
Notes on the Genus Aceratoneuromyia Girault (Hymenoptera: Eulophidae) †


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Collection and Rearing
2.2. Specimens Preparations
2.3. Molecular Analyses
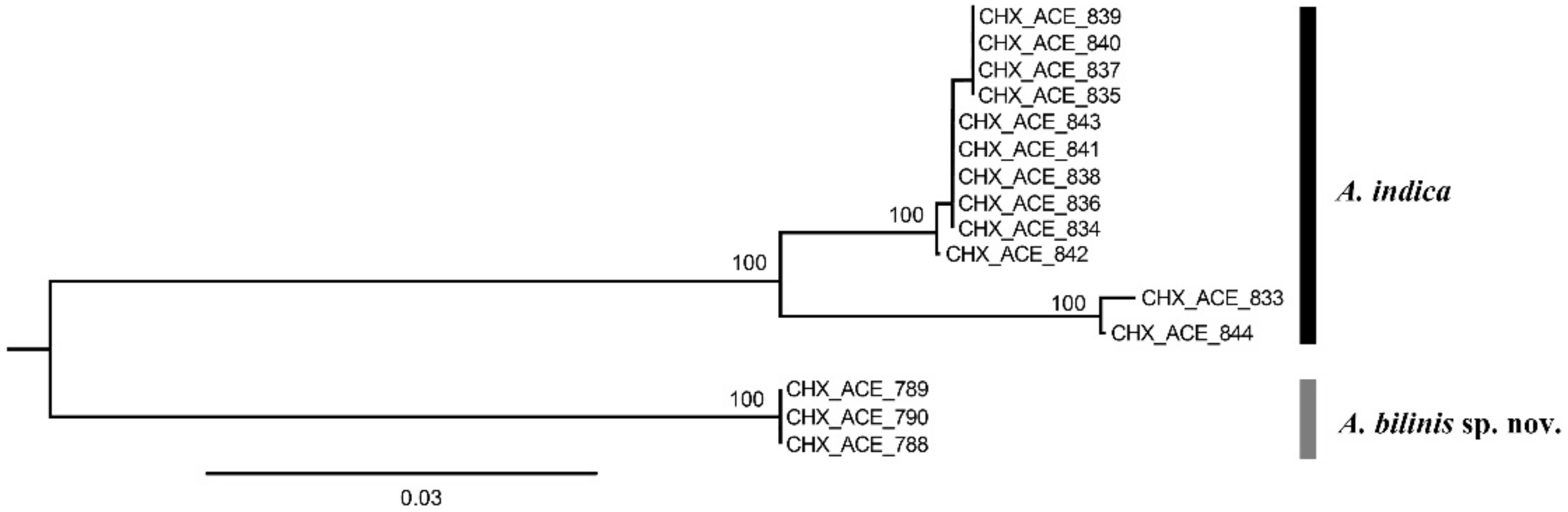
3. Results
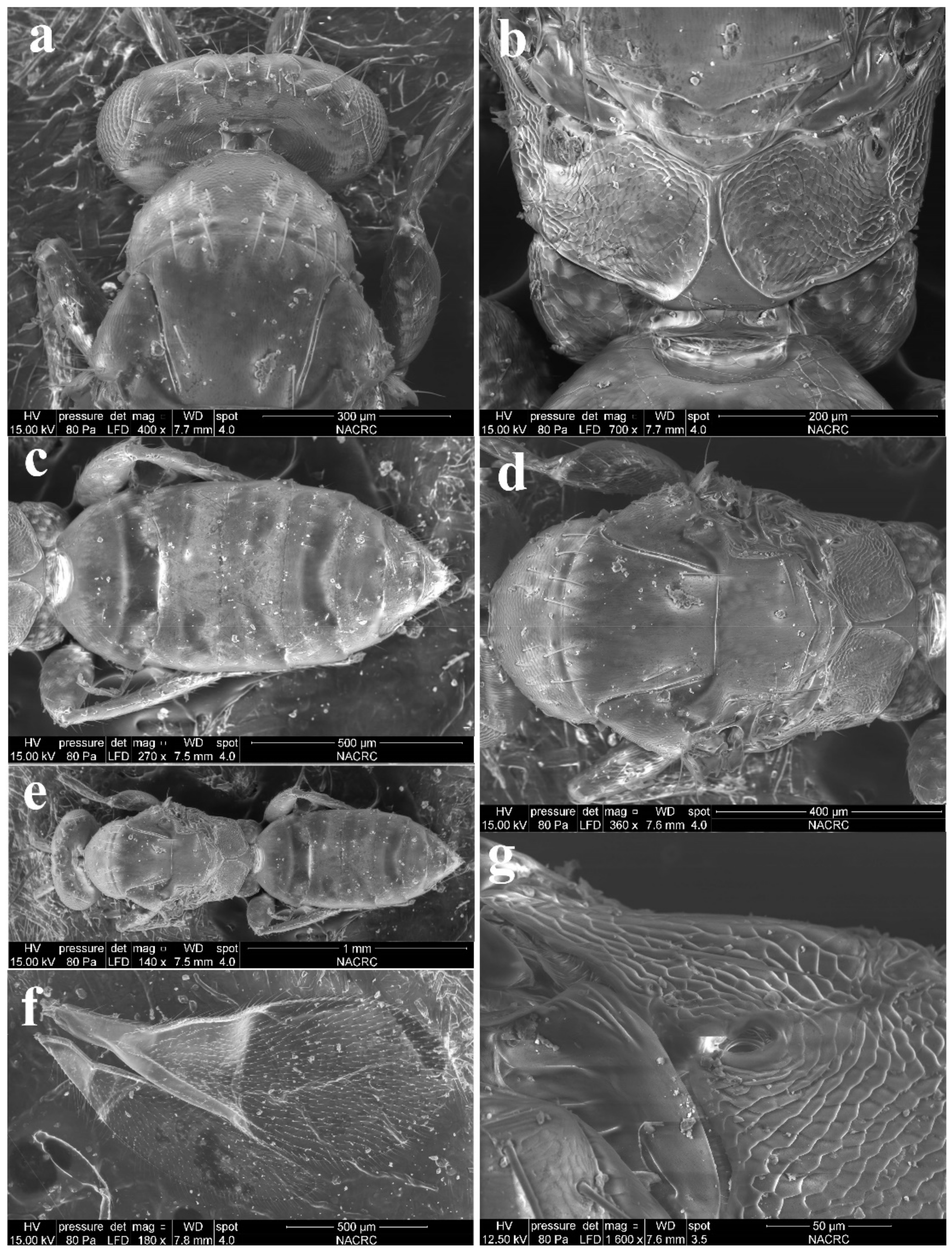
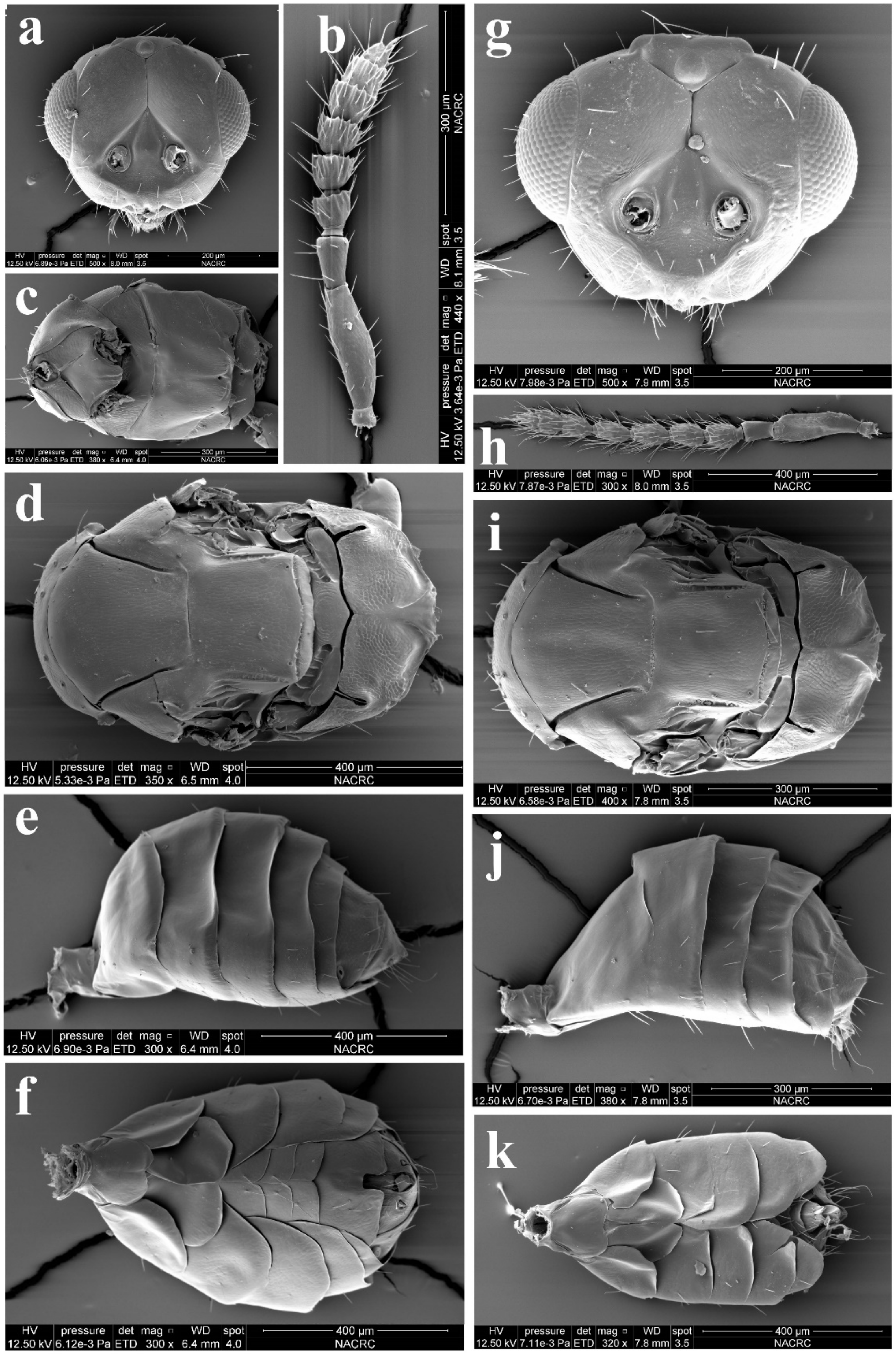
3.1. Morphological Diagnosis and Species Treatments of Aceratoneuromyia
3.1.1. Aceratoneuromyia bilinis Huangfu & Cao, sp. nov.
3.1.2. Aceratoneuromyia carinata Cao & Zhu, sp. nov.
3.1.3. Aceratoneuromyia indica (Silvestri, 1910)
3.1.4. Aceratoneuromyia trilinus Cao & Zhu, sp. nov.
3.2. Key to Species of Aceratoneuromyia
- 1.
- -
- 2(1)
- Pronotum with 2 long setae near its posterior margin; scutellum 2.15× as broad as long and with one pair of setae; propodeal spiracle separated by about 1.5× its diameter from metanotum; length of longest seta on vertex 1.3× maximum diameter of posterior ocellus; eye bare. Russia…A. lakicus (Kostjukov & Gunasheva)
- -
- Pronotum with 4 long setae near posterior margin; scutellum 1.3–1.4× as broad as long, with two pairs of setae; propodeal spiracle separated by less than its diameter from metanotum; eye with short, sparse setae…3
- 3(2)
- Antennal scape somewhat longer than an eye, reaching anterior ocellus; pronotum bare, except for a row of 4 long setae near its posterior margin and sometimes 2–3 short ones at sides; propodeal callus polished, virtually smooth; metapleuron shiny but with weak alutaceous sculpture. Europe…A. politus (Graham)
- -
- Antennal scape at most as long as an eye, not reaching anterior ocellus; pronotum with scattered short setae except for a row of setae near its posterior margin; propodeal callus and metapleuron distinctly alutaceous, sometimes with slightly raised reticulation…4
- 4(3)
- Scutellum with a trace of median line anteriorly (Figure 11e)…A. trilinus Cao & Zhu
- -
- Scutellum without median line…5
- 5(4)
- Antenna with each funicular longer than broad; 3–4 tarsal segments dark; costal cell 15× as long as broad. North Korea…A. leleji Kostjukov & Kosheleva
- -
- Antenna at least with F3 distinctly or slightly transverse; tarsal segments testaceous; costal cell at most 12× as long as broad…6
- 6(5)
- Eye 1.15× as long as broad; spine of clava 0.7× length of C3; pronotum 0.2× length of mesoscutum; MLM 1.45× as broad as long; propodeal spiracle separated by 0.25 its length from metanotum; tegula black; small species with body length 1.2 mm. Russia…A. sugonjaevi Kostjukov & Kosheleva
- -
- Eye 1.3× as long as broad; spine of clava 0.5× length of C3; pronotum 0.3× length of mesoscutum; MLM about 1.3× as broad as long; propodeal spiracles separated by about 0.3 its length from metanotum; tegula fuscous; body length 1.4–1.6 mm. Germany…A. evanescens (Ratzeburg)
- 7(1)
- Females…8
- -
- Males…16
- 8(7)
- Scutellum with submedian lines or with traces of submedian lines (e.g., Figure 3a)…9
- -
- Scutellum without submedian lines (e.g., Figure 9d)…12
- 9(8)
- -
- Scutellum with somewhat distinct submedian lines although incomplete; torulus situated above the lower margin of eyes; frontofacial sutures absent or indistinct, scrobal grooves shallow (e.g., Figure 3b)…11
- 10(9)
- Vertex without vertexal sutures; pronotum with 4 long setae near posterior margin; propodeum with anterior part of median carina having narrow groove or small fovea; outer rim of spiracle partially covered by the raised lobe of callus; hind coxa without dorsal carina. Europe…A. atherigonae Ferriere
- -
- Vertex with vertexal sutures; pronotum with 7–8 long setae near posterior margin; propodeum with anterior part of median carina without narrow groove or fovea; outer rim of spiracle not covered by the raised lobe of callus; hind coxa with complete dorsal carina (Figure 7b). China…A. carinata Cao & Zhu
- 11(9)
- Antenna with each funicular longer than broad; frontofacial sutures absent (Figure 17 in Ikeda [20]); propodeal callus with 3–5 setae; spiracles of penultimate gastral tergite situated on lateral sides of gaster, invisible in dorsal view, Gt5 with posterior margin straight (Figure 23 in Ikeda [20]). Japan, South Korea…A. kamijoi Ikedai
- -
- Antenna with F2 and F3 transverse (Figure 3d); frontofacial sutures short and narrowed separated from each other and divergent on the vertex (Figure 3b,c); propodeal callus with 1 seta; spiracles of penultimate gastral tergite situated on dorsolateral sides of gaster, distinct and visible in dorsal view, Gt5 with posterior margin emarginated at dorsolateral corners (Figure 4f). China…A. bilinis Huangfu & Cao
- 12(8)
- Pronotum with a row of 8–12 long setae near posterior margin…13
- -
- Pronotum with a row of 4 long setae near posterior margin…14
- 13(12)
- Pronotum with a row of 10–12 long setae near posterior margin; MLM with 3 adnotaular setae on each side; Larger species, 1.4–1.7 mm. UK…A. claridgei Graham
- -
- Pronotum with a row of 8–10 long setae near posterior margin. MLM with 2–3 adnotaular setae on each side. Smaller species, length 1.1–1.4 mm. Europe…A. granularis Domenichini
- 14(12)
- F1 and F3 longer than broad, F2 subquadrate. Forewing with costal cell 21× as long as broad. Propodeam distinctly longer than scutellum. India…A. wayanadensis Narendran and Santhosh
- -
- Antenna with each funicular transverse. Forewing with costal cell less than 14× as long as broad. Propodeam distinctly shorter than scutellum…15
- 15(14)
- Anterior adnotaular seta shorter than the posterior; tibiae infuscate with base and tip testaceous; forewing sparsely pilose all over, with costal cell more than 12× as long as broad. Czechoslovakia, USA…A. filbriata Graham
- -
- Adnotaular setae equal in length; tibiae completely testaceous (Figure 8a); forewing tending to be thickly pilose all over, with costal cell less than 10× as long as broad. Worldwide…A. indica (Silvestri)
- 16(7)
- Antennal scape with ventral plaque only about 0.4 its length, situated in the upper half…17
- -
- Antennal scape with ventral plaque 0.55–0.6× its length, extending somewhat into the lower half…18
- 17(16)
- Anterior adnotaular seta shorter than the posterior; tibiae infuscate with base and tips testaceous. Forewing sparsely pilose all over. Small species, length about 1 mm…A. filbriata Graham
- -
- 18(16)
- Scutellum with submedian lines or with traces of submedian lines…19
- -
- Scutellum without submedian lines…21
- 19(18)
- Scutellum only with traces of submedian lines; torulli situated below the lower margin of eyes; vertex without vertexal sutures; frontofacial sutures distinct, scrobal grooves deep…A. atherigonae Ferriere
- -
- Scutellum with distinct submedian lines; torulus situated above the lower margin of eyes; vertex with vertexal sutures; frontofacial sutures absent or indistinct, scrobal grooves shallow…20
- 20(19)
- Frontofacial sutures absent; propodeal callus with 3–5 setae…A. kamijoi Ikedai
- -
- 21(18)
- Pronotum with a row of 10–12 long setae near posterior margin; MLM with 3 adnotaular setae on each side; Larger species, 1.4–1.7 mm. UK…A. claridgei Graham
- -
- Pronotum with a row of 8–10 long setae near posterior margin. MLM with 2–3 adnotaular setae on each side. Smaller species, length 1.1–1.4 mm. Europe…A. granularis Domenichini
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.P.; Li, D.S.; Zhao, Y.C.; Huang, S.H.; Zhang, B.X. Aceratoneuromyia indica (Silvestri), a New Recorded Species of Parasitoid for Bactrocera dorsalis(Hendel)in China and Its Parasitic Efficiency. Chin. J. Biol. Control 2009, 25, 106–111, (In Chinese with English abstract). [Google Scholar]
- Ovruski, S.M.; Schliserman, P. Biological control of tephritid fruit flies in Argentina: Historical review, current status, and future trends for developing a parasitoid mass-release program. Insects 2012, 3, 870–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wharton, R.A.; Yoder, M.J. Parasitoids of Fruit-Infesting Tephritidae. 2018. Available online: http://paroffit.org (accessed on 19 April 2022).
- Pourhaji, A.; Lotfalizadeh, H.; Farshbaf-Pourabad, R.; Gharali, B. Parasitic wasps (Hymenoptera: Chalcidoidea) associated with flower-heads of Asteraceae in Iran. Biologia 2020, 75, 547–565. [Google Scholar] [CrossRef]
- Amini, A.; Sadeghi, H.; Lotfalizadeh, H.; Notton, D. Parasitoids (Hymenoptera: Pteromalidae, Diapriidae) of Carpomya vesuviana Costa (Diptera: Tephritidae) in South Khorasan province of Iran. Biharean Biol. 2014, 8, 122–123. [Google Scholar]
- Ovruski, S.M.; Schliserman, P.; Olga, R.; Peñaloza, C.; Oroño, L.E.; Colin, C. The establishment of Aceratoneuromyia indica (Hymenoptera: Eulophidae) in three biogeographical regions of Argentina. Fla. Entomol. 2006, 89, 270–273. [Google Scholar] [CrossRef]
- Graham, M.W.R.d.V. A reclassification of the European Tetrastichinae (Hymenoptera: Eulophidae): Revision of the remaiming genera. Mem. Amer. Ent. Inst. 1991, 49, 1–322. [Google Scholar]
- LaSalle, J. North american genera of tetrastichinae (Hymenoptera: Eulophidae). J. Nat. Hist. 1994, 28, 109–236. [Google Scholar] [CrossRef]
- Noyes, J. Universal Chalcidoidea Database. 2019. Available online: http://www.nhm.ac.uk/chalcidoids (accessed on 17 April 2022).
- Kostjukov, V.V.; Kosheleva, O.V. Taxonomic status of the species described in the genus Tetrastichus Haliday from the USSR and description of Trjapitzinichus gen. nov. In Biologicheskaya Zashchita Rasteniy—osnova stabilizatsii agrozkosistem 4; Nadykta, V.D., Ismailov, V.Y., Sugonyaev, E.S., Levaschova, G.I., Eds.; Krasnodar, Russia, 2006; pp. 103–111. Available online: https://www.nhm.ac.uk/resources/research-curation/projects/chalcidoids/pdf_X/KostjuKo2006.pdf (accessed on 17 April 2022). (In Russian)
- Kostjukov, V.V.; Kosheleva, O.V. Two new species of Trjapitzinichus Kostjukov et Kosheleva, 2006 (Hymenoptera: Eulophidae: Tetrastichinae) from the North Korea and Russia, with redescription of the genus. Евразиатский энтoмoлoгический журнал 2016, 15, 84–89. (In Russian) [Google Scholar]
- Gibson, G.A.P. Morphology and terminology. In Annotated Keys to the Genera of Nearctic Chalcidoidea (Hymenoptera); Gibson, G.A.P., Huber, J.T., Woolley, J.B., Eds.; NRC Research Press: Ottawa, ON, Canada, 1997; pp. 279–284. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Carpenter, J.M. Towards simultaneous analysis of morphological and molecular data in Hymenoptera. Zool. Scr. 1999, 28, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girault, A.A. New Australian Chaleid-flies (Hymenoptera, Chalcididae). Insecutor Inscitiae Menstruus 1917, 5, 151. [Google Scholar]
- Bouček, Z. Australasian Chalcidoidea (Hymenoptera). A Biosystematic Revision of Genera of Fourteen Families, with a Reclassification of Species; CAB International: Wallingford, UK, 1988. [Google Scholar]
- Ikeda, E. An unusual new species of Aceratoneuromyia Girault (Hymenoptera, Eulophidae), with redescription of A. therigonae Ferriѐre. Insecta Matsumurana New Ser. 1999, 55, 1–11. [Google Scholar]
- Silvestri, F. Introduzione in Italia d’un Imenottero indiano per combattere la mosca delle arance. Boll. Lab. Zool. Gen. Agr. Portici 1910, 4, 228–245. [Google Scholar]
- Kurdjumov, N.V. Notes on Tetrastichini (Hymenoptera, Chalcidoidea). Rus. Entomol. Obozr. 1913, 13, 243–256. [Google Scholar]
- Gahan, A.B. Notes on some genera and species of Chalcidoidea (Hymenoptera). Proc. Entomol. Soc. Wash. 1938, 40, 209–227. [Google Scholar]
- Gradwell, G.R. The selection of a neotype for Melittobia hawaiiensis Perkins and re-erection of the genus Aceratoneuromyia Girault (Hym., Eulophidae). Entomol. Mon. Mag. 1959, 94, 277–278. [Google Scholar]
- Sugonjaev, E.S.; Kostjukov, V.V. A note on the genus Aceratoneuromyia Girault (Hymenoptera, Eulophidae). Zool. Zhurnal 1976, 55, 1110–1112. [Google Scholar]
- Bosch, H.A.J.i.d.; Assem, J.v.d. The taxonomic position of Aceratoneuromyia granulans Domenichini (Hymenoptera: Eulophidae) as judged by characteristics of its courtship behaviour. Syst. Entomol. 1986, 11, 19–23. [Google Scholar] [CrossRef]
- Cao, H.X.; La Salle, J.; Fornoff, F.; Guo, P.F.; Zhu, C.D. Notes on Kocourekia Bouček (Hymenoptera: Eulophidae: Tetrastichinae) with description of a new species from China. Zootaxa 2017, 4317, 391–400. [Google Scholar] [CrossRef]
- Bouček, Z. Descriptions of Tachinobia gen. n. and three new species of Tetrastichinae (Hymenoptera: Eulophidae), with a tentative key to genera. Bull. Entomol. Res. 1977, 67, 17–30. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Species | Sex | Host | Primer Pair | Accession Number |
|---|---|---|---|---|---|
| CHX_ACE_833 | A. indica | female | Bactrocera dorsalis | LCO1490 & HCOout | ON260937 |
| CHX_ACE_834 | A. indica | female | Bactrocera dorsalis | LCO1490 & HCOout | ON260938 |
| CHX_ACE_835 | A. indica | female | Bactrocera dorsalis | LCO1490 & HCOout | ON260939 |
| CHX_ACE_836 | A. indica | female | Bactrocera dorsalis | LCO1490 & HCOout | ON260940 |
| CHX_ACE_837 | A. indica | female | Bactrocera dorsalis | LCO1490 & HCOout | ON260941 |
| CHX_ACE_838 | A. indica | female | Bactrocera dorsalis | LCO1490 & HCOout | ON260942 |
| CHX_ACE_839 | A. indica | male | Bactrocera dorsalis | LCO1490 & HCOout | ON260943 |
| CHX_ACE_840 | A. indica | male | Bactrocera dorsalis | LCO1490 & HCOout | ON260944 |
| CHX_ACE_841 | A. indica | male | Bactrocera dorsalis | LCO1490 & HCOout | ON260945 |
| CHX_ACE_842 | A. indica | male | Bactrocera dorsalis | LCO1490 & HCOout | ON260946 |
| CHX_ACE_843 | A. indica | male | Bactrocera dorsalis | LCO1490 & HCOout | ON260947 |
| CHX_ACE_844 | A. indica | male | Bactrocera dorsalis | LCO1490 & HCOout | ON260948 |
| CHX_ACE_788 | A. bilinis | female | Pelmatops ichneumoneus | C1-J-1514 & C1-N-2194 | ON260949 |
| CHX_ACE_789 | A. bilinis | female | Pelmatops ichneumoneus | C1-J-1514 & C1-N-2194 | ON260950 |
| CHX_ACE_790 | A. bilinis | female | Pelmatops ichneumoneus | C1-J-1514 & C1-N-2194 | ON260951 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huangfu, N.; Cao, H.-X.; Zhu, C.-D. Notes on the Genus Aceratoneuromyia Girault (Hymenoptera: Eulophidae) . Insects 2022, 13, 450. https://doi.org/10.3390/insects13050450
Huangfu N, Cao H-X, Zhu C-D. Notes on the Genus Aceratoneuromyia Girault (Hymenoptera: Eulophidae) . Insects. 2022; 13(5):450. https://doi.org/10.3390/insects13050450
Chicago/Turabian StyleHuangfu, Ning, Huan-Xi Cao, and Chao-Dong Zhu. 2022. "Notes on the Genus Aceratoneuromyia Girault (Hymenoptera: Eulophidae) " Insects 13, no. 5: 450. https://doi.org/10.3390/insects13050450
APA StyleHuangfu, N., Cao, H.-X., & Zhu, C.-D. (2022). Notes on the Genus Aceratoneuromyia Girault (Hymenoptera: Eulophidae) . Insects, 13(5), 450. https://doi.org/10.3390/insects13050450