
Alpha, Beta and Gamma Taxonomy of Biocontrol Agent Diaeretiella rapae (Hymenoptera, Braconidae)



Simple Summary
Abstract
1. Introduction

2. Materials and Methods
2.1. Sample Collection and Morphological Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aphidius absinthii Marshall, 1896 | Aphidius matricariae Haliday, 1834 |
| Aphidius aquilus Mackauer, 1961 | Aphidius microlophii Pennachio & Tremblay, 1987 |
| Aphidius areolatus Ashmead, 1906 | Aphidius phalangomyzi Starý, 1963 |
| Aphidius arvensis (Starý, 1960) | Aphidius platensis Brethes, 1913 |
| Aphidius asiaticus Kim & Tomanović, 2021 | Aphidius plocomaphidis (Starý, 1973) |
| Aphidius avenae Haliday, 1834 | Aphidius rhopalosiphi de Stefani-Perez, 1902 |
| Aphidius avenaphis (Fitchk, 1861) | Aphidius ribis Haliday, 1834 |
| Aphidius balcanicus Tomanović & Petrović, 2011 | Aphidius rosae Haliday, 1834 |
| Aphidius banksae Kittel, 2016 | Aphidius salicis Haliday, 1834 |
| Aphidius cingulatus Ruthe, 1859 | Aphidius schimitscheki (Starý, 1960) |
| Aphidius colemani Viereck, 1912 | Aphidius setiger (Mackauer, 1961) |
| Aphidius eadyi Starý, Gonzáles & Hall, 1980 | Aphidius smithi Sharma & Subba Rao, 1959 |
| Aphidius ericaphidis Pike & Starý, 2011 | Aphidius sonchi Marshall, 1896 |
| Aphidius ervi Haliday, 1834 | Aphidius staticobii Tomanović & Petrović, 2012 |
| Aphidius funebris Mackauer, 1961 | Aphidius sussi Pennachio& Tremblay, 1989 |
| Aphidius gerani Tomanović & Kavallieratos, 2009 | Aphidius tanacetarius Mackauer, 1962 |
| Aphidius gifuensis Ashmead, 1906 | Aphidius tarsalis van Achterberg, 2006 |
| Aphidius hieraciorum Starý, 1962 | Aphidius transcaspicus Telenga, 1958 |
| Aphidius hortensis Marshall, 1896 | Aphidius urticae Haliday, 1834 |
| Aphidius leclanti Tomanović & Chaubet, 2013 | Aphidius uzbekistanicus Luzhetzki, 1960 |
| Aphidius longipetiolus Takada, 1968 | Aphidius viaticus (Sedlag, 1968) |
| Aphidius longistigmus Kim & Tomanović, 2021 |

2.2. Molecular Analysis
3. Results
3.1. Morphology
| D. rapae | Aphidius (Minimal Value) | Aphidius (Maximal Value) | |||
|---|---|---|---|---|---|
| Antennae | (12) 13–14 | 12–13 | (A. salicis) | (19) 20–21 | (A. eadyi) |
| F1 l/w | 2.5–3.5 | 2.11–2.52 | (A. longistigmus) | 4–5 | (A. gifuensis) |
| F2 l/w | 2–3 | 1.63–1.76 | (A. areolatus) | 3–4,3 | (A. banksae) |
| MPS F1 | 0–1 | 0 | (A. ribis) | 3–6 | (A. rosae) |
| MPS F2 | 2–4 | 0 | (A. leclanti) | 4–7 | (A. cingulatus) |
| F1/F2 | 1 | 0.85–0.93 | (A. banksae) | 1.06 | (A. leclanti) |
| MP | 3–4 | 2 | (A. arvensis) | 4 | (A. absinthii) |
| LP | 2 | 1 | (A. arvensis) | 3 | (A. uzbekistanicus) |
| Ti | 0.29–0.36 | 0.3–0.4 | (A. schimitscheki) | 0.74–0.8 | (A. cingulatus) |
| Pt l/w | 2.9–4 | 2.8–3 | (A. plocomaphidis) | 4.96–5.46 | (A. longistigmus) |
| Ptl/Mtl | 1.39–2.63 | 0.93–1.05 | (A. urticae) | 1.5–2 | (A. avenae) |
| Pet l/w | 1.93–2.65 | 2.17 | (A. areolatus) | 3.88–4 | (A. geranii) |




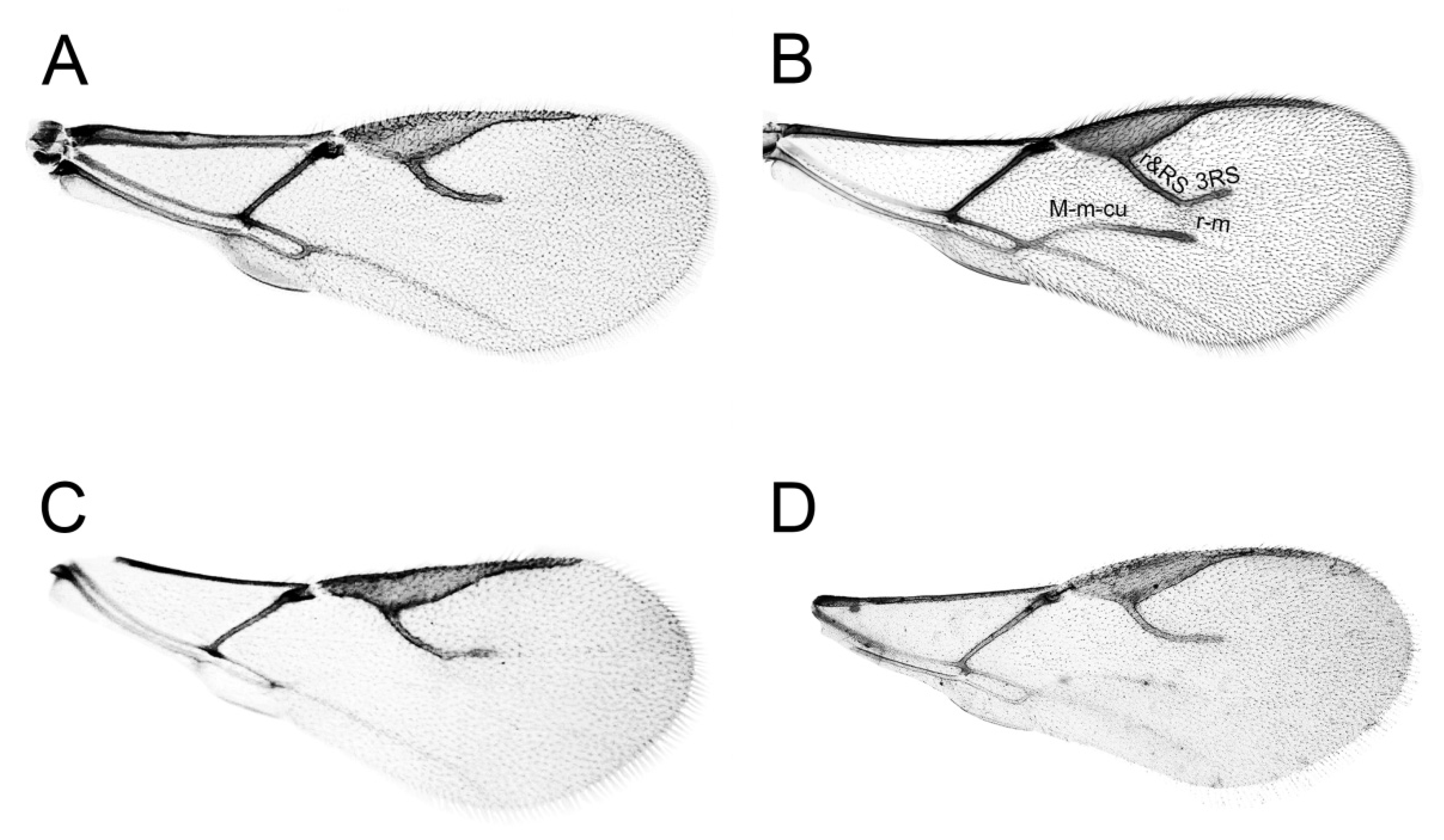
3.2. Molecular Analysis
3.3. Re-Description of Aphidius Rapae McIntosh, 1855



4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pike, K.S.; Starý, P.; Miller, T.; Allison, D.; Graf, G.; Boydston, L.; Miller, R.; Gillespie, R. Host range and habitats of the aphid parasitoid Diaeretiella rapae (Hymenoptera: Aphidiidae) in Washington State. Environ. Entomol. 1999, 28, 61–71. [Google Scholar] [CrossRef]
- Singh, R.; Singh, G. Systematics, distribution and host range of Diaeretiella rapae (McIntosh) (Hymenoptera: Braconidae, Aphidiinae). Int. J. Res. Stud. Biosci. 2015, 3, 1–36. [Google Scholar]
- Kavallieratos, N.G.; Tomanović, Ž.; Starý, P.; Athanassiou, C.G.; Sarlis, G.P.; Petrović, O.; Niketić, M.; Veroniki, M.A. A survey of aphid parasitoids (Hymenoptera: Braconidae: Aphidiinae) of Southeastern Europe and their aphid-plant associations. Appl. Entomol. Zool. 2004, 39, 527–563. [Google Scholar] [CrossRef]
- Zhang, W.Q.; Hassan, S.A. Use of the parasitoid Diaeretiella rapae (McIntoch) to control the cabbage aphid Brevicoryne brassicae (L.). J. Appl. Entomol. 2003, 127, 522–526. [Google Scholar] [CrossRef]
- Baker, D.A.; Loxdale, H.D.; Edwards, O.R. Genetic variation and founder effects in the parasitoid wasp, Diaeretiella rapae (M’intosh) (Hymenoptera: Braconidae: Aphidiidae), affecting its potential as a biological control agent. Mol. Ecol. 2003, 12, 3303–3311. [Google Scholar] [CrossRef]
- Desneux, N.; Fauvergue, X.; Dechaume-Moncharmont, F.X.; Kerhoas, L.; Ballanger, Y.; Kaiser, L. Diaeretiella rapae limits Myzus persicae populations after applications of deltamethrin in oilseed rape. J. Econ. Entomol. 2005, 98, 9–17. [Google Scholar] [CrossRef]
- Neuville, S.; Le Ralec, A.; Outreman, Y.; Jaloux, B. The delay in arrival of the parasitoid Diaeretiella rapae influences the efficiency of cabbage aphid biological control. BioControl 2016, 61, 115–126. [Google Scholar] [CrossRef]
- Soni, S.; Kumar, S. Biological control potential of an aphid parasitoid, Diaeretiella rapae (McIntosh) (Hymenoptera: Braconidae) against Brevicoryne brassicae (Linnaeus) (Hemiptera: Aphididae), a pest of oilseed brassicas in India. Int. J. Trop. Insect Sci. 2021, 41, 2361–2372. [Google Scholar] [CrossRef]
- Baer, C.F.; Tripp, D.W.; Bjorksten, T.A.; Antolin, M.F. Phylogeography of a parasitoid wasp (Diaeretiella rapae): No evidence of host-associated lineages. Mol. Ecol. 2004, 13, 1859–1869. [Google Scholar] [CrossRef]
- Hafez, M. Seasonal fluctuations of population density of the cabbage aphid, Brevicoryne brassicae (L.), in the Netherlands, and the role of its parasite, Aphidius (Diaeretiella) rapae (Curtis). Tijdschr. Plantenziekten 1961, 67, 345–548. [Google Scholar] [CrossRef]
- Sedlag, U. Zur Biologie und Bedeutung von Diaeretiella rapae (McIntosh) als Parasit der Kohlblattlaus (Brevicoryne brassicae L.). Nachrichtenbl. Dtsch. Pflanzenschutzdienstes 1964, 18, 81–86. [Google Scholar]
- Askari, A.; Alishah, A. Courtship behavior and evidence for a sex pheromone in Diaeretiella rapae (Hymenoptera: Braconidae), the cabbage aphid primary parasitoid. Ann. Entomol. Soc. Am. 1979, 72, 749–750. [Google Scholar] [CrossRef]
- Blande, J.D.; Pickett, J.A.; Poppy, G.M. Attack rate and success of the parasitoid Diaeretiella rapae on specialist and generalist feeding aphids. J. Chem. Ecol. 2004, 30, 1781–1795. [Google Scholar] [CrossRef]
- Martínez, M.Á.; Duarte, L.; Ceballos, M. Biology and vertical life table of Diaeretiella rapae McIntosh under laboratory conditions. Rev. Prot. Veg. 2013, 28, 23–26. [Google Scholar]
- Gazmer, R.; Gupta, M.K.; Singh, M.D. Biology of Diaeretiella rapae (McIntosh) (Hymenoptera: Aphidiidae) on cabbage aphid (Brevicoryne brassicae Linnaeus) and influence of host age on the developmental duration. J. Biol. Control 2015, 29, 38–42. [Google Scholar] [CrossRef]
- McIntosh, C. The Book of the Garden, 2nd ed.; William Blackwood and Sons: Edinburgh, Scotland; London, UK, 1855; p. 194. [Google Scholar]
- Curtis, J. Farm Insects: Being the Natural History and Economy of the Insects Injurious to the Field Crops of Great Britain and Ireland, and also Those Which Infest Barns and Granaries. With Suggestions for Their Destruction, 1st ed.; Blackie and Son: Glasgow, Scotland; London, UK, 1860; pp. 37–74. [Google Scholar]
- Mackauer, M. Comments on the authorship, generic position, and synonymy of Diaeretiella rapae (M‘Intosh) (Hymenoptera: Aphidiidae). Contrib. Entomol. 1964, 14, 53–58. [Google Scholar]
- Gahan, A.B. Some synonymy and other notes on Aphidiinae. Proc. Entomol. Soc. Wash. 1910, 12, 179–189. [Google Scholar]
- Marshall, T.A. A Catalogue of British Hymenoptera: Chrysididae, Ichneumonidae, Braconidae and Evaniidae; Entomolical Society of London: London, UK, 1872; p. 136. [Google Scholar]
- Starý, P. The generic classification of the family Aphidiidae (Hymenoptera). Acta Soc. Entomol. Czechoslov. 1960, 57, 238–252. [Google Scholar]
- Mackauer, M. Die Typen der Unterfamilie Aphidiinae des Britischen Museums London (Hymenoptera: Braconidae). Contrib. Entomol. 1961, 11, 96–154. [Google Scholar]
- Mackauer, M. Die Gattungen der Familie Aphidiidae und ihreverwandtschaftliche Zuordnung (Hymenoptera: Ichneumonoidea). Contrib. Entomol. 1961, 11, 792–803. [Google Scholar]
- Akhtar, M.S.; Dey, D.; Usmani, M.K. Redescription of Diaeretiella rapae (M’Intosh) (Hymenoptera: Braconidae: Aphidiinae) with morphological variability of several populations from India. Munis Entomol. Zool. 2011, 6, 194–203. [Google Scholar]
- Pramanik, A.; Dey, D.; Kumar, A. Redescription of Diaeretiella rapae (McIntosh) (hymenoptera: Braconidae: Aphidiinae) with emphasis on morphometrics. J. Entomol. Res. 2012, 36, 77–82. [Google Scholar]
- Herrick, G.W.; Hungate, J.W. The Cabbage Aphis; Cornell University: New York, NY, USA, 1911; Volume 300, p. 734. [Google Scholar]
- Ashmead, W.H. Descriptions of new Braconidae in the collection of the US National Museum. Proc. U.S. Natl. Mus. 1889, 11, 611–671. [Google Scholar] [CrossRef]
- Marshall, T.A. A Monograph of British Braconidae. Part VIII. Trans. R. Entomol. Soc. Lond. 1899, 47, 1–79. [Google Scholar] [CrossRef]
- Dalla Torre, K.W. Braconidae. In Catalogus Hymenopterorum; Guilelmi Engelmann: Leipzig, Germany, 1898; Volume 4. [Google Scholar]
- Marshall, T.A. Les Braconides. In Species des Hyménoptères d’Europe et d’Algérie; André, E., Ed.; Gray: Beaune, Germany, 1891. [Google Scholar]
- Mackauer, M.; Starý, P. Hymenoptera: Ichneumonoidea, World Aphidiidae. In Index of Entomophagous Insects; Delucchi, V., Remaudière, G., Eds.; Le Fransois: Paris, France, 1967. [Google Scholar]
- Mackauer, M. A re-examination of CF Baker’s collection of aphid parasites (Hymenoptera: Aphidiidae). Can. Entomol. 1963, 95, 921–935. [Google Scholar] [CrossRef]
- Gahan, A.B. Aphidiinae of North America. Agric. Exp. Stn. Maryland College Park Bull. 1911, 152, 147–200. [Google Scholar]
- Starý, P. A revision of the genus Diaeretiella Starý (Hymenoptera: Aphidiidae). Acta Entomol.Musei Natl. Pragae. 1961, 34, 383–397. [Google Scholar]
- Mackauer, M. Hymenopterorum Catalogus. Pars 3. Aphidiidae; Junk: Gravenhage, Netherlands, 1968. [Google Scholar]
- Starý, P. Aphid parasites of Czechoslovakia. A review of the Czechoslovak Aphidiidae (Hymenoptera); Springer: Hague, The Netherlands, 1966; p. 242. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Tomanović, Ž.; Žikić, V.; Petrović, A. Aphidius Nees (Hymenoptera, Braconidae, Aphidiinae) in Serbia: Key to species identification including parasitoid–aphid host list. Acta Entomol. Serbica 2023, 28, 61–76. [Google Scholar]
- Sharkey, M.J.; Wharton, R.A. Morphology and terminology. In Manual of the New World Genera of the Family Braconidae; Wharton, R.A., Marsh, P.M., Sharkey, M.J., Eds.; International Society of Hymenopterists: Washington, DC, USA, 1997. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol 1994, 3, 294–299. [Google Scholar]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-Del Barrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsis, S.E.; Sanchez-Gracia, A. DnaSP v6: DNA sequence polymorphism analysis of large datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, syy032. [Google Scholar] [CrossRef] [PubMed]
- Xia, X. 2018. DAMBE7: New and improved tools for data analysis in molecular biology and evolution. Mol. Biol. Evol. 2018, 35, 1550–1552. [Google Scholar] [CrossRef]
- Xia, X.; Xie, Z.; Salemi, M.; Chen, L.; Wang, Y. An index of substitution saturation and its application. Mol. Phylogenet. Evol. 2003, 26, 1–7. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree, version 1.4.4; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, Scotland, 2009. [Google Scholar]
- Tomanović, Ž.; Rakhshani, E.; Starý, P.; Kavallieratos, N.G.; Stanisavljević, L.Ž.; Žikić, V.; Athanassiou, C.G. Phylogenetic relationships between the genera Aphidius and Lysaphidus (Hymenoptera: Braconidae: Aphidiinae) with description of Aphidius iranicus sp. nov. The Canadian Entomologist 2007, 139, 297–307. [Google Scholar] [CrossRef]
- Ferreri, M.; Qu, W.; Han, B.O. Phylogenetic networks: A tool to display character conflict and demographic history. Afr. J. Biotechnol. 2011, 10, 12799–12803. [Google Scholar]
- Elliott, N.C.; Burd, J.D.; Armstrong, J.; Walker, C.B.; Reed, D.K.; Peairs, F.B. Release and recovery of imported parasitoids of the Russian wheat aphid in eastern Colorado. Southwest. Entomol. 1995, 20, 125–129. [Google Scholar]
- Brewer, M.J.; Nelson, D.J.; Ahern, R.G.; Donahue, J.D.; Prokrym, D.R. Recovery and range expansion of parasitoids (Hymenoptera: Aphelinidae and Braconidae) released for biological control of Diuraphis noxia (Homoptera: Aphididae) in Wyoming. Environ. Entomol. 2001, 30, 578–588. [Google Scholar] [CrossRef]
- Tanigoshi, L.K.; Pike, K.S.; Miller, R.H.; Miller, T.D.; Allison, D. Search for, and release of, parasitoids for the biological control of Russian wheat aphid in Washington State (USA). Agric. Ecosyst. Environ. 1995, 52, 25–30. [Google Scholar] [CrossRef]
- Carver, M.; Starý, P. A preliminary review of the Aphidiidae (Hymenoptera: Ichneumonoidea) of Australia and New Zealand. Aust. J. Entomol. 1974, 13, 235–240. [Google Scholar] [CrossRef]
- Teulon, D.A.J.; Drayton, G.M.; Scott, I.A.W. Exotic introductions of primary parasitoids of aphids in New Zealand: The good and the bad. In Proceedings of the Third International Symposium on Biological Control of Arthropods, Christchurch, New Zealand, 8–13 February 2008; pp. 421–430. [Google Scholar]
- Žikić, V.; Stanković, S.S.; Petrović, A.; Ilić Milošević, M.; Tomanović, Ž.; Klingenberg, C.P.; Ivanović, A. Evolutionary relationships of wing venation and wing size and shape in Aphidiinae (Hymenoptera: Braconidae). Org. Divers. Evol. 2017, 17, 607–617. [Google Scholar] [CrossRef]
- Peters, R.S.; Krogmann, L.; Mayer, C.; Donath, A.; Gunkel, S.; Meusemann, K.; Kolozov, A.; Podsiadlowski, L.; Petersen, M.; Lanfear, R.; et al. Evolutionary history of the Hymenoptera. Curr. Biol. 2017, 27, 1013–1018. [Google Scholar] [CrossRef]
- Polilov, A.A. Small is beautiful: Features of the smallest insects and limits to miniaturization. Annu. Rev. Entomol. 2015, 60, 103–121. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popović, N.; Kocić, K.; Tomanović, Ž.; Petrović, A. Alpha, Beta and Gamma Taxonomy of Biocontrol Agent Diaeretiella rapae (Hymenoptera, Braconidae). Insects 2025, 16, 736. https://doi.org/10.3390/insects16070736
Popović N, Kocić K, Tomanović Ž, Petrović A. Alpha, Beta and Gamma Taxonomy of Biocontrol Agent Diaeretiella rapae (Hymenoptera, Braconidae). Insects. 2025; 16(7):736. https://doi.org/10.3390/insects16070736
Chicago/Turabian StylePopović, Nemanja, Korana Kocić, Željko Tomanović, and Andjeljko Petrović. 2025. "Alpha, Beta and Gamma Taxonomy of Biocontrol Agent Diaeretiella rapae (Hymenoptera, Braconidae)" Insects 16, no. 7: 736. https://doi.org/10.3390/insects16070736
APA StylePopović, N., Kocić, K., Tomanović, Ž., & Petrović, A. (2025). Alpha, Beta and Gamma Taxonomy of Biocontrol Agent Diaeretiella rapae (Hymenoptera, Braconidae). Insects, 16(7), 736. https://doi.org/10.3390/insects16070736