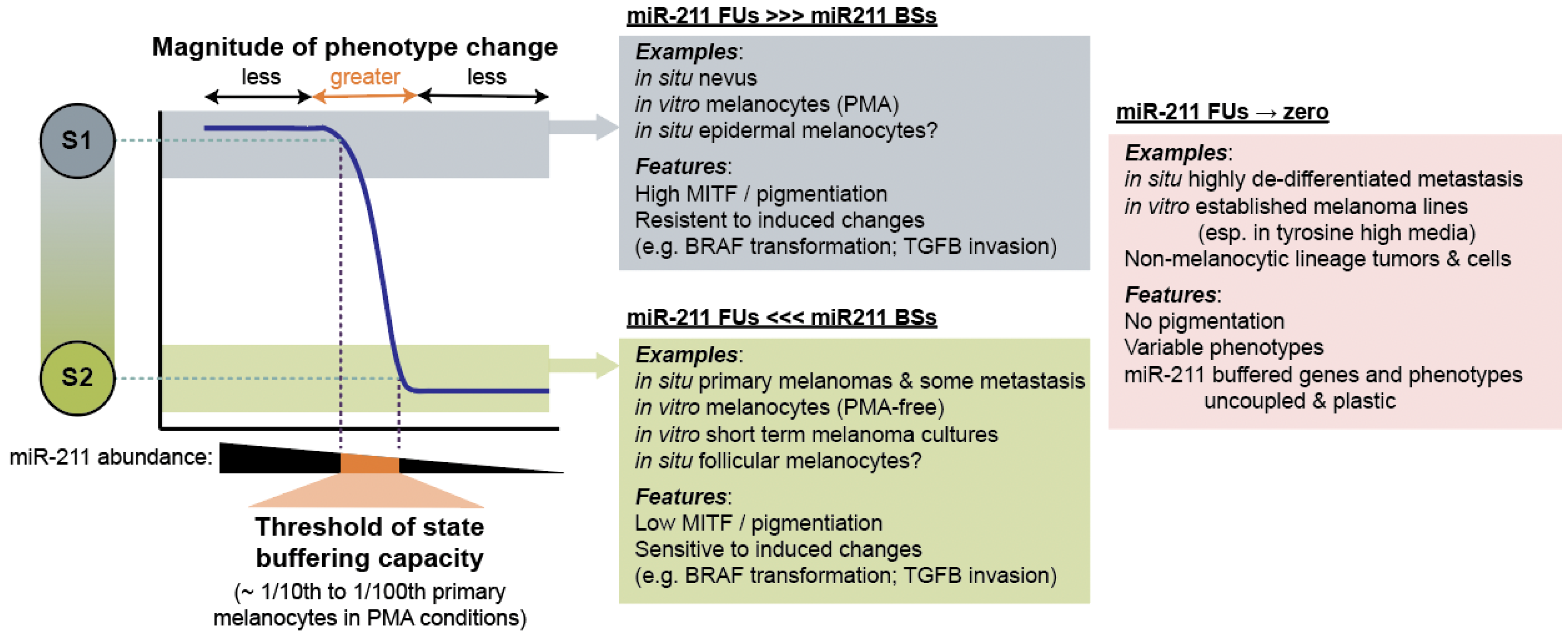
4.3. MicroRNA-211 Establishes Thresholds for Target Gene Expression and Cell Phenotypes
5. Perspectives, Future Directions, and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- The ENCODE Project Consortium. An Integrated Encyclopedia of DNA Elements in the Human Genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef] [PubMed]
- International Human Genome Sequencing Consortium. Finishing the Euchromatic Sequence of the Human Genome. Nature 2004, 431, 931–945. [Google Scholar] [CrossRef] [PubMed]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al. Landscape of Transcription in Human Cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. Elegans Heterochronic Gene Lin-4 Encodes Small RNAs with Antisense Complementarity to Lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Deveson, I.W.; Hardwick, S.A.; Mercer, T.R.; Mattick, J.S. The Dimensions, Dynamics, and Relevance of the Mammalian Noncoding Transcriptome. Trends Genet. 2017, 33, 464–478. [Google Scholar] [CrossRef] [PubMed]
- Guerra-Assunção, J.A.; Enright, A.J. Large-Scale Analysis of MicroRNA Evolution. BMC Genom. 2012, 13, 218. [Google Scholar] [CrossRef]
- Zolotarov, G.; Fromm, B.; Legnini, I.; Ayoub, S.; Polese, G.; Maselli, V.; Chabot, P.J.; Vinther, J.; Styfhals, R.; Seuntjens, E.; et al. MicroRNAs Are Deeply Linked to the Emergence of the Complex Octopus Brain. Sci. Adv. 2022, 8, eadd9938. [Google Scholar] [CrossRef]
- Heimberg, A.M.; Sempere, L.F.; Moy, V.N.; Donoghue, P.C.J.; Peterson, K.J. MicroRNAs and the Advent of Vertebrate Morphological Complexity. Proc. Natl. Acad. Sci. USA 2008, 105, 2946–2950. [Google Scholar] [CrossRef]
- Takuno, S.; Innan, H. Evolution of Complexity in MiRNA-Mediated Gene Regulation Systems. Trends Genet. 2008, 24, 56–59. [Google Scholar] [CrossRef]
- Wheeler, B.M.; Heimberg, A.M.; Moy, V.N.; Sperling, E.A.; Holstein, T.W.; Heber, S.; Peterson, K.J. The Deep Evolution of Metazoan MicroRNAs. Evol. Dev. 2009, 11, 50–68. [Google Scholar] [CrossRef]
- McCreight, J.C.; Schneider, S.E.; Wilburn, D.B.; Swanson, W.J. Evolution of MicroRNA in Primates. PLoS ONE 2017, 12, e0176596. [Google Scholar] [CrossRef] [PubMed]
- Peterson, K.J.; Dietrich, M.R.; McPeek, M.A. MicroRNAs and Metazoan Macroevolution: Insights into Canalization, Complexity, and the Cambrian Explosion. BioEssays 2009, 31, 736–747. [Google Scholar] [CrossRef] [PubMed]
- Christodoulou, F.; Raible, F.; Tomer, R.; Simakov, O.; Trachana, K.; Klaus, S.; Snyman, H.; Hannon, G.J.; Bork, P.; Arendt, D. Ancient Animal MicroRNAs and the Evolution of Tissue Identity. Nature 2010, 463, 1084–1088. [Google Scholar] [CrossRef] [PubMed]
- Farh, K.K.-H.; Grimson, A.; Jan, C.; Lewis, B.P.; Johnston, W.K.; Lim, L.P.; Burge, C.B.; Bartel, D.P. The Widespread Impact of Mammalian MicroRNAs on MRNA Repression and Evolution. Science 2005, 310, 1817–1821. [Google Scholar] [CrossRef] [PubMed]
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional Regulation of the Heterochronic Gene Lin-14 by Lin-4 Mediates Temporal Pattern Formation in C. Elegans. Cell 1993, 75, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Wienholds, E.; Plasterk, R.H.A. MicroRNA Function in Animal Development. FEBS Lett. 2005, 579, 5911–5922. [Google Scholar] [CrossRef]
- Hobert, O. Architecture of a MicroRNA-Controlled Gene Regulatory Network That Diversifies Neuronal Cell Fates. Cold Spring Harb. Symp. Quant. Biol. 2006, 71, 181–188. [Google Scholar] [CrossRef]
- Zhao, Y.; Ransom, J.F.; Li, A.; Vedantham, V.; von Drehle, M.; Muth, A.N.; Tsuchihashi, T.; McManus, M.T.; Schwartz, R.J.; Srivastava, D. Dysregulation of Cardiogenesis, Cardiac Conduction, and Cell Cycle in Mice Lacking MiRNA-1-2. Cell 2007, 129, 303–317. [Google Scholar] [CrossRef]
- Liu, N.; Bezprozvannaya, S.; Williams, A.H.; Qi, X.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. MicroRNA-133a Regulates Cardiomyocyte Proliferation and Suppresses Smooth Muscle Gene Expression in the Heart. Genes Dev. 2008, 22, 3242–3254. [Google Scholar] [CrossRef]
- Crist, C.G.; Montarras, D.; Pallafacchina, G.; Rocancourt, D.; Cumano, A.; Conway, S.J.; Buckingham, M. Muscle Stem Cell Behavior Is Modified by MicroRNA-27 Regulation of Pax3 Expression. Proc. Natl. Acad. Sci. USA 2009, 106, 13383–13387. [Google Scholar] [CrossRef]
- Belote, R.L.; Le, D.; Maynard, A.; Lang, U.E.; Sinclair, A.; Lohman, B.K.; Planells-Palop, V.; Baskin, L.; Tward, A.D.; Darmanis, S.; et al. Human Melanocyte Development and Melanoma Dedifferentiation at Single-Cell Resolution. Nat. Cell. Biol. 2021, 23, 1035–1047. [Google Scholar] [CrossRef] [PubMed]
- Tsoi, J.; Robert, L.; Paraiso, K.; Galvan, C.; Sheu, K.M.; Lay, J.; Wong, D.J.L.; Atefi, M.; Shirazi, R.; Wang, X.; et al. Multi-Stage Differentiation Defines Melanoma Subtypes with Differential Vulnerability to Drug-Induced Iron-Dependent Oxidative Stress. Cancer Cell 2018, 33, 890–904.e5. [Google Scholar] [CrossRef] [PubMed]
- Slack, F.J.; Chinnaiyan, A.M. The Role of Non-Coding RNAs in Oncology. Cell 2019, 179, 1033–1055. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Siverly, A.N.; Chen, D.; Wang, M.; Yuan, Y.; Wang, Y.; Lee, H.; Zhang, J.; Muller, W.J.; Liang, H.; et al. Ablation of MiR-10b Suppresses Oncogene-Induced Mammary Tumorigenesis and Metastasis and Reactivates Tumor-Suppressive Pathways. Cancer Res. 2016, 76, 6424–6435. [Google Scholar] [CrossRef]
- Hatley, M.E.; Patrick, D.M.; Garcia, M.R.; Richardson, J.A.; Bassel-Duby, R.; van Rooij, E.; Olson, E.N. Modulation of K-Ras-Dependent Lung Tumorigenesis by MicroRNA-21. Cancer Cell 2010, 18, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Edmonds, M.D.; Boyd, K.L.; Moyo, T.; Mitra, R.; Duszynski, R.; Arrate, M.P.; Chen, X.; Zhao, Z.; Blackwell, T.S.; Andl, T.; et al. MicroRNA-31 Initiates Lung Tumorigenesis and Promotes Mutant KRAS-Driven Lung Cancer. J. Clin. Investig. 2015, 126, 349–364. [Google Scholar] [CrossRef]
- Lv, C.; Li, F.; Li, X.; Tian, Y.; Zhang, Y.; Sheng, X.; Song, Y.; Meng, Q.; Yuan, S.; Luan, L.; et al. MiR-31 Promotes Mammary Stem Cell Expansion and Breast Tumorigenesis by Suppressing Wnt Signaling Antagonists. Nat. Commun. 2017, 8, 1036. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Lewis, B.P.; Shih, I.; Jones-Rhoades, M.W.; Bartel, D.P.; Burge, C.B. Prediction of Mammalian MicroRNA Targets. Cell 2003, 115, 787–798. [Google Scholar] [CrossRef]
- Friedman, R.C.; Farh, K.K.-H.; Burge, C.B.; Bartel, D.P. Most Mammalian MRNAs Are Conserved Targets of MicroRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef]
- Lim, L.P.; Lau, N.C.; Garrett-Engele, P.; Grimson, A.; Schelter, J.M.; Castle, J.; Bartel, D.P.; Linsley, P.S.; Johnson, J.M. Microarray Analysis Shows That Some MicroRNAs Downregulate Large Numbers of Target MRNAs. Nature 2005, 433, 769–773. [Google Scholar] [CrossRef] [PubMed]
- Stark, A.; Brennecke, J.; Bushati, N.; Russell, R.B.; Cohen, S.M. Animal MicroRNAs Confer Robustness to Gene Expression and Have a Significant Impact on 3′UTR Evolution. Cell 2005, 123, 1133–1146. [Google Scholar] [CrossRef] [PubMed]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved Seed Pairing, Often Flanked by Adenosines, Indicates That Thousands of Human Genes Are MicroRNA Targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Brennecke, J.; Stark, A.; Russell, R.B.; Cohen, S.M. Principles of MicroRNA–Target Recognition. PLoS Biol. 2005, 3, e85. [Google Scholar] [CrossRef] [PubMed]
- Krek, A.; Grün, D.; Poy, M.N.; Wolf, R.; Rosenberg, L.; Epstein, E.J.; MacMenamin, P.; da Piedade, I.; Gunsalus, K.C.; Stoffel, M.; et al. Combinatorial MicroRNA Target Predictions. Nat. Genet. 2005, 37, 495–500. [Google Scholar] [CrossRef]
- Wang, J.; Chen, J.; Sen, S. MicroRNA as Biomarkers and Diagnostics. J. Cell Physiol. 2016, 231, 25–30. [Google Scholar] [CrossRef]
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. MiRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef]
- Bonneau, E.; Neveu, B.; Kostantin, E.; Tsongalis, G.J.; De Guire, V. How Close Are MiRNAs from Clinical Practice? A Perspective on the Diagnostic and Therapeutic Market. Ejifcc 2019, 30, 114–127. [Google Scholar]
- Pogribny, I.P. MicroRNAs as Biomarkers for Clinical Studies. Exp. Biol. Med. 2018, 243, 283–290. [Google Scholar] [CrossRef]
- Saliminejad, K.; Khorram Khorshid, H.R.; Ghaffari, S.H. Why Have MicroRNA Biomarkers Not Been Translated from Bench to Clinic? Future Oncol. 2019, 15, 801–803. [Google Scholar] [CrossRef]
- Labourier, E.; Shifrin, A.; Busseniers, A.E.; Lupo, M.A.; Manganelli, M.L.; Andruss, B.; Wylie, D.; Beaudenon-Huibregtse, S. Molecular Testing for MiRNA, MRNA, and DNA on Fine-Needle Aspiration Improves the Preoperative Diagnosis of Thyroid Nodules With Indeterminate Cytology. J. Clin. Endocrinol. Metab. 2015, 100, 2743–2750. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Slack, F.J. MicroRNA Therapeutics: Towards a New Era for the Management of Cancer and Other Diseases. Nat. Rev. Drug. Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- van Zandwijk, N.; Pavlakis, N.; Kao, S.C.; Linton, A.; Boyer, M.J.; Clarke, S.; Huynh, Y.; Chrzanowska, A.; Fulham, M.J.; Bailey, D.L.; et al. Safety and Activity of MicroRNA-Loaded Minicells in Patients with Recurrent Malignant Pleural Mesothelioma: A First-in-Man, Phase 1, Open-Label, Dose-Escalation Study. Lancet Oncol. 2017, 18, 1386–1396. [Google Scholar] [CrossRef] [PubMed]
- Foss, F.M.; Querfeld, C.; Kim, Y.H.; Pinter-Brown, L.C.; William, B.M.; Porcu, P.; Pacheco, T.; Haverkos, B.; DeSimone, J.; Guitart, J.; et al. Ph 1 Study of MRG-106, an Inhibitor of MiR-155, in CTCL. JCO 2018, 36, 2511. [Google Scholar] [CrossRef]
- Zhang, S.; Cheng, Z.; Wang, Y.; Han, T. The Risks of MiRNA Therapeutics: In a Drug Target Perspective. Drug Des. Dev. Ther. 2021, 15, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Jarry, J.; Schadendorf, D.; Greenwood, C.; Spatz, A.; van Kempen, L.C. The Validity of Circulating MicroRNAs in Oncology: Five Years of Challenges and Contradictions. Mol. Oncol. 2014, 8, 819–829. [Google Scholar] [CrossRef]
- Torres, R.; Lang, U.E.; Hejna, M.; Shelton, S.J.; Joseph, N.M.; Shain, A.H.; Yeh, I.; Wei, M.L.; Oldham, M.C.; Bastian, B.C.; et al. MicroRNA Ratios Distinguish Melanomas from Nevi. J. Invest. Dermatol. 2020, 140, 164–173.e7. [Google Scholar] [CrossRef]
- Witwer, K.W.; Halushka, M.K. Toward the Promise of MicroRNAs—Enhancing Reproducibility and Rigor in MicroRNA Research. RNA Biol. 2016, 13, 1103–1116. [Google Scholar] [CrossRef]
- Wang, C.; Yang, X.; Chen, Y.; Chen, L. The Versatile Role of MicroRNA-30a in Human Cancer. Cell Physiol. Biochem. 2017, 41, 1616–1632. [Google Scholar] [CrossRef]
- Senfter, D.; Madlener, S.; Krupitza, G.; Mader, R.M. The MicroRNA-200 Family: Still Much to Discover. Biomol. Concepts 2016, 7, 311–319. [Google Scholar] [CrossRef]
- Rezaei, T.; Amini, M.; Hashemi, Z.S.; Mansoori, B.; Rezaei, S.; Karami, H.; Mosafer, J.; Mokhtarzadeh, A.; Baradaran, B. MicroRNA-181 Serves as a Dual-Role Regulator in the Development of Human Cancers. Free Radic. Biol. Med. 2020, 152, 432–454. [Google Scholar] [CrossRef] [PubMed]
- Banzhaf-Strathmann, J.; Edbauer, D. Good Guy or Bad Guy: The Opposing Roles of MicroRNA 125b in Cancer. Cell Commun. Signal. 2014, 12, 30. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Kunhiraman, H.; Perera, R.J. The Paradoxical Behavior of MicroRNA-211 in Melanomas and Other Human Cancers. Front. Oncol. 2021, 10, 628367. [Google Scholar] [CrossRef] [PubMed]
- Shirvani, H.; Ghanavi, J.; Aliabadi, A.; Mousavinasab, F.; Talebi, M.; Majidpoor, J.; Najafi, S.; Miryounesi, S.M.; Aghaei Zarch, S.M. MiR-211 Plays a Dual Role in Cancer Development: From Tumor Suppressor to Tumor Enhancer. Cell. Signal. 2023, 101, 110504. [Google Scholar] [CrossRef]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. OncomiR or Tumor Suppressor? The Duplicity of MicroRNAs in Cancer. Cancer Res. 2016, 76, 3666–3670. [Google Scholar] [CrossRef]
- Gustafson, D.; Tyryshkin, K.; Renwick, N. MicroRNA-Guided Diagnostics in Clinical Samples. Best Pract. Res. Clin. Endocrinol. Metab. 2016, 30, 563–575. [Google Scholar] [CrossRef]
- Faraldi, M.; Gomarasca, M.; Sansoni, V.; Perego, S.; Banfi, G.; Lombardi, G. Normalization Strategies Differently Affect Circulating MiRNA Profile Associated with the Training Status. Sci. Rep. 2019, 9, 1584. [Google Scholar] [CrossRef]
- Baek, D.; Villén, J.; Shin, C.; Camargo, F.D.; Gygi, S.P.; Bartel, D.P. The Impact of MicroRNAs on Protein Output. Nature 2008, 455, 64–71. [Google Scholar] [CrossRef]
- Ebert, M.S.; Sharp, P.A. Roles for MicroRNAs in Conferring Robustness to Biological Processes. Cell 2012, 149, 515–524. [Google Scholar] [CrossRef]
- Siciliano, V.; Garzilli, I.; Fracassi, C.; Criscuolo, S.; Ventre, S.; di Bernardo, D. MiRNAs Confer Phenotypic Robustness to Gene Networks by Suppressing Biological Noise. Nat. Commun. 2013, 4, 2364. [Google Scholar] [CrossRef]
- Mukherji, S.; Ebert, M.S.; Zheng, G.X.Y.; Tsang, J.S.; Sharp, P.A.; van Oudenaarden, A. MicroRNAs Can Generate Thresholds in Target Gene Expression. Nat. Genet. 2011, 43, 854–859. [Google Scholar] [CrossRef] [PubMed]
- Arvey, A.; Larsson, E.; Sander, C.; Leslie, C.S.; Marks, D.S. Target MRNA Abundance Dilutes MicroRNA and SiRNA Activity. Mol. Syst. Biol. 2010, 6, 363. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.M.; Baek, D.; Shin, C.; Bell, G.W.; Grimson, A.; Bartel, D.P. Weak Seed-Pairing Stability and High Target-Site Abundance Decrease the Proficiency of Lsy-6 and Other MicroRNAs. Nat. Struct. Mol. Biol. 2011, 18, 1139–1146. [Google Scholar] [CrossRef] [PubMed]
- Denzler, R.; McGeary, S.E.; Title, A.C.; Agarwal, V.; Bartel, D.P.; Stoffel, M. Impact of MicroRNA Levels, Target-Site Complementarity, and Cooperativity on Competing Endogenous RNA-Regulated Gene Expression. Mol. Cell 2016, 64, 565–579. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-T.; Risom, T.; Strauss, W.M. Evolutionary Conservation of MicroRNA Regulatory Circuits: An Examination of MicroRNA Gene Complexity and Conserved MicroRNA-Target Interactions through Metazoan Phylogeny. DNA Cell Biol. 2007, 26, 209–218. [Google Scholar] [CrossRef]
- Thomson, J.M.; Newman, M.; Parker, J.S.; Morin-Kensicki, E.M.; Wright, T.; Hammond, S.M. Extensive Post-Transcriptional Regulation of MicroRNAs and Its Implications for Cancer. Genes Dev. 2006, 20, 2202–2207. [Google Scholar] [CrossRef]
- Schmiedel, J.M.; Klemm, S.L.; Zheng, Y.; Sahay, A.; Blüthgen, N.; Marks, D.S.; van Oudenaarden, A. MicroRNA Control of Protein Expression Noise. Science 2015, 348, 128–132. [Google Scholar] [CrossRef]
- Li, C.; Liau, E.S.; Lee, Y.; Huang, Y.; Liu, Z.; Willems, A.; Garside, V.; McGlinn, E.; Chen, J.; Hong, T. MicroRNA Governs Bistable Cell Differentiation and Lineage Segregation via a Noncanonical Feedback. Mol. Syst. Biol. 2021, 17, 20209945. [Google Scholar] [CrossRef]
- Choi, W.-Y.; Giraldez, A.J.; Schier, A.F. Target Protectors Reveal Dampening and Balancing of Nodal Agonist and Antagonist by MiR-430. Science 2007, 318, 271–274. [Google Scholar] [CrossRef]
- Li, X.; Carthew, R.W. A MicroRNA Mediates EGF Receptor Signaling and Promotes Photoreceptor Differentiation in the Drosophila Eye. Cell 2005, 123, 1267–1277. [Google Scholar] [CrossRef]
- Li, X.; Cassidy, J.J.; Reinke, C.A.; Fischboeck, S.; Carthew, R.W. A MicroRNA Imparts Robustness against Environmental Fluctuation during Development. Cell 2009, 137, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Herranz, H.; Cohen, S.M. MicroRNAs and Gene Regulatory Networks: Managing the Impact of Noise in Biological Systems. Genes Dev. 2010, 24, 1339–1344. [Google Scholar] [CrossRef] [PubMed]
- Van Rooij, E.; Sutherland, L.B.; Qi, X.; Richardson, J.A.; Hill, J.; Olson, E.N. Control of Stress-Dependent Cardiac Growth and Gene Expression by a MicroRNA. Science 2007, 316, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.X.Y.; Ravi, A.; Calabrese, J.M.; Medeiros, L.A.; Kirak, O.; Dennis, L.M.; Jaenisch, R.; Burge, C.B.; Sharp, P.A. A Latent Pro-Survival Function for the Mir-290-295 Cluster in Mouse Embryonic Stem Cells. PLoS Genet. 2011, 7, e1002054. [Google Scholar] [CrossRef] [PubMed]
- Brenner, J.L.; Jasiewicz, K.L.; Fahley, A.F.; Kemp, B.J.; Abbott, A.L. Loss of Individual MicroRNAs Causes Mutant Phenotypes in Sensitized Genetic Backgrounds in C. Elegans. Curr. Biol. 2010, 20, 1321–1325. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA Expression Profiles Classify Human Cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef]
- Xu, X.; Wang, K.; Vera, O.; Verma, A.; Jasani, N.; Bok, I.; Elemento, O.; Du, D.; Yu, X.; Karreth, F.A. Gain of Chromosome 1q Perturbs a Competitive Endogenous RNA Network to Promote Melanoma Metastasis. Cancer Res. 2022, 82, 3016–3031. [Google Scholar] [CrossRef]
- Bastian, B.C.; LeBoit, P.E.; Hamm, H.; Bröcker, E.B.; Pinkel, D. Chromosomal Gains and Losses in Primary Cutaneous Melanomas Detected by Comparative Genomic Hybridization. Cancer Res. 1998, 58, 2170–2175. [Google Scholar]
- Elder, D.E.; Bastian, B.C.; Cree, I.A.; Massi, D.; Scolyer, R.A. The 2018 World Health Organization Classification of Cutaneous, Mucosal, and Uveal Melanoma: Detailed Analysis of 9 Distinct Subtypes Defined by Their Evolutionary Pathway. Arch. Pathol. Lab. Med. 2020, 144, 500–522. [Google Scholar] [CrossRef]
- Bastian, B.C. The Molecular Pathology of Melanoma: An Integrated Taxonomy of Melanocytic Neoplasia. Annu. Rev. Pathol. Mech. Dis. 2014, 9, 239–271. [Google Scholar] [CrossRef]
- Jentsch, M.C.; Burger, H.; Meddens, M.B.M.; Beijers, L.; van den Heuvel, E.R.; Meddens, M.J.M.; Schoevers, R.A. Gender Differences in Developing Biomarker-Based Major Depressive Disorder Diagnostics. Int. J. Mol. Sci. 2020, 21, 3039. [Google Scholar] [CrossRef]
- McNeal, A.S.; Belote, R.L.; Zeng, H.; Urquijo, M.; Barker, K.; Torres, R.; Curtin, M.; Shain, A.H.; Andtbacka, R.H.; Holmen, S.; et al. BRAFV600E Induces Reversible Mitotic Arrest in Human Melanocytes via Microrna-Mediated Suppression of AURKB. Elife 2021, 10, e70385. [Google Scholar] [CrossRef] [PubMed]
- Elmore, J.G.; Barnhill, R.L.; Elder, D.E.; Longton, G.M.; Pepe, M.S.; Reisch, L.M.; Carney, P.A.; Titus, L.J.; Nelson, H.D.; Onega, T.; et al. Pathologists’ Diagnosis of Invasive Melanoma and Melanocytic Proliferations: Observer Accuracy and Reproducibility Study. BMJ 2017, 357, j2813. [Google Scholar] [CrossRef] [PubMed]
- Deacon, D.C.; Smith, E.A.; Judson-Torres, R.L. Molecular Biomarkers for Melanoma Screening, Diagnosis and Prognosis: Current State and Future Prospects. Front. Med. 2021, 8, 642380. [Google Scholar] [CrossRef] [PubMed]
- Ronchi, A.; Pagliuca, F.; Zito Marino, F.; Argenziano, G.; Brancaccio, G.; Alfano, R.; Signoriello, G.; Moscarella, E.; Franco, R. Second Diagnostic Opinion by Experienced Dermatopathologists in the Setting of a Referral Regional Melanoma Unit Significantly Improves the Clinical Management of Patients With Cutaneous Melanoma. Front. Med. 2021, 7, 568946. [Google Scholar] [CrossRef] [PubMed]
- Shain, A.H.; Yeh, I.; Kovalyshyn, I.; Sriharan, A.; Talevich, E.; Gagnon, A.; Dummer, R.; North, J.; Pincus, L.; Ruben, B.; et al. The Genetic Evolution of Melanoma from Precursor Lesions. N. Engl. J. Med. 2015, 373, 1926–1936. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Feilotter, H.E.; Paré, G.C.; Zhang, X.; Pemberton, J.G.W.; Garady, C.; Lai, D.; Yang, X.; Tron, V.A. MicroRNA-193b Represses Cell Proliferation and Regulates Cyclin D1 in Melanoma. Am. J. Pathol. 2010, 176, 2520–2529. [Google Scholar] [CrossRef]
- Gao, Y.; Xu, J.; Li, H.; Hu, Y.; Yu, G. Identification of Metastasis-Associated MicroRNAs in Metastatic Melanoma by MiRNA Expression Profile and Experimental Validation. Front. Genet. 2021, 12, 663110. [Google Scholar] [CrossRef]
- Komina, A.; Palkina, N.; Aksenenko, M.; Tsyrenzhapova, S.; Ruksha, T. Antiproliferative and Pro-Apoptotic Effects of MiR-4286 Inhibition in Melanoma Cells. PLoS ONE 2016, 11, e0168229. [Google Scholar] [CrossRef] [PubMed]
- Kozubek, J.; Ma, Z.; Fleming, E.; Duggan, T.; Wu, R.; Shin, D.-G.; Dadras, S.S. In-Depth Characterization of MicroRNA Transcriptome in Melanoma. PLoS ONE 2013, 8, e72699. [Google Scholar] [CrossRef]
- Kutlay, A.; Aydin Son, Y. Integrative Predictive Modeling of Metastasis in Melanoma Cancer Based on MicroRNA, MRNA, and DNA Methylation Data. Front. Mol. Biosci. 2021, 8, 637355. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Tetzlaff, M.T.; Liu, A.; Liegl-Atzwanger, B.; Guo, J.; Xu, X. Loss of MicroRNA-205 Expression Is Associated with Melanoma Progression. Lab. Invest. 2012, 92, 1084–1096. [Google Scholar] [CrossRef] [PubMed]
- Lohcharoenkal, W.; Das Mahapatra, K.; Pasquali, L.; Crudden, C.; Kular, L.; Akkaya Ulum, Y.Z.; Zhang, L.; Xu Landén, N.; Girnita, L.; Jagodic, M.; et al. Genome-Wide Screen for MicroRNAs Reveals a Role for MiR-203 in Melanoma Metastasis. J. Investig. Dermatol. 2018, 138, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Chen, S.; Qu, L.; Wang, Y.; Chen, H.; He, C. Identification of a Five-MiRNA Signature Predicting Survival in Cutaneous Melanoma Cancer Patients. PeerJ 2019, 7, e7831. [Google Scholar] [CrossRef]
- Saldanha, G.; Elshaw, S.; Sachs, P.; Alharbi, H.; Shah, P.; Jothi, A.; Pringle, J.H. MicroRNA-10b Is a Prognostic Biomarker for Melanoma. Mod. Pathol. 2016, 29, 112–121. [Google Scholar] [CrossRef]
- Sand, M.; Skrygan, M.; Sand, D.; Georgas, D.; Gambichler, T.; Hahn, S.A.; Altmeyer, P.; Bechara, F.G. Comparative Microarray Analysis of MicroRNA Expression Profiles in Primary Cutaneous Malignant Melanoma, Cutaneous Malignant Melanoma Metastases, and Benign Melanocytic Nevi. Cell Tissue Res. 2013, 351, 85–98. [Google Scholar] [CrossRef]
- Schultz, J.; Lorenz, P.; Gross, G.; Ibrahim, S.; Kunz, M. MicroRNA Let-7b Targets Important Cell Cycle Molecules in Malignant Melanoma Cells and Interferes with Anchorage-Independent Growth. Cell Res. 2008, 18, 549–557. [Google Scholar] [CrossRef]
- Segura, M.F.; Belitskaya-Lévy, I.; Rose, A.E.; Zakrzewski, J.; Gaziel, A.; Hanniford, D.; Darvishian, F.; Berman, R.S.; Shapiro, R.L.; Pavlick, A.C.; et al. Melanoma MicroRNA Signature Predicts Post-Recurrence Survival. Clin. Cancer Res. 2010, 16, 1577–1586. [Google Scholar] [CrossRef]
- Singh, N.; Eberhardt, M.; Wolkenhauer, O.; Vera, J.; Gupta, S.K. An Integrative Network-Driven Pipeline for Systematic Identification of LncRNA-Associated Regulatory Network Motifs in Metastatic Melanoma. BMC Bioinform. 2020, 21, 329. [Google Scholar] [CrossRef]
- Wandler, A.; Riber-Hansen, R.; Hager, H.; Hamilton-Dutoit, S.J.; Schmidt, H.; Nielsen, B.S.; Stougaard, M.; Steiniche, T. Quantification of MicroRNA-21 and MicroRNA-125b in Melanoma Tissue. Melanoma Res. 2017, 27, 417–428. [Google Scholar] [CrossRef]
- Wang, J.; Tao, Y.; Bian, Q. MiRNA and MRNA Expression Profiling Reveals Potential Biomarkers for Metastatic Cutaneous Melanoma. Expert Rev. Anticancer Ther. 2021, 21, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Watt, K.; Tyryshkin, K.; Renwick, N.; Craig, A.W.B. Distinguishing Tumor and Stromal Sources of MicroRNAs Linked to Metastasis in Cutaneous Melanoma. Transl. Oncol. 2020, 13, 100802. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wang, Y.; Chen, C.; Xue, Y.; Zheng, S.; Cai, L. Hsa-Let-7c-5p, Hsa-MiR-130b-3p, and Hsa-MiR-142-3p as Novel MiRNA Biomarkers for Melanoma Progression. Genet. Res. 2022, 2022, 5671562. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Brenn, T.; Brown, E.R.S.; Doherty, V.; Melton, D.W. Differential Expression of MicroRNAs during Melanoma Progression: MiR-200c, MiR-205 and MiR-211 Are Downregulated in Melanoma and Act as Tumour Suppressors. Br. J. Cancer 2012, 106, 553–561. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, L.; Jiang, L.; Zhang, X. Novel MicroRNA Biomarkers, MiR-142-5p, MiR-550a, MiR-1826, and MiR-1201, Were Identified for Primary Melanoma. J. Comput. Biol. 2020, 27, 815–824. [Google Scholar] [CrossRef]
- Zhao, C.-C.; Guo, H.; Wang, Y.; Li, J.-H. Comprehensive Upstream and Downstream Regulatory Analyses Identify MiR-675-3p as a Potential Prognostic Biomarker in Melanoma. Hum. Cell 2021, 34, 654–666. [Google Scholar] [CrossRef]
- Griffiths-Jones, S. The MicroRNA Registry. Nucleic Acids Res. 2004, 32, 109D–111D. [Google Scholar] [CrossRef]
- Vitiello, M.; Tuccoli, A.; D’Aurizio, R.; Sarti, S.; Giannecchini, L.; Lubrano, S.; Marranci, A.; Evangelista, M.; Peppicelli, S.; Ippolito, C.; et al. Context-Dependent MiR-204 and MiR-211 Affect the Biological Properties of Amelanotic and Melanotic Melanoma Cells. Oncotarget 2017, 8, 25395–25417. [Google Scholar] [CrossRef]
- Díaz-Martínez, M.; Benito-Jardón, L.; Alonso, L.; Koetz-Ploch, L.; Hernando, E.; Teixidó, J. MiR-204-5p and MiR-211-5p Contribute to BRAF Inhibitor Resistance in Melanoma. Cancer Res. 2018, 78, 1017–1030. [Google Scholar] [CrossRef]
- Shain, A.H.; Joseph, N.M.; Yu, R.; Benhamida, J.; Liu, S.; Prow, T.; Ruben, B.; North, J.; Pincus, L.; Yeh, I.; et al. Genomic and Transcriptomic Analysis Reveals Incremental Disruption of Key Signaling Pathways during Melanoma Evolution. Cancer Cell 2018, 34, 45–55.e4. [Google Scholar] [CrossRef]
- Babapoor, S.; Horwich, M.; Wu, R.; Levinson, S.; Gandhi, M.; Makkar, H.; Kristjansson, A.; Chang, M.; Dadras, S.S. MicroRNA in Situ Hybridization for MiR-211 Detection as an Ancillary Test in Melanoma Diagnosis. Mod. Pathol. 2016, 29, 461–475. [Google Scholar] [CrossRef] [PubMed]
- Boyle, G.M.; Woods, S.L.; Bonazzi, V.F.; Stark, M.S.; Hacker, E.; Aoude, L.G.; Dutton-Regester, K.; Cook, A.L.; Sturm, R.A.; Hayward, N.K. Melanoma Cell Invasiveness Is Regulated by MiR-211 Suppression of the BRN2 Transcription Factor: MiR-211 Regulates Melanoma Invasiveness via BRN2. Pigment Cell Melanoma Res. 2011, 24, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.T.; Fisher, D.E. MITF and UV Responses in Skin: From Pigmentation to Addiction. Pigment Cell Melanoma Res. 2019, 32, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Fane, M.E.; Chhabra, Y.; Smith, A.G.; Sturm, R.A. BRN 2, a POU Erful Driver of Melanoma Phenotype Switching and Metastasis. Pigment Cell Melanoma Res. 2019, 32, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Infarinato, N.R.; Stewart, K.S.; Yang, Y.; Gomez, N.C.; Pasolli, H.A.; Hidalgo, L.; Polak, L.; Carroll, T.S.; Fuchs, E. BMP Signaling: At the Gate between Activated Melanocyte Stem Cells and Differentiation. Genes Dev. 2020, 34, 1713–1734. [Google Scholar] [CrossRef]
- Carney, B.C.; Travis, T.E.; Moffatt, L.T.; Johnson, L.S.; McLawhorn, M.M.; Simbulan-Rosenthal, C.M.; Rosenthal, D.S.; Shupp, J.W. Hypopigmented Burn Hypertrophic Scar Contains Melanocytes That Can Be Signaled to Re-Pigment by Synthetic Alpha-Melanocyte Stimulating Hormone in Vitro. PLoS ONE 2021, 16, e0248985. [Google Scholar] [CrossRef]
- Michalak-Mićka, K.; Büchler, V.L.; Zapiórkowska-Blumer, N.; Biedermann, T.; Klar, A.S. Characterization of a Melanocyte Progenitor Population in Human Interfollicular Epidermis. Cell Rep. 2022, 38, 110419. [Google Scholar] [CrossRef]
- Mazar, J.; DeYoung, K.; Khaitan, D.; Meister, E.; Almodovar, A.; Goydos, J.; Ray, A.; Perera, R.J. The Regulation of MiRNA-211 Expression and Its Role in Melanoma Cell Invasiveness. PLoS ONE 2010, 5, e13779. [Google Scholar] [CrossRef]
- Mazar, J.; Qi, F.; Lee, B.; Marchica, J.; Govindarajan, S.; Shelley, J.; Li, J.-L.; Ray, A.; Perera, R.J. MicroRNA 211 Functions as a Metabolic Switch in Human Melanoma Cells. Mol Cell Biol. 2016, 36, 1090–1108. [Google Scholar] [CrossRef]
- Bell, R.E.; Khaled, M.; Netanely, D.; Schubert, S.; Golan, T.; Buxbaum, A.; Janas, M.M.; Postolsky, B.; Goldberg, M.S.; Shamir, R.; et al. Transcription Factor/MicroRNA Axis Blocks Melanoma Invasion Program by MiR-211 Targeting NUAK1. J. Investig. Dermatol. 2014, 134, 441–451. [Google Scholar] [CrossRef]
- De Luca, T.; Pelosi, A.; Trisciuoglio, D.; D’Aguanno, S.; Desideri, M.; Farini, V.; Di Martile, M.; Bellei, B.; Tupone, M.G.; Candiloro, A.; et al. MiR-211 and MITF Modulation by Bcl-2 Protein in Melanoma Cells: MiR-211 AND MITF MODULATION BY Bcl-2 PROTEIN. Mol. Carcinog. 2016, 55, 2304–2312. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Sahoo, A.; Sawada, J.; Marchica, J.; Sahoo, S.; Layng, F.I.A.L.; Finlay, D.; Mazar, J.; Joshi, P.; Komatsu, M.; et al. MicroRNA-211 Modulates the DUSP6-ERK5 Signaling Axis to Promote BRAFV600E-Driven Melanoma Growth In Vivo and BRAF/MEK Inhibitor Resistance. J. Investig. Dermatol. 2021, 141, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, A.; Sahoo, S.K.; Joshi, P.; Lee, B.; Perera, R.J. MicroRNA-211 Loss Promotes Metabolic Vulnerability and BRAF Inhibitor Sensitivity in Melanoma. J. Investig. Dermatol. 2019, 139, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Chen, Y.; Chen, H.; Zhao, Q.; Sun, Z.; Liu, D.; Li, X.; Zhang, Y.; Wang, J.; Xing, H.R. Exosomal MiR-211-5p Regulates Glucose Metabolism, Pyroptosis, and Immune Microenvironment of Melanoma through GNA15. Pharmacol. Res. 2023, 188, 106660. [Google Scholar] [CrossRef]
- Li, N.; Liu, Y.; Pang, H.; Lee, D.; Zhou, Y.; Xiao, Z. Methylation-Mediated Silencing of MicroRNA-211 Decreases the Sensitivity of Melanoma Cells to Cisplatin. Med. Sci. Monit. 2019, 25, 1590–1599. [Google Scholar] [CrossRef] [PubMed]
- Dror, S.; Sander, L.; Schwartz, H.; Sheinboim, D.; Barzilai, A.; Dishon, Y.; Apcher, S.; Golan, T.; Greenberger, S.; Barshack, I.; et al. Melanoma MiRNA Trafficking Controls Tumour Primary Niche Formation. Nat. Cell. Biol. 2016, 18, 1006–1017. [Google Scholar] [CrossRef]
- Sakurai, E.; Maesawa, C.; Shibazaki, M.; Yasuhira, S.; Oikawa, H.; Sato, M.; Tsunoda, K.; Ishikawa, Y.; Watanabe, A.; Takahashi, K.; et al. Downregulation of MicroRNA-211 Is Involved in Expression of Preferentially Expressed Antigen of Melanoma in Melanoma Cells. Int. J. Oncol. 2011, 39, 665–672. [Google Scholar] [CrossRef]
- Margue, C.; Philippidou, D.; Reinsbach, S.E.; Schmitt, M.; Behrmann, I.; Kreis, S. New Target Genes of MITF-Induced MicroRNA-211 Contribute to Melanoma Cell Invasion. PLoS ONE 2013, 8, e73473. [Google Scholar] [CrossRef]
- Levy, C.; Khaled, M.; Iliopoulos, D.; Janas, M.M.; Schubert, S.; Pinner, S.; Chen, P.-H.; Li, S.; Fletcher, A.L.; Yokoyama, S.; et al. Intronic MiR-211 Assumes the Tumor Suppressive Function of Its Host Gene in Melanoma. Mol. Cell 2010, 40, 841–849. [Google Scholar] [CrossRef]
- Linck, L.; Liebig, J.; Völler, D.; Eichner, N.; Lehmann, G.; Meister, G.; Bosserhoff, A. MicroRNA-Sequencing Data Analyzing Melanoma Development and Progression. Exp. Mol. Pathol. 2018, 105, 371–379. [Google Scholar] [CrossRef]
- Krepler, C.; Sproesser, K.; Brafford, P.; Beqiri, M.; Garman, B.; Xiao, M.; Shannan, B.; Watters, A.; Perego, M.; Zhang, G.; et al. A Comprehensive Patient-Derived Xenograft Collection Representing the Heterogeneity of Melanoma. Cell Rep. 2017, 21, 1953–1967. [Google Scholar] [CrossRef] [PubMed]
- Ben-David, U.; Siranosian, B.; Ha, G.; Tang, H.; Oren, Y.; Hinohara, K.; Strathdee, C.A.; Dempster, J.; Lyons, N.J.; Burns, R.; et al. Genetic and Transcriptional Evolution Alters Cancer Cell Line Drug Response. Nature 2018, 560, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Najem, A.; Wouters, J.; Krayem, M.; Rambow, F.; Sabbah, M.; Sales, F.; Awada, A.; Aerts, S.; Journe, F.; Marine, J.-C.; et al. Tyrosine-Dependent Phenotype Switching Occurs Early in Many Primary Melanoma Cultures Limiting Their Translational Value. Front. Oncol. 2021, 11, 780654. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Rao, C.; Li, H.; Chen, Y.; Fan, L.; Geng, H.; Li, S.; Qu, J.; Hou, L. Regulation of Pigmentation by MicroRNAs: MITF-Dependent MicroRNA-211 Targets TGF- β Receptor 2. Pigment Cell Melanoma Res. 2015, 28, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Cook, A.L.; Donatien, P.D.; Smith, A.G.; Murphy, M.; Jones, M.K.; Herlyn, M.; Bennett, D.C.; Helen Leonard, J.; Sturm, R.A. Human Melanoblasts in Culture: Expression of BRN2 and Synergistic Regulation by Fibroblast Growth Factor-2, Stem Cell Factor, and Endothelin-3. J. Investig. Dermatol. 2003, 121, 1150–1159. [Google Scholar] [CrossRef]
- Tirosh, I.; Izar, B.; Prakadan, S.M.; Wadsworth, M.H.; Treacy, D.; Trombetta, J.J.; Rotem, A.; Rodman, C.; Lian, C.; Murphy, G.; et al. Dissecting the Multicellular Ecosystem of Metastatic Melanoma by Single-Cell RNA-Seq. Science 2016, 352, 189–196. [Google Scholar] [CrossRef]
- Rabbie, R.; Ferguson, P.; Molina-Aguilar, C.; Adams, D.J.; Robles-Espinoza, C.D. Melanoma Subtypes: Genomic Profiles, Prognostic Molecular Markers and Therapeutic Possibilities. J. Pathol. 2019, 247, 539–551. [Google Scholar] [CrossRef]
- Michaloglou, C.; Vredeveld, L.C.W.; Soengas, M.S.; Denoyelle, C.; Kuilman, T.; van der Horst, C.M.A.M.; Majoor, D.M.; Shay, J.W.; Mooi, W.J.; Peeper, D.S. BRAFE600-Associated Senescence-like Cell Cycle Arrest of Human Naevi. Nature 2005, 436, 720–724. [Google Scholar] [CrossRef]
- Garnett, M.J.; Marais, R. Guilty as Charged: B-RAF Is a Human Oncogene. Cancer Cell 2004, 6, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Golan, T.; Parikh, R.; Jacob, E.; Vaknine, H.; Zemser-Werner, V.; Hershkovitz, D.; Malcov, H.; Leibou, S.; Reichman, H.; Sheinboim, D.; et al. Adipocytes Sensitize Melanoma Cells to Environmental TGF-β Cues by Repressing the Expression of MiR-211. Sci. Signal. 2019, 12, eaav6847. [Google Scholar] [CrossRef]
- Hanniford, D.; Zhong, J.; Koetz, L.; Gaziel-Sovran, A.; Lackaye, D.J.; Shang, S.; Pavlick, A.; Shapiro, R.; Berman, R.; Darvishian, F.; et al. A MiRNA-Based Signature Detected in Primary Melanoma Tissue Predicts Development of Brain Metastasis. Clin. Cancer Res. 2015, 21, 4903–4912. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
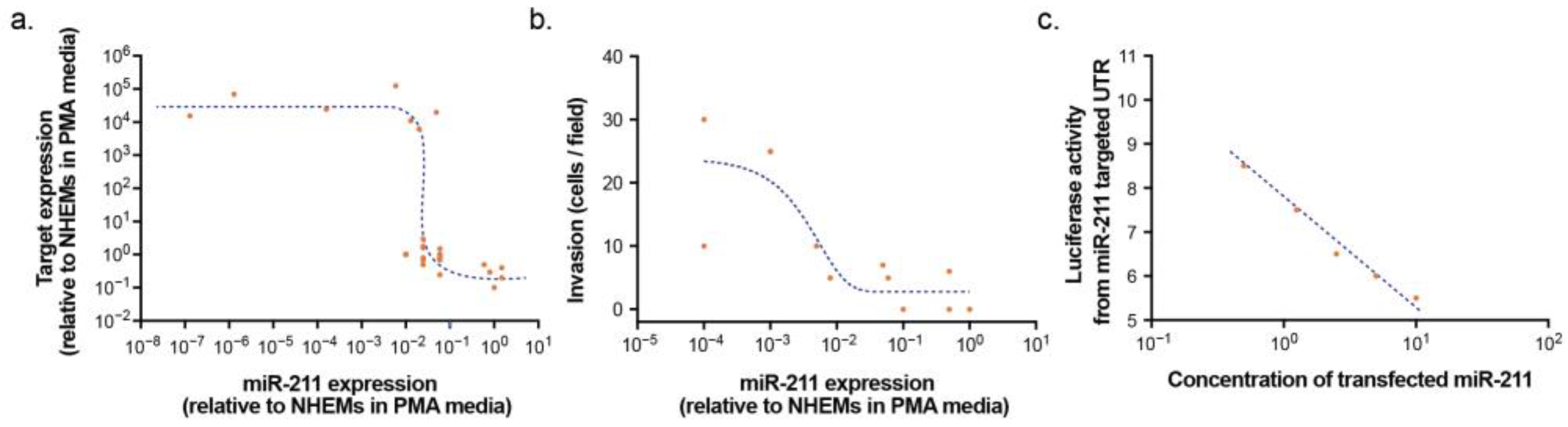{kind=link}
| miRNA | Nevus to Primary Melanoma | Primary Melanoma to Melanoma Metastasis | Nevus to Melanoma Metastasis |
|---|---|---|---|
| miR-23B | 6 | ||
| miR-125B | 6 | ||
| miR-211 | 5 | 3 | |
| miR-204 | 5 | 3 | |
| miR-125A | 4 | ||
| miR-455 | 4 | ||
| LET7-A | 4 | ||
| LET7-B | 3 | ||
| miR-100 | 3 | ||
| miR-183 | 3 | ||
| miR-149 | 3 | ||
| miR-99A | 3 | ||
| miR-26B | 3 | ||
| miR-155 | -4 | ||
| miR-21 | -5 | -3 | |
| miR-205 | 5 | 3 | |
| miR-141 | 4 | 5 | |
| miR-200A | 4 | 4 | |
| miR-203 | 4 | 4 | |
| miR-200B | 3 | 5 | 3 |
| miR-200C | 4 | ||
| miR-142 | -4 | -3 | -3 |
| miR-224 | 4 | ||
| miR-203A | 4 | ||
| miR-29C | -3 | ||
| miR-218-2 | -3 | ||
| miR-326 | -3 | ||
| miR-4491 | -3 | ||
| miR-625 | -3 | ||
| miR-675 | -3 | ||
| miR-766 | -3 | ||
| miR-215 | -4 | ||
| miR-3130 | -4 | ||
| MN-specific | 5 | 5 | |
| PM-specific | -5 | 5 | |
| MM-specific | -5 | -5 |
| Family | Nevus to Primary Melanoma | CV | Nevus to Melanoma Metastasis | CV | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MIR204/211 | 1 | 1 | 0.5 | 0.5 | 1 | 0 | 0.5 | 0.5 | 0 | 0.7 | 1 | 0.5 | 0.5 | 0 | 1 | 1.04 |
| MIR23 | 0.5 | 0.5 | 0 | 0.5 | 1 | 0 | 0.5 | 0 | 0.5 | 0.86 | 0.5 | 0 | 0 | 0 | 0 | 2.24 |
| MIR141/200 | 0 | 0 | 0.8 | 0 | 0.4 | 0.6 | 0.4 | 0 | 0.4 | 1.04 | 0.6 | 0.8 | 0.6 | 0 | 0 | 1.2 |
| MIR203 | 0 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0 | 0 | 0 | 0.95 | 0.5 | 0.5 | 0 | 0 | 0 | 1.37 |
| MIR10/100 | 0.38 | 0 | 0.38 | 0.25 | 0.5 | 0.13 | 0.13 | 0 | 0.25 | 0.78 | 0.25 | 0.38 | 0 | -0.13 | 0 | 1.68 |
| [47] | [90] | [92] | [105] | [100] | [104] | [96] | [89] | [97] | [87] | [92] | [96] | [98] | [94] | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vand-Rajabpour, F.; Savage, M.; Belote, R.L.; Judson-Torres, R.L. Critical Considerations for Investigating MicroRNAs during Tumorigenesis: A Case Study in Conceptual and Contextual Nuances of miR-211-5p in Melanoma. Epigenomes 2023, 7, 9. https://doi.org/10.3390/epigenomes7020009
Vand-Rajabpour F, Savage M, Belote RL, Judson-Torres RL. Critical Considerations for Investigating MicroRNAs during Tumorigenesis: A Case Study in Conceptual and Contextual Nuances of miR-211-5p in Melanoma. Epigenomes. 2023; 7(2):9. https://doi.org/10.3390/epigenomes7020009
Chicago/Turabian StyleVand-Rajabpour, Fatemeh, Meghan Savage, Rachel L. Belote, and Robert L. Judson-Torres. 2023. "Critical Considerations for Investigating MicroRNAs during Tumorigenesis: A Case Study in Conceptual and Contextual Nuances of miR-211-5p in Melanoma" Epigenomes 7, no. 2: 9. https://doi.org/10.3390/epigenomes7020009
APA StyleVand-Rajabpour, F., Savage, M., Belote, R. L., & Judson-Torres, R. L. (2023). Critical Considerations for Investigating MicroRNAs during Tumorigenesis: A Case Study in Conceptual and Contextual Nuances of miR-211-5p in Melanoma. Epigenomes, 7(2), 9. https://doi.org/10.3390/epigenomes7020009