Effects of Natural Habitat and Season on Cursorial Spider Assemblages in Mediterranean Vineyards

 , , ,
, , ,  and
and
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
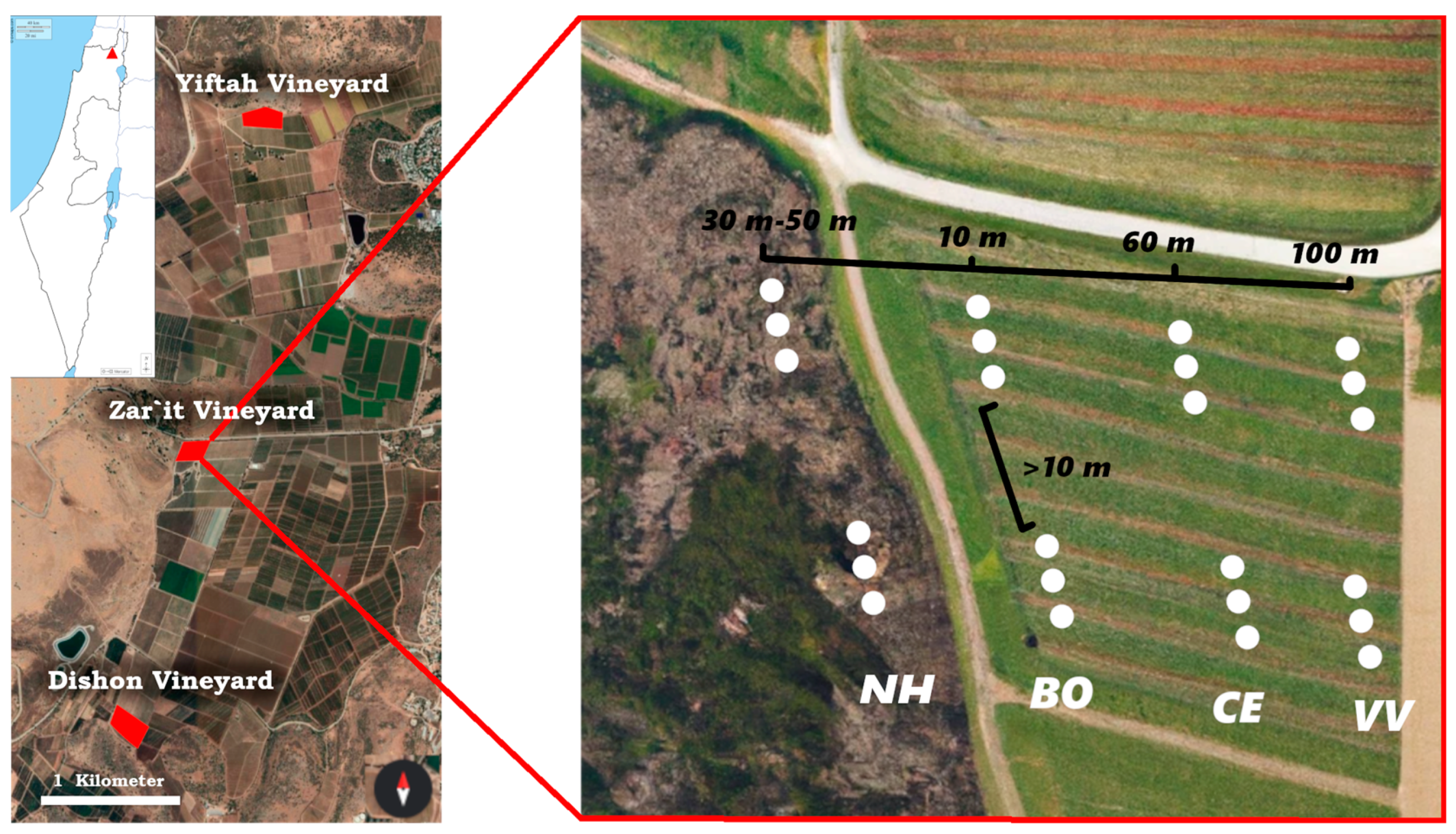
2.1. Study Sites
2.2. Spider Sampling
2.3. Data Analysis
3. Results
3.1. Spider Abundance
3.2. Spider Richness
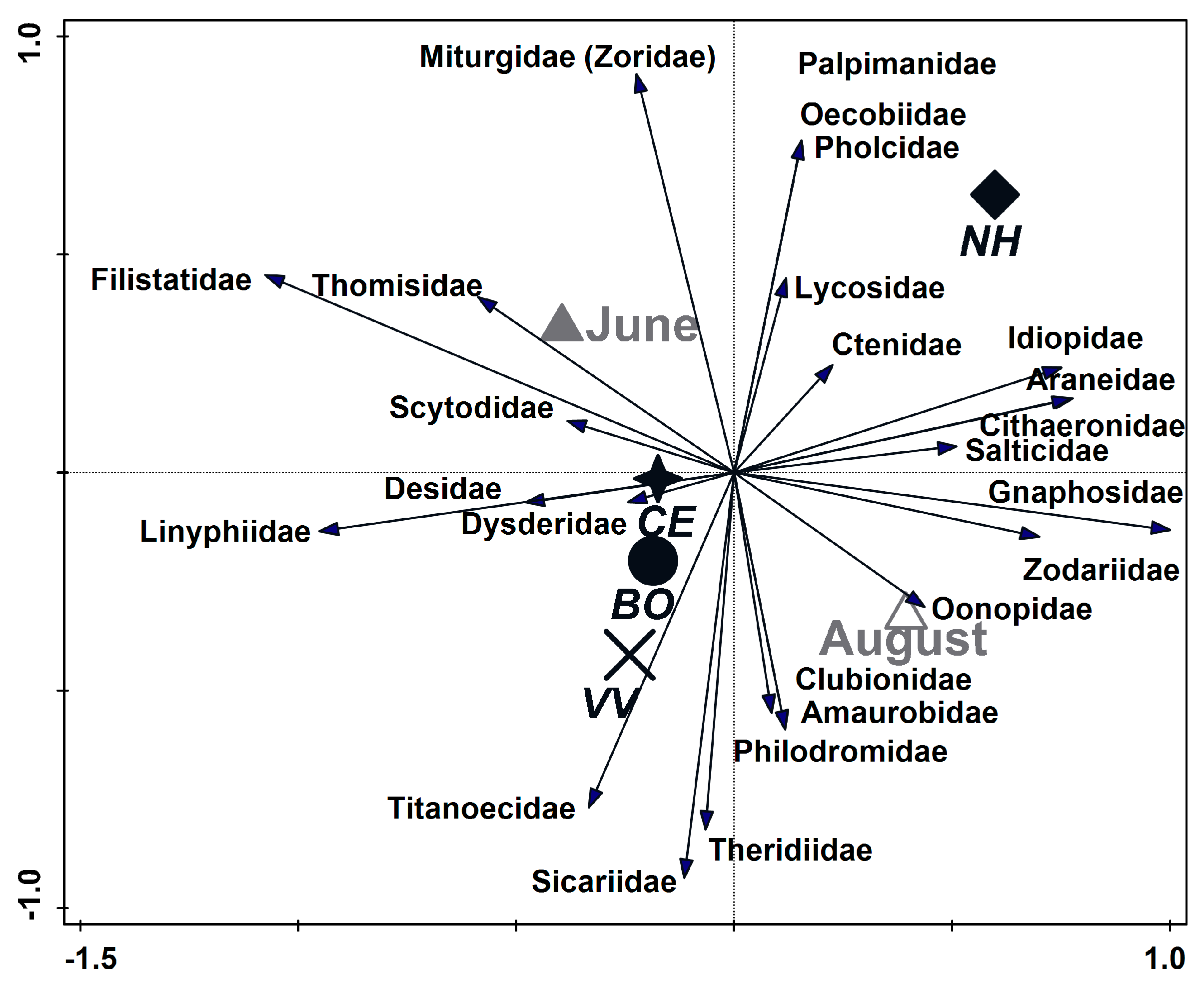
3.3. Spider Species and Family Composition
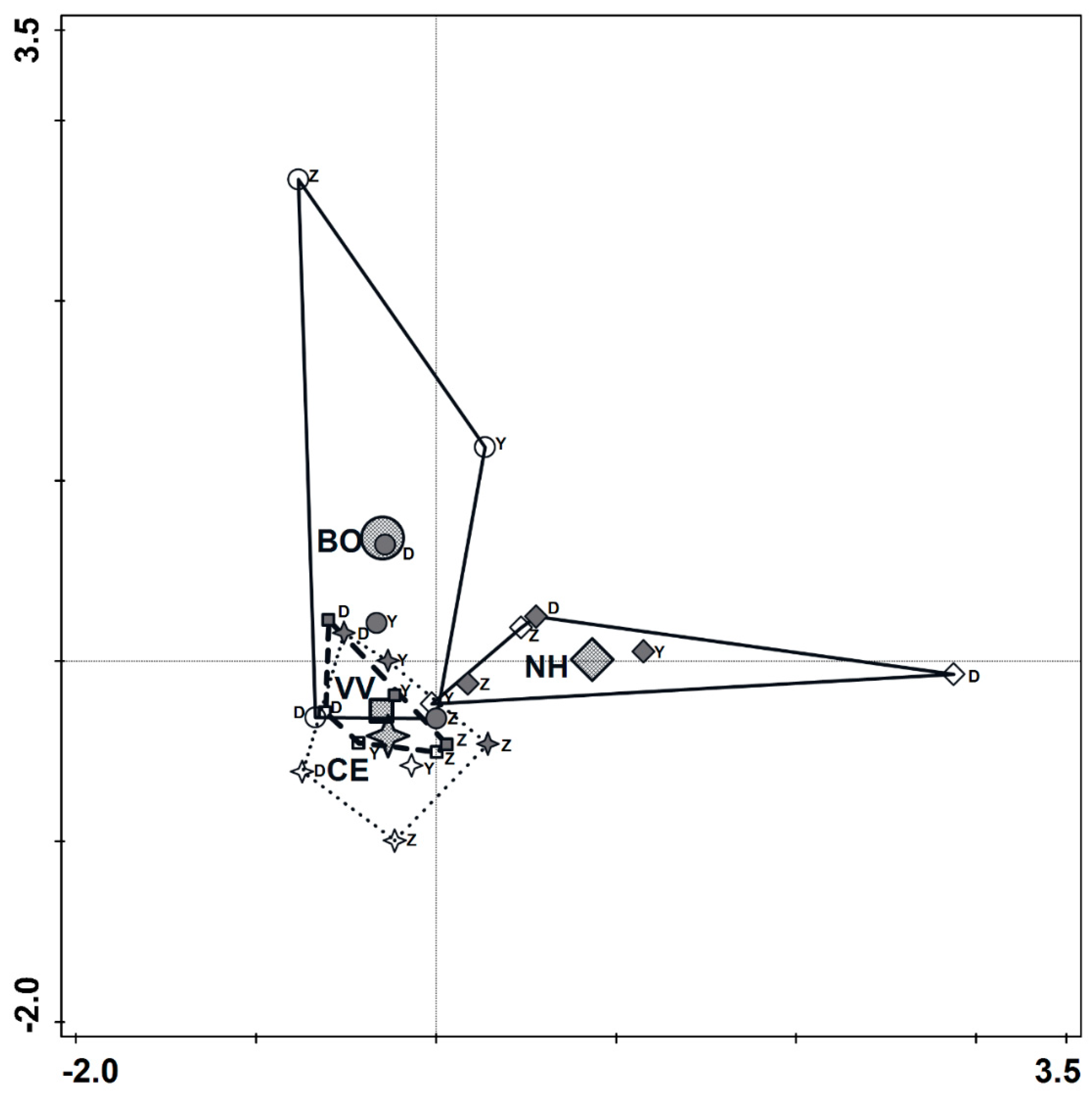
3.4. Spider Diversity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| June | August | ||||||
|---|---|---|---|---|---|---|---|
| Genus or Species | Dishon | Yiftah | Zar’it | Dishon | Yiftah | Zar’it | |
| Amaurobiidae | Amaurobius (C. L. Koch, 1837) | 1 | |||||
| Araneidae | Singa neta (O. Pickard-Cambridge, 1872) | 1 | 2 | ||||
| Cithaeronidae | Cithaeron praedonius (O. Pickard-Cambridge, 1872) | 2 | |||||
| Clubionidae | Clubiona (Latreille, 1804) | 1 | |||||
| Ctenidae | Anahita syriaca (O. Pickard-Cambridge, 1872) | 1 | 1 | ||||
| Desidae | Desid sp. | 1 | |||||
| Dysderidae | Dysdera (Latreille, 1804) | 1 | 2 | 3 | |||
| Harpactea (Bristowe, 1939) | 1 | ||||||
| Filistatidae | Pritha albimaculata (O. Pickard-Cambridge, 1872) | 3 | 5 | ||||
| Zaitunia schmitzi (Kulczyński, 1911) | 1 | ||||||
| Gnaphosidae | Civizelotes solstitialis (Levy, 1998) | 3 | 3 | ||||
| Heser aradensis (Levy, 1998) | 2 | 3 | |||||
| Heser nilicola (O. Pickard-Cambridge, 1874) | 4 | 7 | |||||
| Marinarozelotes miniglossus (Levy, 2009) | 1 | ||||||
| Micaria ignea (O. Pickard-Cambridge, 1872) | 17 | ||||||
| Pterotricha cambridgei (L. Koch, 1872) | 1 | ||||||
| Pterotricha engediensis (Levy, 1995) | 1 | ||||||
| Pterotricha levantina (Levy, 1995) | 2 | 3 | 4 | 1 | 25 | 16 | |
| Pterotricha parasyriaca (Levy, 1995) | 1 | ||||||
| Zelotes paradderet (Levy, 2009) | 1 | 1 | |||||
| Zelotes zin (Levy, 1998) | 4 | 6 | 1 | ||||
| Gnaphosid spp. (juveniles) | 8 | 5 | 5 | 8 | 16 | 9 | |
| Gnaphosid sp. D | 1 | ||||||
| Gnaphosid sp. E | 1 | ||||||
| Gnaphosid sp. F | 1 | ||||||
| Gnaphosid sp. G | 1 | ||||||
| Idiopidae | Titanidiops syriacus (O. Pickard-Cambridge, 1870) | 1 | |||||
| Linyphiidae | Agyneta pseudorurestris (Wunderlich, 1980) | 1 | 7 | 1 | |||
| Linyphiid spp. (juveniles) | 7 | 4 | 4 | 5 | 4 | 2 | |
| Linyphiid sp. A | 1 | 2 | |||||
| Linyphiid sp. B | 1 | 1 | 5 | ||||
| Linyphiid sp. C | 1 | ||||||
| Lycosidae | Alopecosa albofasciata (Brullé, 1832) | 1 | 1 | 2 | 2 | ||
| Hogna graeca (Roewer, 1951) | 10 | 5 | 3 | 5 | 3 | 4 | |
| Lycosa piochardi Simon, 1876 | 2 | 1 | 5 | 3 | 1 | ||
| Pardosa subsordidatula (Strand, 1915) | 8 | 4 | 1 | 1 | |||
| Lycosid spp. (juveniles) | 9 | 3 | 1 | 1 | |||
| Miturgidae | Prochora lycosiformis (O. Pickard-Cambridge, 1872) | 3 | 2 | ||||
| Oecobiidae | Oecobius (Lucas, 1846) | 4 | |||||
| Oonopidae | Oonopid spp. (juveniles) | 3 | 1 | ||||
| Palpimanidae | Palpimanus sogdianus (Charitonov, 1946) | 2 | |||||
| Philodromidae | Thanatus vulgaris (Simon, 1870) | 9 | 2 | 1 | 9 | 16 | |
| Oonopid spp. (juveniles) | 6 | 5 | 6 | 2 | 8 | 9 | |
| Pholcidae | Pholcid spp. (juveniles) | 2 | |||||
| Salticidae | Aelurillus politiventris (O. Pickard-Cambridge, 1872) | 3 | 5 | 4 | 1 | 3 | |
| Euophrys (C. L. Koch, 1834) | 1 | ||||||
| Neaetha oculata (O. Pickard-Cambridge, 1876) | 1 | ||||||
| Pellenes nigrociliatus (Simon, 1875) | 1 | ||||||
| Pseudicius amicus (Prószyński, 2000) | 1 | ||||||
| Rafalus (Prószyński, 1999) | 1 | ||||||
| Salticid spp. (juveniles) | 9 | 11 | 6 | 18 | 3 | 15 | |
| Scytodidae | Scytodes thoracica (Latreille, 1802) | 4 | 3 | ||||
| Scytodid spp. (juveniles) | 5 | ||||||
| Sicariidae | Loxosceles rufescens (Dufour, 1820) | 1 | 2 | 5 | 3 | ||
| Theridiidae | Euryopis episinoides (Walckenaer, 1847) | 1 | |||||
| Theridiid spp. (juveniles) | 1 | 1 | 1 | ||||
| Theridion negebense (Levy & Amitai, 1982) | 2 | ||||||
| Steatoda erigoniformis (O. Pickard-Cambridge, 1872) | 1 | ||||||
| Theridiid sp. H | 1 | ||||||
| Thomisidae | Ozyptila (Simon, 1864) | 1 | |||||
| Xysticus (C. L. Koch, 1835) | 1 | 1 | |||||
| Thomisid spp. (juveniles) | 2 | 2 | 2 | ||||
| Titanoecidae | Nurscia albomaculata (Lucas, 1846) | 1 | 1 | 1 | 2 | 2 | |
| Titanoecid sp. (juvenile) | 1 | ||||||
| Zodariidae | Lachesana (Strand, 1932) | 1 | |||||
| Palaestina eremica (Levy, 1992) | 3 | ||||||
| Palaestina expolita (O. Pickard-Cambridge, 1872) | 5 | 1 | 1 | 1 | 4 | ||
| Palaestina spp. (juveniles) | 1 | 1 | |||||
| Pax libani (Simon, 1873) | 1 | ||||||
| Pax palmonii (Levy, 1990) | 1 | ||||||
| Trygetus sexoculatus (O. Pickard-Cambridge, 1872) | 2 | 1 | 1 | ||||
| Zodarion geshur (Levy, 2007) | 4 | 3 | |||||
| Zodarion lutipes (O. Pickard-Cambridge, 1872) | 2 | 1 | 2 | 15 | |||
| Zodariid spp. (juveniles) | 7 | 4 | 6 | 32 | |||
| Araneae | Unidentified spiders (juveniles and poorly preserved) | 28 | 7 | 7 | 1 | 10 | 3 |
References
- Heimpel, G.E.; Mills, N.J. Biological Control: Ecology and Applications; Cambridge University Press: New York, NY, USA, 2017; ISBN 978-0-521-84514-4. [Google Scholar]
- Holland, J.M.; Douma, J.C.; Crowley, L.; James, L.; Kor, L.; Stevenson, D.R.W.; Smith, B.M. Semi-Natural Habitats Support Biological Control, Pollination and Soil Conservation in Europe. A Review. Agron. Sustain. Dev. 2017, 37, 31. [Google Scholar] [CrossRef]
- Hendrickx, F.; Maelfait, J.-P.; Van Wingerden, W.; Schweiger, O.; Speelmans, M.; Aviron, S.; Augenstein, I.; Billeter, R.; Bailey, D.; Bukacek, R.; et al. How Landscape Structure, Land-Use Intensity and Habitat Diversity Affect Components of Total Arthropod Diversity in Agricultural Landscapes: Agricultural Factors and Arthropod Biodiversity. J. Appl. Ecol. 2007, 44, 340–351. [Google Scholar] [CrossRef]
- Dainese, M.; Martin, E.A.; Aizen, M.A.; Albrecht, M.; Bartomeus, I.; Bommarco, R.; Carvalheiro, L.G.; Chaplin-Kramer, R.; Gagic, V.; Garibaldi, L.A.; et al. A Global Synthesis Reveals Biodiversity-Mediated Benefits for Crop Production. Sci. Adv. 2019, 5, eaax0121. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Haan, N.L.; Landis, D.A. Landscape Composition and Configuration Have Scale-Dependent Effects on Agricultural Pest Suppression. Agric. Ecosyst. Environ. 2020, 302, 107085. [Google Scholar] [CrossRef]
- Letourneau, D.K.; Armbrecht, I.; Rivera, B.S.; Lerma, J.M.; Carmona, E.J.; Daza, M.C.; Escobar, S.; Galindo, V.; Gutiérrez, C.; López, S.D.; et al. Does Plant Diversity Benefit Agroecosystems? A Synthetic Review. Ecol. Appl. 2011, 21, 9–21. [Google Scholar] [CrossRef]
- Álvarez, H.A.; Morente, M.; Oi, F.S.; Rodríguez, E.; Campos, M.; Ruano, F. Semi-Natural Habitat Complexity Affects Abundance and Movement of Natural Enemies in Organic Olive Orchards. Agric. Ecosyst. Environ. 2019, 285, 106618. [Google Scholar] [CrossRef]
- Mota, L.; Hevia, V.; Rad, C.; Alves, J.; Silva, A.; González, J.A.; Ortega-Marcos, J.; Aguado, O.; Alcorlo, P.; Azcárate, F.M.; et al. Flower Strips and Remnant Semi-natural Vegetation Have Different Impacts on Pollination and Productivity of Sunflower Crops. J. Appl. Ecol. 2022, 59, 2386–2397. [Google Scholar] [CrossRef]
- Birkhofer, K.; Entling, M.H.; Lubin, Y. Agroecology: Trait Composition, Spatial Relationships, Trophic Interactions. In Spider Research in the 21st Century: Trends & perspectives; Penney, D., Ed.; Siri Scientific Press: Rochdale, UK, 2013; pp. 200–228. [Google Scholar]
- Nyffeler, M.; Birkhofer, K. An Estimated 400–800 Million Tons of Prey Are Annually Killed by the Global Spider Community. Sci. Nat. 2017, 104, 30. [Google Scholar] [CrossRef]
- Pekár, S.; Wolff, J.O.; Černecká, Ľ.; Birkhofer, K.; Mammola, S.; Lowe, E.C.; Fukushima, C.S.; Herberstein, M.E.; Kučera, A.; Buzatto, B.A.; et al. The World Spider Trait Database: A Centralized Global Open Repository for Curated Data on Spider Traits. Database 2021, 2021, baab064. [Google Scholar] [CrossRef]
- Birkhofer, K.; Gavish-Regev, E.; Endlweber, K.; Lubin, Y.D.; von Berg, K.; Wise, D.H.; Scheu, S. Cursorial Spiders Retard Initial Aphid Population Growth at Low Densities in Winter Wheat. Bull. Entomol. Res. 2008, 98, 249–255. [Google Scholar] [CrossRef]
- Gavish-Regev, E.; Lubin, Y.; Coll, M. Migration Patterns and Functional Groups of Spiders in a Desert Agroecosystem. Ecol. Entomol. 2008, 33, 202–212. [Google Scholar] [CrossRef]
- Chapman, E.G.; Schmidt, J.M.; Welch, K.D.; Harwood, J.D. Molecular Evidence for Dietary Selectivity and Pest Suppression Potential in an Epigeal Spider Community in Winter Wheat. Biol. Control 2013, 65, 72–86. [Google Scholar] [CrossRef]
- Li, X.; Liu, Y.; Duan, M.; Yu, Z.; Axmacher, J.C. Different Response Patterns of Epigaeic Spiders and Carabid Beetles to Varying Environmental Conditions in Fields and Semi-Natural Habitats of an Intensively Cultivated Agricultural Landscape. Agric. Ecosyst. Environ. 2018, 264, 54–62. [Google Scholar] [CrossRef]
- Twardowski, J.P.; Gruss, I.A.; Hurej, M. Can Mixture of Flowering Plants within Intensive Agricultural Landscape Positively Affect Ground-Dwelling Spider Assemblages? Rom. Agric. Res. 2021, 38, 345–355. [Google Scholar] [CrossRef]
- Michalko, R.; Pekár, S.; Entling, M.H. An Updated Perspective on Spiders as Generalist Predators in Biological Control. Oecologia 2019, 189, 21–36. [Google Scholar] [CrossRef]
- Riechert, S.E.; Bishop, L. Prey Control by an Assemblage of Generalist Predators: Spiders in Garden Test Systems. Ecology 1990, 71, 1441–1450. [Google Scholar] [CrossRef]
- Blandenier, G. Ballooning of Spiders (Araneae) in Switzerland: General Results from an Eleven-Year Survey. Arachnology 2009, 14, 308–316. [Google Scholar] [CrossRef]
- Pluess, T.; Opatovsky, I.; Gavish-Regev, E.; Lubin, Y.; Schmidt, M.H. Spiders in Wheat Fields and Semi-Desert in the Negev (Israel). J. Arachnol. 2008, 36, 368–373. [Google Scholar] [CrossRef]
- Sunderland, K.; Samu, F. Effects of Agricultural Diversification on the Abundance, Distribution, and Pest Control Potential of Spiders: A Review. Entomol. Exp. Appl. 2000, 95, 1–13. [Google Scholar] [CrossRef]
- Samu, F. Can Field-Scale Habitat Diversification Enhance the Biocontrol Potential of Spiders? Pest. Manag. Sci. 2003, 59, 437–442. [Google Scholar] [CrossRef]
- Nardi, D.; Lami, F.; Pantini, P.; Marini, L. Using Species-Habitat Networks to Inform Agricultural Landscape Management for Spiders. Biol. Conserv. 2019, 239, 108275. [Google Scholar] [CrossRef]
- Lubin, Y.; Ferrante, M.; Musli, I.; Lövei, G.L. Diversity of Ground-Active Spiders in Negev Desert Habitats, Israel. J. Arid Environ. 2020, 183, 104252. [Google Scholar] [CrossRef]
- Lin, S.; Vasseur, L.; You, M.-S. Seasonal Variability in Spider Assemblages in Traditional and Transgenic Rice Fields. Environ. Entomol. 2016, 45, 537–546. [Google Scholar] [CrossRef]
- Bao, L.; Ginella, J.; Cadenazzi, M.; Castiglioni, E.; Martínez, S.; Casales, L.; Caraballo, M.; Laborda, Á.; Simo, M. Spider Assemblages Associated with Different Crop Stages of Irrigated Rice Agroecosystems from Eastern Uruguay. BDJ 2018, 6, e24974. [Google Scholar] [CrossRef]
- Salman, I.N.A.; Lubin, Y.; Gavish-Regev, E. Multiple Factors at Regional, Landscape, and Local Scales Determine Spider Assemblage Composition in Pomegranate Orchards. Agriculture 2022, 12, 512. [Google Scholar] [CrossRef]
- Gaigher, R.; Samways, M.J. Landscape Mosaic Attributes for Maintaining Ground-Living Spider Diversity in a Biodiversity Hotspot. Insect Conserv. Divers. 2014, 7, 470–479. [Google Scholar] [CrossRef]
- Nascimbene, J.; Marini, L.; Ivan, D.; Zottini, M. Management Intensity and Topography Determined Plant Diversity in Vineyards. PLoS ONE 2013, 8, e76167. [Google Scholar] [CrossRef] [PubMed]
- Möller, G.; Keasar, T.; Shapira, I.; Möller, D.; Ferrante, M.; Segoli, M. Effect of Weed Management on the Parasitoid Community in Mediterranean Vineyards. Biology 2020, 10, 7. [Google Scholar] [CrossRef] [PubMed]
- Isaia, M.; Bona, F.; Badino, G. Influence of Landscape Diversity and Agricultural Practices on Spider Assemblage in Italian Vineyards of Langa Astigiana (Northwest Italy). Environ. Entomol. 2006, 35, 297–307. [Google Scholar] [CrossRef]
- Kolb, S.; Uzman, D.; Leyer, I.; Reineke, A.; Entling, M.H. Differential Effects of Semi-Natural Habitats and Organic Management on Spiders in Viticultural Landscapes. Agric. Ecosyst. Environ. 2020, 287, 106695. [Google Scholar] [CrossRef]
- Gonçalves, F.; Nunes, C.; Carlos, C.; López, Á.; Oliveira, I.; Crespí, A.; Teixeira, B.; Pinto, R.; Costa, C.A.; Torres, L. Do Soil Management Practices Affect the Activity Density, Diversity, and Stability of Soil Arthropods in Vineyards? Agric. Ecosyst. Environ. 2020, 294, 106863. [Google Scholar] [CrossRef]
- Geldenhuys, M.; Gaigher, R.; Pryke, J.S.; Samways, M.J. Interplay between Amount of Natural Vegetation and Effective Vineyard Management Practices Benefits Local Arthropod Diversity. Biodivers. Conserv. 2022, 31, 3039–3053. [Google Scholar] [CrossRef]
- D’Alberto, C.F.; Hoffmann, A.A.; Thomson, L.J. Limited Benefits of Non-Crop Vegetation on Spiders in Australian Vineyards: Regional or Crop Differences? BioControl 2012, 57, 541–552. [Google Scholar] [CrossRef]
- Eckert, M.; Mathulwe, L.L.; Gaigher, R.; Joubert-van der Merwe, L.; Pryke, J.S. Native Cover Crops Enhance Arthropod Diversity in Vineyards of the Cape Floristic Region. J. Insect Conserv. 2020, 24, 133–149. [Google Scholar] [CrossRef]
- Hogg, B.N.; Daane, K.M. The Role of Dispersal from Natural Habitat in Determining Spider Abundance and Diversity in California Vineyards. Agric. Ecosyst. Environ. 2010, 135, 260–267. [Google Scholar] [CrossRef]
- Shapira, I.; Gavish-Regev, E.; Sharon, R.; Harari, A.R.; Kishinevsky, M.; Keasar, T. Habitat Use by Crop Pests and Natural Enemies in a Mediterranean Vineyard Agroecosystem. Agric. Ecosyst. Environ. 2018, 267, 109–118. [Google Scholar] [CrossRef]
- Shapira, I.; Keasar, T.; Harari, A.R.; Gavish-Regev, E.; Kishinevsky, M.; Steinitz, H.; Sofer-Arad, C.; Tomer, M.; Avraham, A.; Sharon, R. Does Mating Disruption of Planococcus Ficus and Lobesia Botrana Affect the Diversity, Abundance and Composition of Natural Enemies in Israeli Vineyards?: Non-Target Effects of Mating Disruption on Natural Enemies. Pest. Manag. Sci. 2018, 74, 1837–1844. [Google Scholar] [CrossRef]
- Nentwig, W.; Blick, T.; Bosmans, R.; Gloor, D.; Hänggi, A.; Kropf, C. Spiders of Europe. Version 08.2023. Available online: https://araneae.nmbe.ch/ (accessed on 11 August 2023).
- Roberts, M. Collins Field Guide to the Spiders of Britain and Northern Europe; Harper Collins: London, UK, 1996. [Google Scholar]
- ter Braak, C.; Smilauer, P. Canoco—Software for Ordination Version 5.10. 2020. Available online: http://www.canoco5.com/index.php (accessed on 11 August 2023).
- ter Braak, C.; Smilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination; Version 5.10; Microcomputer Power: Ithaca, NY, USA, 2018. [Google Scholar]
- Tóthmérész, B. Comparison of Different Methods for Diversity Ordering. J. Veg. Sci. 1995, 6, 283–290. [Google Scholar] [CrossRef]
- Legendre, L.; Legendre, P. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 2012; ISBN 978-0-444-53868-0. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. Vegan: Community Ecology Package. R Package Version 2.6-4. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 8 September 2023).
- Zonstein, S.; Marusik, Y.M. Checklist of the Spiders (Araneae) of Israel. Zootaxa 2013, 3671, 1. [Google Scholar] [CrossRef]
- Pekár, S.; Šobotník, J.; Lubin, Y. Armoured Spiderman: Morphological and Behavioural Adaptations of a Specialised Araneophagous Predator (Araneae: Palpimanidae). Naturwissenschaften 2011, 98, 593–603. [Google Scholar] [CrossRef]
- Pekár, S.; Lubin, Y. Habitats and Interspecific Associations of Zodariid Spiders in the Negev (Araneae: Zodariidae). Isr. J. Zool. 2003, 49, 255–267. [Google Scholar] [CrossRef]
- Opatovsky, I.; Lubin, Y. Coping with Abrupt Decline in Habitat Quality: Effects of Harvest on Spider Abundance and Movement. Acta Oecol. 2012, 41, 14. [Google Scholar] [CrossRef]
- World Spider Catalog Version 23.5. Available online: http://wsc.nmbe.ch (accessed on 8 September 2023).
- Begg, G.S.; Cook, S.M.; Dye, R.; Ferrante, M.; Franck, P.; Lavigne, C.; Lövei, G.L.; Mansion-Vaquie, A.; Pell, J.K.; Petit, S.; et al. A Functional Overview of Conservation Biological Control. Crop Prot. 2017, 97, 145–158. [Google Scholar] [CrossRef]
- Albrecht, M.; Kleijn, D.; Williams, N.M.; Tschumi, M.; Blaauw, B.R.; Bommarco, R.; Campbell, A.J.; Dainese, M.; Drummond, F.A.; Entling, M.H.; et al. The Effectiveness of Flower Strips and Hedgerows on Pest Control, Pollination Services and Crop Yield: A Quantitative Synthesis. Ecol. Lett. 2020, 23, 1488–1498. [Google Scholar] [CrossRef] [PubMed]
- Decocq, G.; Andrieu, E.; Brunet, J.; Chabrerie, O.; De Frenne, P.; De Smedt, P.; Deconchat, M.; Diekmann, M.; Ehrmann, S.; Giffard, B.; et al. Ecosystem Services from Small Forest Patches in Agricultural Landscapes. Curr. For. Rep. 2016, 2, 30–44. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ganem, Z.; Ferrante, M.; Lubin, Y.; Armiach Steinpress, I.; Gish, M.; Sharon, R.; Harari, A.R.; Keasar, T.; Gavish-Regev, E. Effects of Natural Habitat and Season on Cursorial Spider Assemblages in Mediterranean Vineyards. Insects 2023, 14, 782. https://doi.org/10.3390/insects14100782
Ganem Z, Ferrante M, Lubin Y, Armiach Steinpress I, Gish M, Sharon R, Harari AR, Keasar T, Gavish-Regev E. Effects of Natural Habitat and Season on Cursorial Spider Assemblages in Mediterranean Vineyards. Insects. 2023; 14(10):782. https://doi.org/10.3390/insects14100782
Chicago/Turabian StyleGanem, Zeana, Marco Ferrante, Yael Lubin, Igor Armiach Steinpress, Moshe Gish, Rakefet Sharon, Ally R. Harari, Tamar Keasar, and Efrat Gavish-Regev. 2023. "Effects of Natural Habitat and Season on Cursorial Spider Assemblages in Mediterranean Vineyards" Insects 14, no. 10: 782. https://doi.org/10.3390/insects14100782
APA StyleGanem, Z., Ferrante, M., Lubin, Y., Armiach Steinpress, I., Gish, M., Sharon, R., Harari, A. R., Keasar, T., & Gavish-Regev, E. (2023). Effects of Natural Habitat and Season on Cursorial Spider Assemblages in Mediterranean Vineyards. Insects, 14(10), 782. https://doi.org/10.3390/insects14100782