
Fruit Fly Larval Survival in Picked and Unpicked Tomato Fruit of Differing Ripeness and Associated Gene Expression Patterns

 , , and
, , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Larval Survival
2.1.1. Insect Source
2.1.2. Tomato Fruit
2.1.3. Determination of Experimental Time Points
2.1.4. Larval Inoculation and Survival Assessment
2.1.5. Data Analysis
2.2. Gene Expression
2.2.1. Tissue Collection
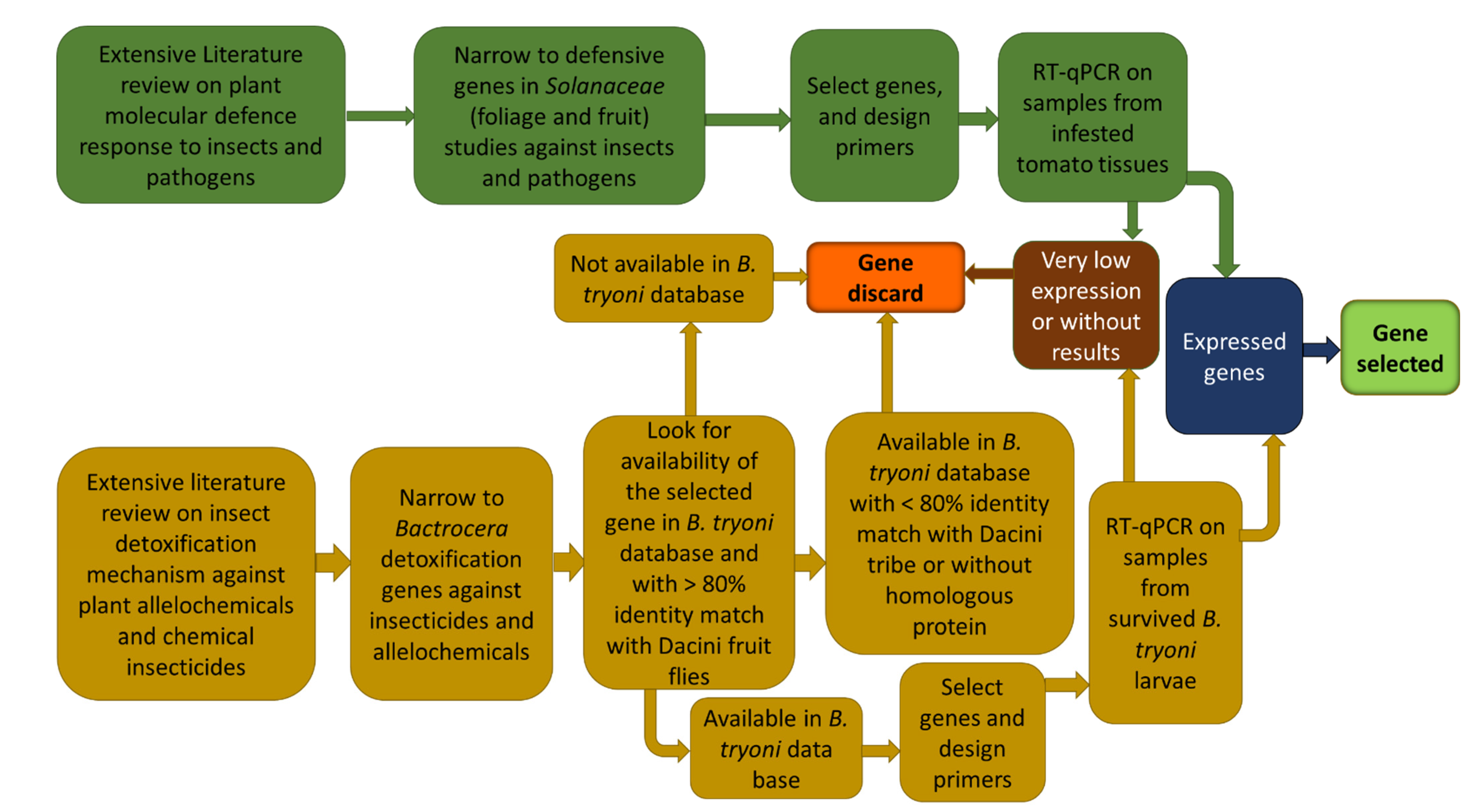
2.2.2. Nominated Genes and Primers
2.2.3. qPCR Conditions
2.2.4. Data Analysis
3. Results
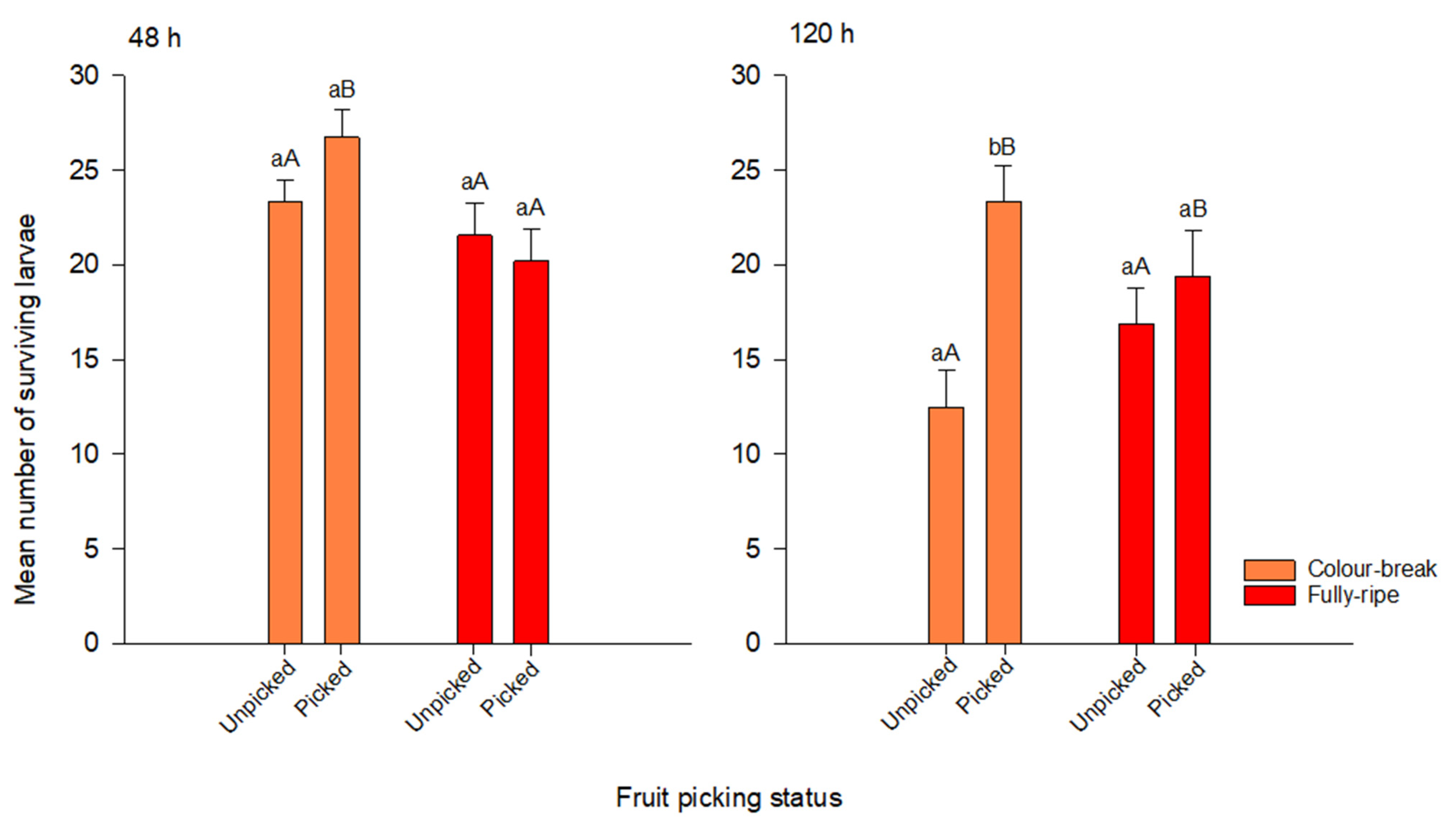
3.1. Larval Survival
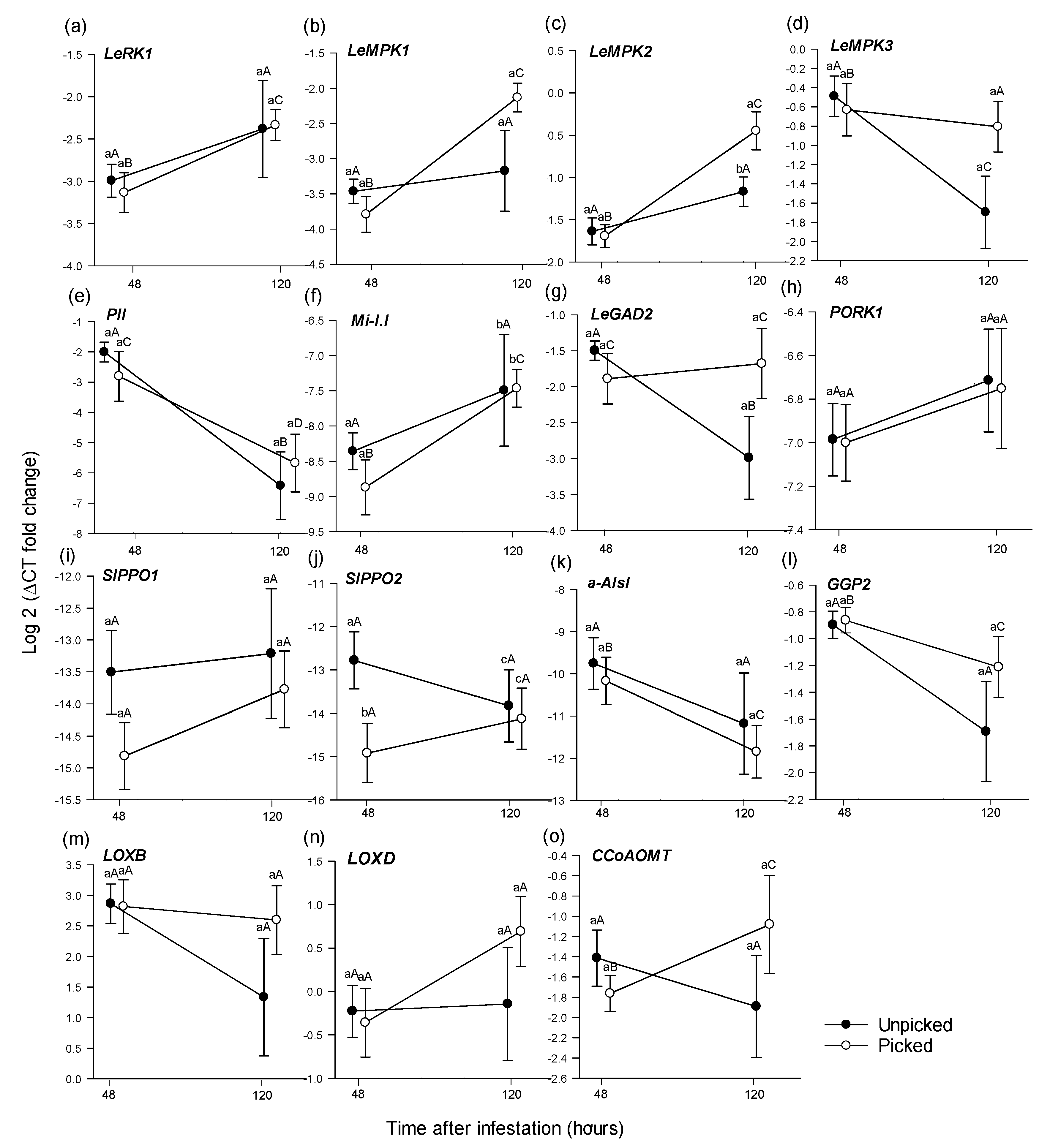
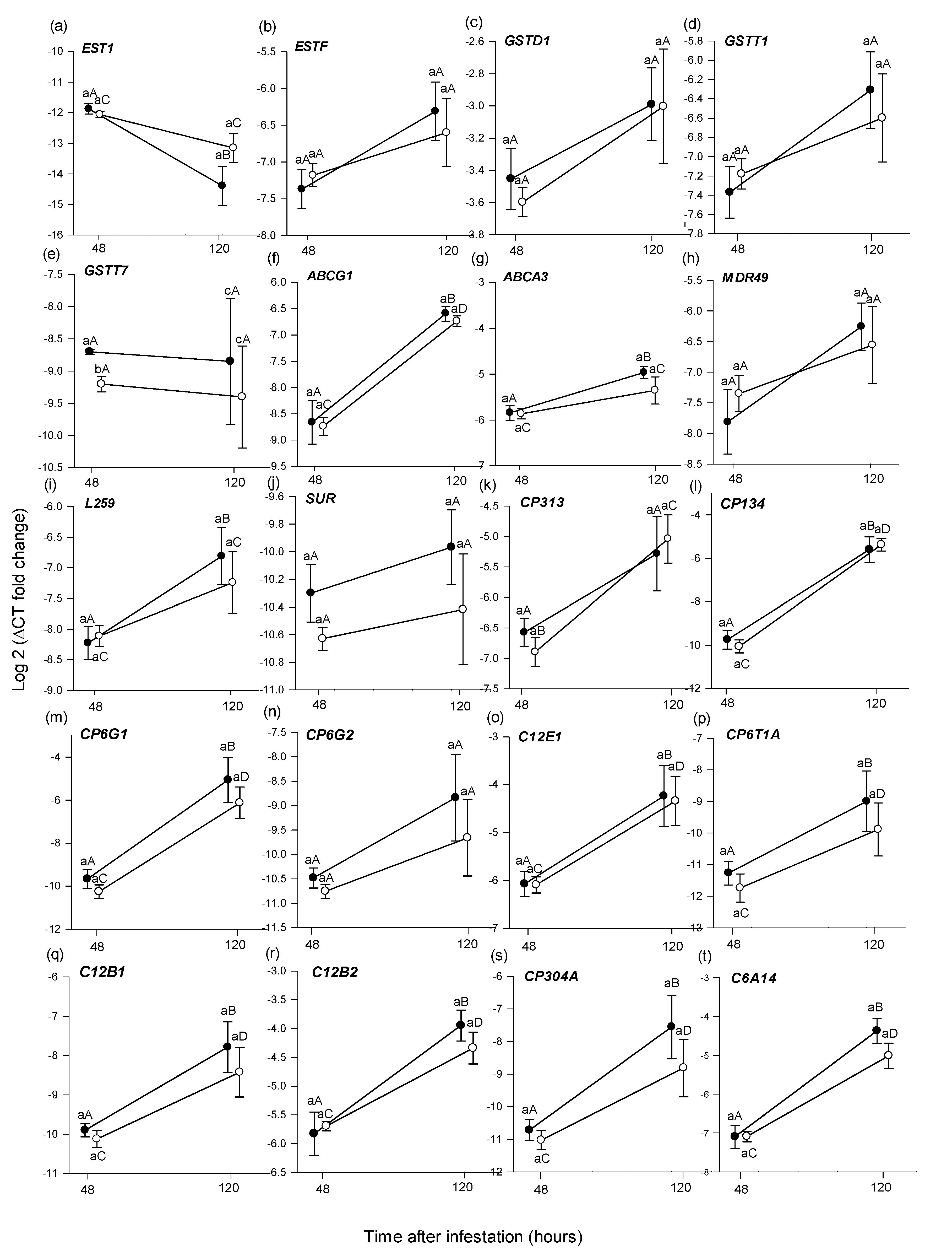
3.2. Comparative Gene Expression
3.2.1. Differential Gene Expression in Picked and Unpicked Roma Tomato Fruit
Within a Sampling Period across Picking States
Across Sampling Periods within a Picking State
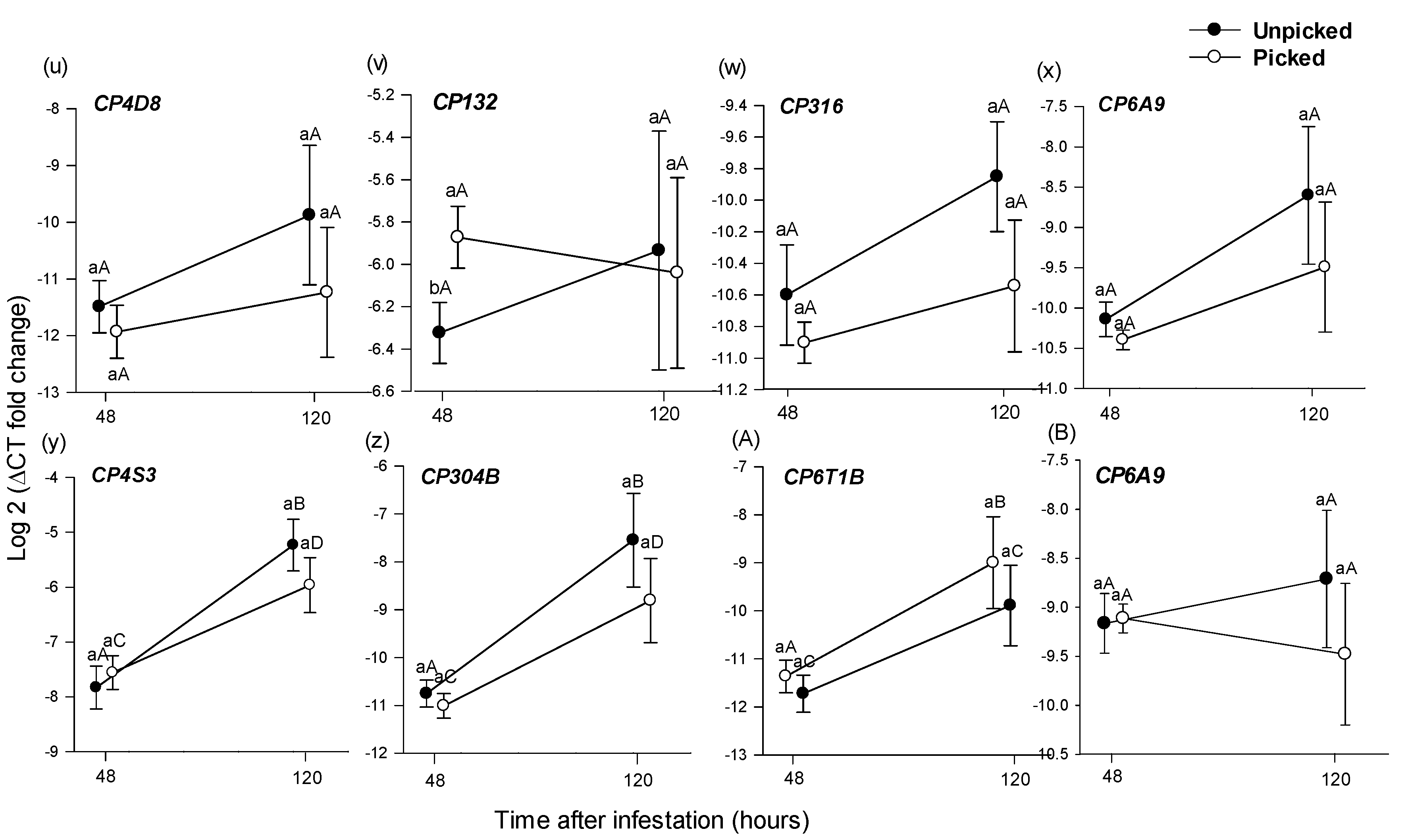
3.2.2. Differential Gene Expression in B. tryoni Larval Tissue
Within a Sampling Period across Picking States
Across Sampling Periods within a Picking State
4. Discussion
4.1. Results Summary
4.2. B. tryoni Larval Survival in Tomato Fruit of Varying Ripeness and Harvest Status
4.3. Larval Gene Expression Indicates a Detoxification Response in Both Picked and Unpicked Fruit
4.4. Tomato Gene Expression Patterns
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McCall, A.C.; Fordyce, J.A. Can optimal defence theory be used to predict the distribution of plant chemical defences? J. Ecol. 2010, 98, 985–992. [Google Scholar] [CrossRef]
- Rhoades, D.F.; Cates, R.G. Toward a general theory of plant antiherbivore chemistry. In Biochemical Interaction Between Plants and Insects; Wallace, J.W., Mansell, R.L., Eds.; Recent Advances in Phytochemistry; Springer: Boston, MA, USA, 1976; pp. 168–213. [Google Scholar] [CrossRef]
- Van der Pijl, L. Principles of Dispersal, 3rd ed.; Springer: Berlin, Germany, 1982. [Google Scholar] [CrossRef]
- Zangerl, A.R.; Rutledge, C.E. The probability of attack and patterns of constitutive and induced defense: A test of optimal defense theory. Am. Nat. 1996, 147, 599–608. [Google Scholar] [CrossRef]
- Cazetta, E.; Schaefer, H.M.; Galetti, M. Does attraction to frugivores or defense against pathogens shape fruit pulp composition status? Oecologia 2008, 155, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Cipollini, M.L.; Levey, D.J. Why are some fruits toxic? glycoalkaloids in solanum and fruit choice by vertebrates. Ecology 1997, 78, 782–798. [Google Scholar] [CrossRef]
- Salerno, G.; Rebora, M.; Piersanti, A.; Gorb, E.; Gorb, S. Mechanical ecology of fruit-insect interaction in the adult Mediterranean fruit fly Ceratitis capitata (Diptera: Tephritidae). Zoology 2020, 139, 125748. [Google Scholar] [CrossRef] [PubMed]
- Nevo, O.; Razafimandimby, D.; Jeffrey, J.A.J.; Schulz, S.; Ayasse, M. Fruit scent as an evolved signal to primate seed dispersal. Sci. Adv. 2018, 4, eaat4871. [Google Scholar] [CrossRef]
- Nevo, O.; Razafimandimby, D.; Valenta, K.; Jeffrey, J.A.J.; Reisdorff, C.; Chapman, C.A.; Ganzhorn, J.U.; Ayasse, M. Signal and reward in wild fleshy fruits: Does fruit scent predict nutrient content? Ecol. Evol. 2019, 9, 10534–10543. [Google Scholar] [CrossRef]
- Whitehead, S.R.; Quesada, M.F.O.; Bowers, M.D. Chem tradeoffs in seed dispersal: Defensive metabolites in fruits deter consumption by mutualist bats. Oikos 2016, 125, 927–937. [Google Scholar] [CrossRef]
- Agrawal, A.A.; Gorski, P.M.; Tallamy, D.W. Polymorphism in plant defense against herbivory: Constitutive and induced resistance in Cucumis sativus. J. Chem. Ecol. 1999, 25, 2285–2304. [Google Scholar] [CrossRef]
- Farmer, E.E. Leaf Defence; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Paudel, S.; Lin, P.-A.; Foolad, M.R.; Ali, J.G.; Rajotte, E.G.; Felton, G.W. Induced plant defenses against herbivory in cultivated and wild tomato. J. Chem. Ecol. 2019, 45, 693–707. [Google Scholar] [CrossRef]
- War, A.R.; Taggar, G.K.; Hussain, B.; Taggar, M.S.; Nair, R.M.; Sharma, H.C. Plant defence against herbivory and insect adaptations. AoB Plants 2018, 10, ply037. [Google Scholar] [CrossRef]
- Diezel, C.; Allmann, S.; Baldwin, I.T. Mechanisms of optimal defense patterns in Nicotiana attenuata: Flowering attenuates herbivory-elicited ethylene and jasmonate signalling. J. Integr. Plant Biol. 2011, 53, 971–983. [Google Scholar] [CrossRef] [PubMed]
- Schiestl, F.P.; Kirk, H.; Bigler, L.; Cozzolino, S.; Desurmont, G.A. Herbivory and floral signaling: Phenotypic plasticity and tradeoffs between reproduction and indirect defense. New Phytol. 2014, 203, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Gogi, M.D.; Ashfaq, M.; Arif, M.J.; Sarfraz, R.M.; Nawab, N.N. Investigating phenotypic structures and allelochemical compounds of the fruits of Momordica charantia L genotypes as sources of resistance against Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae). Crop Prot. 2010, 29, 884–890. [Google Scholar] [CrossRef]
- Staub, C.G.; De Lima, F.; Majer, J.D. Determination of host status of citrus fruits against the Mediterranean fruit fly, Ceratitis capitata (Wiedemann) (Diptera: Tephritidae). Aust. J. Entomol. 2008, 47, 184–187. [Google Scholar] [CrossRef]
- Cipollini, M.L. Secondary metabolites of vertebrate-dispersed fruits: Evidence for adaptive functions. Rev. Chil. Hist. Nat. 2000, 73, 421–440. [Google Scholar] [CrossRef]
- Herrera, C.M. Defense of ripe fruit from pests: Its significance in relation to plant-disperser interactions. Am. Nat. 1982, 120, 218–241. [Google Scholar] [CrossRef]
- Papanastasiou, S.A.; Bali, E.-M.D.; Ioannou, C.S.; Papachristos, D.P.; Zarpas, K.D.; Papadopoulos, N.T. Toxic and hormetic-like effects of three components of citrus essential oils on adult Mediterranean fruit flies (Ceratitis capitata). PLoS ONE 2017, 12, e0177837. [Google Scholar] [CrossRef]
- Salvatore, A.; Borkosky, A.; Willink, E.; Bardón, A. Toxic effects of lemon peel constituents on Ceratitis capitata. J. Chem. Ecol. 2004, 30, 323–333. [Google Scholar] [CrossRef]
- Davis, T.S.; Garczynski, S.F.; Stevens-Rumann, C.; Landolt, P.J. A test of fruit varieties on entry rate and development by neonate larvae of the codling moth, Cydia pomonella. Entomol. Exp. Appl. 2013, 148, 259–266. [Google Scholar] [CrossRef]
- Greany, P.; Styer, S.; Davis, P.; Shaw, P.; Chambers, D. Biochemical resistance of citrus to fruit flies Demonstration and elucidation of resistance to the Caribbean fruit fly, Anastrepha suspensa. Entomol. Exp. Appl. 1983, 34, 40–50. [Google Scholar] [CrossRef]
- Muthuthantri, S.; Clarke, A.R.; Hayes, R.A.; Kevin, J. Effect of citrus peel chemicals on Bactrocera tryoni larval survival. Acta Hortic. 2015, 1105, 349–356. [Google Scholar] [CrossRef]
- Papachristos, D.P.; Kimbaris, A.C.; Papadopoulos, N.T.; Polissiou, M.G. Toxicity of citrus essential oils against Ceratitis capitata (Diptera: Tephritidae) larvae. Ann. Appl. Biol. 2009, 155, 381–389. [Google Scholar] [CrossRef]
- Kreuger, B.; Potter, D.A. Changes in saponins and tannins in ripening holly fruits and effects of fruit consumption on nonadopted insect herbivores. Am. Midl. Nat. 1994, 132, 183–191. [Google Scholar] [CrossRef]
- Lasa, R.; Tadeo, E.; Dinorín, L.A.; Lima, I.; Williams, T. Fruit firmness, superficial damage, and location modulate infestation by Drosophila suzukii and Zaprionus indianus: The case of guava in Veracruz, Mexico. Entomol. Exp. Appl. 2017, 162, 4–12. [Google Scholar] [CrossRef]
- Mulatu, B.; Applebaum, S.; Kerem, Z.; Coll, M. Tomato fruit size, maturity and α-tomatine content influence the performance of larvae of potato tuber moth Phthorimaea operculella (Lepidoptera: Gelechiidae). Bull. Entomol. Res. 2006, 96, 173–178. [Google Scholar] [CrossRef]
- Rashmi, M.A.; Verghese, A.; Shivashankar, S.; Chakravarthy, A.K.; Sumathi, M.; Kandakoors, S. Does change in tannin content in mango (Mangifera indica) fruits influence the extent of fruit fly (Bactrocera dorsalis Hendel) herbivory? J. Entomol. Zool. Stud. 2017, 5, 381–385. [Google Scholar]
- Schaefer, H.M.; Schmidt, V.; Winkler, H. Testing the defence trade-off hypothesis: How contents of nutrients and secondary compounds affect fruit removal. Oikos 2003, 102, 318–328. [Google Scholar] [CrossRef]
- Baba, V.Y.; Constantino, L.V.; Ivamoto, S.T.; Moreira, A.F.P.; Madeira, T.B.; Nixdorf, S.L.; Rodrigues, R.; Gonçalves, L.S.A. Capsicum-Colletotrichum interaction: Identification of resistance sources and quantification of secondary metabolites in unripe and ripe fruits in response to anthracnose infection. Sci. Hortic. 2019, 246, 469–477. [Google Scholar] [CrossRef]
- Cota, I.; Troncoso-Rojas, R.; Sotelo-Mundo, R.; Sánchez-Estrada, A.; Tiznado-Hernández, M. Chitinase and β-1, 3-glucanase enzymatic activities in response to infection by Alternaria alternata evaluated in two stages of development in different tomato fruit varieties. Sci. Hortic. 2007, 112, 42–50. [Google Scholar] [CrossRef]
- González, G.; Fuentes, L.; Moya-León, M.A.; Sandoval, C.; Herrera, R. Characterization of two PR genes from Fragaria chiloensis in response to Botrytis cinerea infection: A comparison with Fragaria × ananassa. Physiol. Mol. Plant Pathol. 2013, 82, 73–80. [Google Scholar] [CrossRef]
- Mikulic-Petkovsek, M.; Schmitzer, V.; Jakopic, J.; Cunja, V.; Veberic, R.; Munda, A.; Stampar, F. Phenolic compounds as defence response of pepper fruits to Colletotrichum coccodes. Physiol. Mol. Plant Pathol. 2013, 84, 138–145. [Google Scholar] [CrossRef]
- Mongkolporn, O.; Montri, P.; Supakaew, T.; Taylor, P.W. Differential reactions on mature green and ripe chili fruit infected by three Colletotrichum spp. Plant Dis. 2010, 94, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Nandineni, M.R. Genome sequencing and comparative genomics reveal a repertoire of putative pathogenicity genes in chilli anthracnose fungus Colletotrichum truncatum. PLoS ONE 2017, 12, e0183567. [Google Scholar] [CrossRef] [PubMed]
- Salzman, R.A.; Tikhonova, I.; Bordelon, B.P.; Hasegawa, P.M.; Bressan, R.A. Coordinate accumulation of antifungal proteins and hexoses constitutes a developmentally controlled defense response during fruit ripening in grape. Plant Physiol. 1998, 117, 465–472. [Google Scholar] [CrossRef]
- Shah, P.; Powell, A.L.; Orlando, R.; Bergmann, C.; Gutierrez-Sanchez, G. Proteomic analysis of ripening tomato fruit infected by Botrytis cinerea. J. Proteome Res. 2012, 11, 2178–2192. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.; Van Strien, E. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar] [CrossRef]
- Vilanova, L.; Wisniewski, M.; Norelli, J.; Viñas, I.; Torres, R.; Usall, J.; Phillips, J.; Droby, S.; Teixidó, N. Transcriptomic profiling of apple in response to inoculation with a pathogen (Penicillium expansum) and a non-pathogen (Penicillium digitatum). Plant Mol. Biol. Rep. 2014, 32, 566–583. [Google Scholar] [CrossRef]
- Wojciechowska, E.; Weinert, C.H.; Egert, B.; Trierweiler, B.; Schmidt-Heydt, M.; Horneburg, B.; Graeff-Hönninger, S.; Kulling, S.K.; Geisen, R. Chlorogenic acid, a metabolite identified by untargeted metabolome analysis in resistant tomatoes, inhibits the colonization by Alternaria alternata by inhibiting alternariol biosynthesis. Eur. J. Plant Pathol. 2014, 139, 735–747. [Google Scholar] [CrossRef]
- Anand, T.; Bhaskaran, R.; Raguchander, T.; Samiyappan, R.; Prakasam, V.; Gopalakrishnan, C. Defence responses of chilli fruits to Colletotrichum capsici and Alternaria alternata. Biol. Plant. 2009, 53, 553–559. [Google Scholar] [CrossRef]
- Ruelas, C.; Tiznado-Hernández, M.; Sánchez-Estrada, A.; Robles-Burgueno, M.; Troncoso-Rojas, R. Changes in phenolic acid content during Alternaria alternata infection in tomato fruit. J. Phytopathol. 2006, 154, 236–244. [Google Scholar] [CrossRef]
- Alkan, N.; Friedlander, G.; Ment, D.; Prusky, D.; Fluhr, R. Simultaneous transcriptome analysis of Colletotrichum gloeosporioides and tomato fruit pathosystem reveals novel fungal pathogenicity and fruit defense strategies. New Phytol. 2015, 205, 801–815. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Ulate, B.; Vincenti, E.; Powell, A.; Cantu, D. Tomato transcriptome and mutant analyses suggest a role for plant stress hormones in the interaction between fruit and Botrytis cinerea. Front. Plant Sci. 2013, 4, 142. [Google Scholar] [CrossRef] [PubMed]
- Corrado, G.; Alagna, F.; Rocco, M.; Renzone, G.; Varricchio, P.; Coppola, V.; Coppola, M.; Garonna, A.; Baldoni, L.; Scaloni, A.; et al. Molecular interactions between the olive and the fruit fly Bactrocera oleae. BMC Plant Biol. 2012, 12, 86. [Google Scholar] [CrossRef]
- Christenson, L.; Foote, R.H. Biology of fruit flies. Annu. Rev. Entomol. 1960, 5, 171–192. [Google Scholar] [CrossRef]
- Clarke, A.R. Biology and Management of Bactrocera and Related Fruit Flies; CAB International: Wallingford, UK, 2019. [Google Scholar]
- Clarke, A.R.; Armstrong, K.F.; Carmichael, A.E.; Milne, J.R.; Raghu, S.; Roderick, G.K.; Yeates, D.K. Invasive phytophagous pests arising through a recent tropical evolutionary radiation: The Bactrocera dorsalis complex of fruit flies. Annu. Rev. Entomol. 2005, 50, 293–319. [Google Scholar] [CrossRef]
- Jessup, A.; Dominiak, B.; Woods, B.; De Lima, C.; Tomkins, A.; Smallridge, C. Area-wide management of fruit flies in Australia. In Area-Wide Control of Insect Pests; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 685–697. [Google Scholar] [CrossRef]
- Mau, R.F.L.; Jang, E.B.; Vargas, R.I. The Hawaii area-wide fruit fly pest management programme: Influence of partnerships and a good education programme. In Area-Wide Control of Insect Pests; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 671–683. [Google Scholar] [CrossRef]
- Hawkes, N.J.; Janes, R.W.; Hemingway, J.; Vontas, J. Detection of resistance-associated point mutations of organophosphate-insensitive acetylcholinesterase in the olive fruit fly, Bactrocera oleae (Gmelin). Pestic Biochem. Physiol. 2005, 81, 154–163. [Google Scholar] [CrossRef]
- Jin, T.; Zeng, L.; Lin, Y.; Lu, Y.; Liang, G. Insecticide resistance of the oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae), in mainland China. Pest Manag. Sci. 2011, 67, 370–376. [Google Scholar] [CrossRef]
- Vontas, J.; Hernández-Crespo, P.; Margaritopoulos, J.T.; Ortego, F.; Feng, H.T.; Mathiopoulos, K.D.; Hsu, J.C. Insecticide resistance in tephritid flies. Pestic Biochem. Physiol. 2011, 100, 199–205. [Google Scholar] [CrossRef]
- Dominiak, B.C. Components of a systems approach for the management of Queensland fruit fly Bactrocera tryoni (Froggatt) in a post dimethoate fenthion era. J. Crop Prot. 2019, 116, 56–67. [Google Scholar] [CrossRef]
- Aluja, M.; Arredondo, J.; Díaz-Fleischer, F.; Birke, A.; Rull, J.; Niogret, J.; Epsky, N. Susceptibility of 15 mango (Sapindales: Anacardiaceae) cultivars to the attack by Anastrepha ludens and Anastrepha obliqua (Diptera: Tephritidae) and the role of underdeveloped fruit as pest reservoirs: Management implications. J. Econ. Entomol. 2014, 107, 375–388. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Muthuthantri, S.; Clarke, A.R. Five commercial citrus rate poorly as hosts of the polyphagous fruit fly Bactrocera tryoni (Froggatt) (Diptera: Tephritidae) in laboratory studies. Aust. J. Entomol. 2012, 51, 289–298. [Google Scholar] [CrossRef]
- Papachristos, D.P.; Papadopoulos, N.T.; Nanos, G.D. Survival and development of immature stages of the Mediterranean fruit fly (Diptera: Tephritidae) in citrus fruit. J. Econ. Entomol. 2008, 101, 866–872. [Google Scholar] [CrossRef]
- Rattanapun, W.; Amornsak, W.; Clarke, A.R. Bactrocera dorsalis preference for and performance on two mango varieties at three stages of ripeness. Entomol. Exp. Appl. 2009, 131, 243–253. [Google Scholar] [CrossRef]
- Mateos, M.; Martinez Montoya, H.; Lanzavecchia, S.B.; Conte, C.; Guillén, K.; Morán-Aceves, B.M.; Toledo, J.; Liedo, P.; Asimakis, E.D.; Doudoumis, V.; et al. Wolbachia pipientis associated with tephritid fruit fly pests: From basic research to applications. Front Microbiol 2020, 11, 1080. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, J.W.; Mitchell, W.C.; Farias, G.J. Resistance of ‘Sharwil’ avocados at harvest maturity to infestation by three fruit fly species (Diptera: Tephritidae) in Hawaii. J. Econ. Entomol. 1983, 76, 119–121. [Google Scholar] [CrossRef]
- Liquido, N.J.; Chan, H.T., Jr.; McQuate, G.T. Hawaiian tephritid fruit flies (Diptera): Integrity of the infestation-free quarantine procedure for ‘Sharwil’avocado. J. Econ. Entomol. 1995, 88, 85–96. [Google Scholar] [CrossRef]
- Papadopoulos, N.T.; Papachristos, D.P.; Ioannou, C. Citrus fruits and the Mediterranean fruit fly. Acta Hortic. 2015, 1065, 1009–1018. [Google Scholar] [CrossRef]
- Nehra, S.; Singh, S.; Samota, R.G.; Choudhary, S.K.; Choudhary, A.L. Screening of round gourd varieties for resistance against fruit fly, Bactrocera cucurbitae (Coquillett). J. Pharmacogn. Phytochem. 2019, 8, 1101–1107. [Google Scholar]
- Bower, C.C. Inhibition of larval growth of the Queensland fruit fly, Dacus tryoni (Diptera: Tephritidae) in apples. Ann. Entomol. Soc. Am. 1977, 70, 97–100. [Google Scholar] [CrossRef]
- Reissig, W. Survival of apple maggot larvae, Rhagoletis pomonella (Diptera: Tephritidae), in picked and unpicked apples. Can. Entomol. 1979, 111, 181–187. [Google Scholar] [CrossRef]
- Balagawi, S.; Vijaysegaran, S.; Drew, R.A.; Raghu, S.J. Influence of fruit traits on oviposition preference and offspring performance of Bactrocera tryoni (Froggatt) (Diptera: Tephritidae) on three tomato (Lycopersicon lycopersicum) cultivars. Aust. J. Entomol. 2005, 44, 97–103. [Google Scholar] [CrossRef]
- Roohigohar, S.; Prentis, P.J.; Clarke, A.R. Effect of tomato fruit cultivar and ripening stage on Bactrocera tryoni (Froggatt) egg and larval survival. J. Appl. Entomol. 2020, 144, 797–805. [Google Scholar] [CrossRef]
- Roohigohar, S.; Clarke, A.R.; Prentis, P.J. Gene selection for studying frugivore-plant interactions: A review and an example using Queensland fruit fly in tomato. PeerJ 2021, 9, e11762. [Google Scholar] [CrossRef]
- Heather, N.W.; Corcoran, R.J. Dacus tryoni. In Handbook of Insect Rearing; Singh, P., Moore, R.F., Eds.; Elsevier: Amsterdam, The Netherlands, 1985; pp. 41–48. [Google Scholar]
- Sargent, S.A. Tomato Production Guide for Florida: Harvest and Handling; Florida Cooperative Extension Service, Institute of Food and Agricultural Sci, University of Florida: Gainesville, FL, USA, 1997. [Google Scholar]
- Leach, P. Phytosanitary measures. In Biology and Management of Bactrocera and Related Fruit Flies; Clarke, A.R., Ed.; CAB International: Wallingford, UK, 2019; pp. 195–225. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research, 4th ed.; WH Freeman and Company: New York, NY, USA, 2012. [Google Scholar]
- Graham, M.H.; Edwards, M.S. Statistical significance versus fit: Estimating the importance of individual factors in ecological analysis of variance. Oikos 2001, 93, 505–513. [Google Scholar] [CrossRef]
- Pfaffl, M.W. Relative quantification. In Real-Time PCR; Dorak, M.T., Ed.; Taylor and Francis: New York, NY, USA, 2007; pp. 89–108. [Google Scholar]
- Ganger, M.T.; Dietz, G.D.; Ewing, S.J. A common base method for analysis of qPCR data and the application of simple blocking in qPCR experiments. BMC Bioinform. 2017, 18, 1–11. [Google Scholar] [CrossRef]
- Orcan, F. Parametric or non-parametric: Skewness to test normality for mean comparison. Int. J. Assess. Tool Educ. 2020, 7, 255–265. [Google Scholar] [CrossRef]
- Arnó, J.; Solà, M.; Riudavets, J.; Gabarra, R. Population dynamics, non-crop hosts, and fruit susceptibility of Drosophila suzukii in Northeast Spain. J. Pest Sci. 2016, 89, 713–723. [Google Scholar] [CrossRef]
- Ben-Yosef, M.; Pasternak, Z.; Jurkevitch, E.; Yuval, B. Symbiotic bacteria enable olive fly larvae to overcome host defences. R. Soc. Open Sci. 2015, 2, 150170. [Google Scholar] [CrossRef]
- Díaz-Fleischer, F.; Aluja, M. Clutch size in frugivorous insects as a function of host firmness: The case of the tephritid fly Anastrepha ludens. Ecol. Entomol. 2003, 28, 268–277. [Google Scholar] [CrossRef]
- Joachim-Baravo, I.S.; Fernandes, O.A.; De Bortoli, S.A.; Zucoloto, F.S. Oviposition behavior of Ceratitis capitata Wiedemann (Diptera: Tephritidae): Association between oviposition preference and larval performance in individual females. Neotrop. Entomol. 2001, 30, 559–564. [Google Scholar] [CrossRef]
- Ishiguri, Y.; Toyoshima, S. Larval survival and development of the peach fruit moth, Carposina sasakii (Lepidoptera: Carposinidae), in picked and unpicked apple fruits. Appl. Entomol. Zool. 2006, 41, 685–690. [Google Scholar] [CrossRef]
- Alkan, N.; Fortes, A.M. Insights into molecular and metabolic events associated with fruit response to post-harvest fungal pathogens. Front. Plant Sci. 2015, 6, 889. [Google Scholar] [CrossRef] [PubMed]
- Beno-Moualem, D.; Prusky, D. Early events during quiescent infection development by Colletotrichum gloeosporioides in unripe avocado fruits. Phytopathology 2000, 90, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Birnbaum, S.S.; Rinker, D.C.; Gerardo, N.M.; Abbot, P. Transcriptional profile and differential fitness in a specialist milkweed insect across host plants varying in toxicity. Mol. Ecol. 2017, 26, 6742–6761. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Lv, S.; Zhang, Z.; Chang, B.H. Phenotypic and transcriptomic response of the grasshopper Oedaleus asiaticus (Orthoptera: Acrididae) to toxic rutin. Front. Physiol. 2020, 11, 52. [Google Scholar] [CrossRef] [PubMed]
- Misra, J.R.; Horner, M.A.; Lam, G.; Thummel, C.S. Transcriptional regulation of xenobiotic detoxification in Drosophila. Genes Dev. 2011, 25, 1796–1806. [Google Scholar] [CrossRef] [PubMed]
- Scanlan, J.L.; Gledhill-Smith, R.S.; Battlay, P.; Robin, C. Genomic and transcriptomic analyses in Drosophila suggest that the ecdysteroid kinase-like (EcKL) gene family encodes the ‘detoxification-by-phosphorylation’ enzymes of insects. Insect Biochem. Mol. Biol. 2020, 123, 103429. [Google Scholar] [CrossRef]
- Xu, C.; Li, C.Y.T.; Kong, A.N.T. Induction of phase I, II and III drug metabolism/transport by xenobiotics. Arch. Pharm. Res. 2005, 28, 249–268. [Google Scholar] [CrossRef]
- Després, L.; David, J.P.; Gallet, C. The evolutionary ecology of insect resistance to plant chemicals. Trends Ecol. Evol. 2007, 22, 298–307. [Google Scholar] [CrossRef]
- Hoang, K.; Matzkin, L.M.; Bono, J.M. Transcriptional variation associated with cactus host plant adaptation in Drosophila mettleri populations. Mol. Ecol. 2015, 24, 5186–5199. [Google Scholar] [CrossRef] [PubMed]
- Pavlidi, N.; Gioti, A.; Wybouw, N.; Dermauw, W.; Ben-Yousef, M.; Tuval, B.; Jurkevich, E.; Kampouraki, A.; Van Leeuwen, T.; Vontas, J. Transcriptomic responses of the olive fruit fly Bactrocera oleae and its symbiont Candidatus erwinia dacicola to olive feeding. Sci. Rep. 2017, 7, 42633. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.H.; Acevedo, T.; Harris, E.V.; Alcaide, T.Y.; Walters, J.R.; Hunter, M.D.; Gerardo, N.M.; de Roode, J.C. Transcriptomics of monarch butterflies (Danaus plexippus) reveals that toxic host plants alter expression of detoxification genes and down-regulate a small number of immune genes. Mol. Ecol. 2019, 28, 4845–4863. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M. Tomato glycoalkaloids: Role in the plant and in the diet. J. Agric. Food Chem. 2002, 50, 5751–5780. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.T.; Tam, S.Y.T. Toxicity of α-tomatine to larvae of the Mediterranean fruit fly (Diptera: Tephritidae). J. Econ. Entomol. 1985, 78, 305–307. [Google Scholar] [CrossRef]
- Baxter, I. We aren’t good at picking candidate genes, and it’s slowing us down. Curr. Opin. Plant Biol. 2020, 54, 57–60. [Google Scholar] [CrossRef]
- Kandoth, P.K.; Ranf, S.; Pancholi, S.S.; Jayanty, S.; Walla, M.D.; Miller, W.; Howe, G.A.; Lincoln, D.E.; Stratmann, J.W. Tomato MAPKs LeMPK1, LeMPK2, and LeMPK3 function in the systemin-mediated defense response against herbivorous insects. Proc. Natl. Acad. Sci. USA 2007, 104, 12205–12210. [Google Scholar] [CrossRef]
- Mahanil, S.; Attajarusit, J.; Stout, M.J.; Thipyapong, P. Overexpression of tomato polyphenol oxidase increases resistance to common cutworm. Plant Sci. 2008, 174, 456–466. [Google Scholar] [CrossRef]
- Xu, S.; Liao, C.-J.; Jaiswal, N.; Lee, S.; Yun, D.-J.; Lee, S.Y.; Garvey, M.; Kaplan, I.; Mengiste, T. Tomato PEPR1 ORTHOLOG RECEPTOR-LIKE KINASE1 regulates responses to systemin, necrotrophic fungi, and insect herbivory. Plant Cell 2018, 30, 2214–2229. [Google Scholar] [CrossRef]
- Zheng, Y.; Yang, Y.; Liu, C.; Chen, L.; Sheng, J.; Shen, L. Inhibition of SlMPK1, SlMPK2, and SlMPK3 disrupts defense signaling pathways and enhances tomato fruit susceptibility to Botrytis cinerea. J. Agric. Food Chem. 2015, 63, 5509–5517. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Family/Pathway | Gene Symbol | Gene Function |
|---|---|---|
| B. tryoni detoxification pathway genes | ||
| Cytochrome P450 | CP6A9, CP313, CP134, CP4D8, CP6G1, C12E1, CP6T1A, CP6T1B, C12C1, C12B1, C12B2, CP304A, C304B, CP306, C6A14, C4AC2, CP4S3, CP132, CP316, CP6G2 | Catalysis of oxidative reactions during endogenous and exogenous metabolism and metabolism of xenobiotics and plant allelochemicals |
| Carboxylesterase | EST F, EST 1 | Hydrolysis drugs, environmental toxicants, and insecticides |
| Glutathione S-transferase | GST D1, GST T1, GST T7 | Detoxification of endogenous and xenobiotic compounds |
| ATP-binding cassette (ABC) transporters | ABCG1, ABCA3, SUR, L259, MDR49 | Facilitate cellular excretion of insecticides or metabolites |
| S. lycopersicumdefensive pathway genes | ||
| Receptor-like kinase | PORK1, LecRK1 | Phytophagous arthropod attacks perception in plant tissue |
| D-mannose/L-galactose | GGP2 | Oxidative stress response in plant against abiotic and biotic stresses |
| Mitogen-activated protein kinase | LeMPK1, LeMPK2, LeMPK3 | Plant signal transduction in response to biotic and abiotic stresses |
| Lipoxygenase | LOXB, LOXD | Plant defence response against pathogens and herbivores |
| Gamma-aminobutyric Acid | LeGAD2 | Increases plant resistance to insect herbivory |
| Polyphenol oxidase | SlPPO1, SlPPO2 | Plant defence response against pathogens and insects |
| Proteinase inhibitor | PII, a-AIs1 | Inhibiting insects’ digestive enzymes |
| Caffeoyl-CoAO-methyltransferase | CCoAOMT | Plant phytoalexins against herbivores and pathogens |
| Resistance (R) gene | Mi-1.1 | Plant resistance against pests |
| Gene Symbol | Forward Sequence 5′-3′ | Reverse Sequence 5′-3′ |
|---|---|---|
| B. tryoni detoxification pathways genes and primers | ||
| GST D1 | GCCGATTTCACCACGTATGC | GCGTGTATCGCTGAAACGTC |
| GST T1 | TTAGCACCATAGACGTGGCG | TGG GCAATACTGCGGAACTT |
| GST T7 | TGGCCGGTGATCAGTTGAAA | GCTGATCGACCATAGCACGA |
| EST F | AGCTAAACCTTCCACCACGG | CACCCATTGCAAAGCCAGAC |
| EST 1 | CGCTGTTTACGCATTCCTCG | AGCGGACGCATACTCATAGC |
| SUR | TTGCTCAAGGCAAAGCGAAC | CATCGTCATCCGTCTGCTCA |
| ABCG1 | TTCTTTGTCGGTGCTACGCT | ATGGGCGTTCCAAGCCATAA |
| ABCA3 | GGGAATAGCGATTGCGGGTA | CGCTTCTTCCATGTGATGCG |
| L259 | CAGGAGCCAGCACGTAAAGA | GGTCCAATGACGGCCACTAA |
| MDR49 | TGAGGCAACCTCGGCTTTAG | CCGAGCGCATAAGTTCAACG |
| CP6A9 | GTATCGCTTGCAACTCGCTG | CGCACGATGCGCATAAAGAA |
| CP313 | AACACTTCAAACCGGAGGCA | CTCCAGCTGACACAACGGAT |
| CP134 | AGGGCATTTCGATTGGCAGA | TCACCCGCATCGTTTCGTTA |
| CP4D8 | ATTTACTCGCACGCCATCCA | CGGCACACTGGGATAGAGAC |
| CP6G1 | TGGACGAAGTGTTGCGCTTA | GGATCGAAAGTGTCCGGGTT |
| C12E1 | ATGTGGACTTGGAGAACGCA | TCCATTTCCCGAATGGCAGT |
| CP6T1A | TGCATAATCATGCGCTGCTG | GTCTCCAGCTTACCGCCAAT |
| CP6T1B | CGCGCACATCTTTACTCAGC | GCCAGTAACAAGAAAGCGGC |
| C12B2 | CAGCTTTCGGATGTTGCGAG | ACCGGCCAGATGGTTTCATT |
| C12B1 | TACGCACACTGCCGAAAGAT | TTCCGGACAAGCACTCTCAC |
| CP306 | CCTGCTCGCGCTATTAGTCA | TTCAAGAATTCCCGCACCGA |
| CP304A | AGCGTCGTGCTGACGATTAT | GTATGCCCATTCGCGTGTTC |
| C6A14 | ACACTGCGGAAATACACGGT | CGAAACGATCGGGTTCAGGA |
| CP4S3 | AAGCGCTGAAGGTACTGCAT | AAGTGTCGACTTCTTCGCGT |
| CP132 | AGCACACCTCTTCAATCCCG | CTGCGATCTCAGCATAACGC |
| CP316 | AATCGGTTCGGTGCAGAAGT | ATGATCTGCGCTGTGTAGCA |
| CP304B | TGAGGTCGTAGGTAGAGGGC | GCTCCGTGTCTACCAATGCT |
| CP6G2 | CGCGCTGTGTTCAAGTTCAG | CGCAGAAACTCGGTAGAGGT |
| S. lycopersicum defensive pathways genes and primers | ||
| PORK1 | AGACCCTCAATGAAAGAGGTA | GGTGGAGCTAGAAGTGAGACA |
| slPPO1 | GTGGACAGGATGTGGAACGA | CTTCTTGGTGTCCAGGCAGT |
| slPPO2 | AGTTGTTGCCCTCCTGTACC | CCCTCATTCGACTCGTAGCC |
| LecRK1 | CTTTGCAGGCATCGTGCTTT | GCGCAAAGGTGAAGGGATTG |
| PII | TGGTGTACCAACAAAGCTTGC | GCATTTGTACAACAAAGCCCA |
| LeMPK1 | GATGGTTCCGTTCCGCAAAC | GAACCTGCCACCATGGCTTA |
| LeMPK2 | GCGCTTGCTCATCCTTACCT | AATCCAACAGCAAACGAGCG |
| LeMPK3 | CGCCCTTACGAAGGGAGTTT | ACTTTAGCCCACGGAGAAGC |
| GGP2 | CCTCCACTTCCAGGCGTATT | GCATCAGACAAATCACGGGC |
| Mi-1.1 | AAAGCTCACCAGTGGATCGG | CCATGCACGAAGGTCGAAAC |
| LOXB | GCGTTTAAGGCTTTGTGCGA | GTAGGCCTTGACCATCCGTT |
| LOXD | GCAGATCGCTAAAGCACACG | GCGCTTAACTGCCTATGTGC |
| CCoAOMT | ACCAAATGATTGACGACGGC | TCCGTTCCAAAGGGTGTTGT |
| LeGAD2 | TGAGCCCTGAGAAAGCTGTG | GGAGTGTCCCACCCTGTTTC |
| a-AIs1 | AAGTGCCTCACCAACACCAT | CAGAATTCGTCGCGGATGGA |
| Treatment/Interaction | df | F | p |
|---|---|---|---|
| 48 h after inoculation | |||
| Ripening stage | 1, 79 | 3.528 | 0.019 |
| Picking status | 1, 79 | 0.461 | 0.499 |
| Ripening * picking status | 1, 79 | 2.475 | 0.120 |
| 120 h after inoculation | |||
| Ripening stage | 1, 79 | 0.012 | 0.913 |
| Picking status | 1, 79 | 10.61 | 0.002 |
| Ripening * picking status | 1, 79 | 4.154 | 0.045 |
| A-Tomato Fruit 48 h | Unpicked | Picked | ||||
|---|---|---|---|---|---|---|
| Gene symbol | Mean of 2−ΔCT (n = 10) | t-value/z-value | df | p-Value | Expressed higher | |
| PORK1 | 0.0084 | 0.0083 | −0.006 | 18 | 0.994 | |
| slPPO1 | 1.86 × 10−4 | 5.77 × 10−5 | −1.568 | - | 0.121 | |
| slPPO2 | 2.98 × 10−4 | 7.61 × 10−5 | −2.238 | 11 | 0.047 * | Unpicked |
| LecRK1 | 0.1370 | 0.1318 | −0.245 | - | 0.586 | |
| PII | 0.3110 | 0.2877 | −0.219 | 18 | 0.828 | |
| LeMPK1 | 0.0968 | 0.0860 | −1.423 | - | 0.212 | |
| LeMPK2 | 0.3408 | 0.3228 | −0.327 | 18 | 0.623 | |
| LeMPK3 | 0.7928 | 0.7578 | −0.173 | 18 | 0.864 | |
| GGP2 | 0.5486 | 0.5602 | 0.222 | 18 | 0.826 | |
| Mi-1.1 | 0.0036 | 0.0035 | −1.937 | - | 0.064 | |
| LOXB | 9.0365 | 11.2538 | 0.497 | 13 | 0.627 | |
| LOXD | 1.0514 | 1.1575 | −0.320 | - | 0.628 | |
| CCoAOMT | 0.4390 | 0.3162 | −1.337 | 18 | 0.197 | |
| LeGAD2 | 0.3698 | 0.3422 | −0.305 | 12 | 0.765 | |
| a-AIs1 | 0.0025 | 0.0015 | −0.489 | - | 0.840 | |
| B-Tomato tissue 120 h | Unpicked | Picked | ||||
| PORK1 | 0.0109 | 0.0108 | −0.024 | 18 | 0.980 | |
| slPPO1 | 0.0006 | 0.0001 | −0.916 | - | 0.361 | |
| slPPO2 | 0.0003 | 0.0001 | −0.239 | - | 0.708 | |
| LecRK1 | 0.2125 | 0.7589 | −0.874 | 9 | 0.404 | |
| PII | 0.0901 | 0.26787 | 0.697 | 10 | 0.501 | |
| LeMPK1 | 0.1596 | 0.2477 | 2.083 | 18 | 0.051 | |
| LeMPK2 | 0.4760 | 0.8220 | −2.419 | - | 0.016 * | Picked |
| LeMPK3 | 0.6701 | 0.6701 | 1.754 | 18 | 0.096 | |
| GGP2 | 0.4207 | 0.4837 | 0.454 | 18 | 0.654 | |
| Mi-1.1 | 0.0229 | 0.0066 | −0.677 | - | 0.619 | |
| LOXB | 5.9229 | 11.8689 | −0.939 | - | 0.421 | |
| LOXD | 3.5808 | 2.1924 | −0.484 | 10 | 0.639 | |
| CCoAOMT | 0.4748 | 0.6363 | 0.652 | 18 | 0.522 | |
| LeGAD2 | 0.2815 | 0.4583 | −1.637 | - | 0.121 | |
| a-AIs1 | 0.0275 | 0.0004 | −0.209 | - | 0.850 | |
| C-Tomato tissue Unpicked | 48 h | 120 h | ||||
| PORK1 | 0.0084 | 0.0109 | −1.008 | 13 | 0.332 | |
| slPPO1 | 1.86 × 10−4 | 0.0006 | −1.408 | - | 0.189 | |
| slPPO2 | 2.98 × 10−4 | 0.0003 | −0.213 | - | 0.834 | |
| LecRK1 | 0.1370 | 0.2125 | −0.995 | - | 0.345 | |
| PII | 0.3110 | 0.0901 | 2.457 | 18 | 0.024 * | 48 h |
| LeMPK1 | 0.0968 | 0.1596 | −1.986 | 12 | 0.069 | |
| LeMPK2 | 0.3408 | 0.4760 | −1.870 | 18 | 0.077 | |
| LeMPK3 | 0.7928 | 0.6701 | 2.496 | 18 | 0.022 * | 48 h |
| GGP2 | 0.5486 | 0.4207 | 1.069 | 11 | 0.308 | |
| Mi-1.1 | 0.0036 | 0.0229 | −1.278 | - | 0.307 | |
| LOXB | 9.0365 | 5.9229 | 1.192 | 18 | 0.248 | |
| LOXD | 1.0514 | 3.5808 | −0.894 | 9 | 0.394 | |
| CCoAOMT | 0.4390 | 0.4748 | −1.159 | - | 0.241 | |
| LeGAD2 | 0.3698 | 0.2815 | −2.449 | - | 0.017 * | 48 h |
| a-AIs1 | 0.0025 | 0.0275 | −1.521 | - | 0.104 | |
| D-Tomato tissue Picked | 48 h | 120 h | ||||
| PORK1 | 0.0083 | 0.0108 | −1.049 | 18 | 0.308 | |
| slPPO1 | 5.77 × 10−5 | 0.0001 | −1.125 | - | 0.289 | |
| slPPO2 | 7.61 × 10−5 | 0.0001 | −0.821 | - | 0.596 | |
| LecRK1 | 0.1318 | 0.7589 | −2.343 | - | 0.021 * | 120 h |
| PII | 0.2877 | 0.26787 | −2.419 | - | 0.022 * | 48 h |
| LeMPK1 | 0.0860 | 0.2477 | −4.255 | 18 | 0.0004 * | 120 h |
| LeMPK2 | 0.3228 | 0.8220 | −3.142 | 10 | 0.010 * | 120 h |
| LeMPK3 | 0.7578 | 0.6701 | 0.441 | 18 | 0.664 | |
| GGP2 | 0.5602 | 0.4837 | 0.877 | 13 | 0.396 | |
| Mi-1.1 | 0.0035 | 0.0066 | −2.873 | - | 0.004 * | 120 h |
| LOXB | 11.2538 | 11.8689 | −0.090 | 18 | 0.928 | |
| LOXD | 1.1575 | 2.1924 | −1.542 | 18 | 0.140 | |
| CCoAOMT | 0.3162 | 0.6363 | −2.301 | 11 | 0.042 * | 120 h |
| LeGAD2 | 0.3422 | 0.4583 | −0.804 | 18 | 0.431 | |
| a-AIs1 | 0.0015 | 0.0004 | −2.041 | - | 0.041 * | 48 h |
| A-Larvae Tissue 48 h | Unpicked | Picked | ||||
|---|---|---|---|---|---|---|
| Gene symbol | Mean of 2−ΔCT (n = 8) | t-value/z-value | df | p-Value | Expressed higher | |
| GST D1 | 0.1000 | 0.0820 | −1.442 | 14 | 0.171 | |
| GST T1 | 0.4758 | 0.3974 | −1.195 | 14 | 0.251 | |
| GST T7 | 0.0023 | 0.0017 | −4.002 | 14 | 0.001 * | Unpicked |
| EST F | 0.0071 | 0.0069 | −0.135 | 14 | 0.894 | |
| EST 1 | 0.00027 | 0.00023 | −1.129 | 14 | 0.277 | |
| SUR | 0.0008 | 0.0006 | −2.078 | 9 | 0.066 | |
| ABCG1 | 0.0031 | 0.0024 | −1.308 | 14 | 0.211 | |
| ABCA3 | 0.0177 | 0.0174 | −0.155 | 14 | 0.878 | |
| L259 | 0.0036 | 0.0037 | −0.184 | - | 0.864 | |
| MDR49 | 0.0063 | 0.0067 | 0.116 | - | 0.955 | |
| CP6A9 | 0.0009 | 0.0007 | −1.216 | 10 | 0.252 | |
| CP313 | 0.0119 | 0.0093 | −1.096 | 14 | 0.291 | |
| CP134 | 0.0014 | 0.0010 | −0.231 | - | 0.867 | |
| CP4D8 | 0.0004 | 0.0003 | −0.547 | 14 | 0.592 | |
| CP6G1 | 0.00160 | 0.00162 | −0.347 | - | 0.779 | |
| C12E1 | 0.0155 | 0.0151 | −0.157 | 14 | 0.877 | |
| CP6T1A | 0.0003 | 0.0002 | −1.042 | 14 | 0.314 | |
| CP6T1B | 0.0004 | 0.0003 | −0.615 | 14 | 0.548 | |
| C12B2 | 0.00046 | 0.00037 | −0.615 | 12 | 0.549 | |
| C12B1 | 0.0005 | 0.0004 | −0.616 | 14 | 0.548 | |
| CP306 | 0.00194 | 0.00193 | −0.030 | 14 | 0.975 | |
| CP304A | 0.00035 | 0.000350 | −0.022 | 14 | 0.982 | |
| C6A14 | 0.0079 | 0.0073 | −0.694 | - | 0.536 | |
| CP4S3 | 0.0051 | 0.0058 | 0.459 | 14 | 0.652 | |
| CP132 | 0.0127 | 0.0169 | 2.203 | 14 | 0.044 * | Picked |
| CP316 | 0.0007 | 0.0005 | −0.243 | 8 | 0.246 | |
| CP304B | 0.00066 | 0.00053 | −0.783 | 14 | 0.446 | |
| CP6G2 | 0.00072 | 0.00059 | −1.288 | 14 | 0.218 | |
| B-Larvae tissue 120 h (n = 7) | Unpicked | Picked | ||||
| GST D1 | 0.1353 | 0.1594 | −0.694 | - | 0.536 | |
| GST T1 | 0.7766 | 1.0905 | −0.958 | - | 0.338 | |
| GST T7 | 0.0108 | 0.0051 | −0.447 | - | 0.710 | |
| EST F | 0.0162 | 0.0138 | −0.343 | 12 | 0.737 | |
| EST 1 | 7.13 × 10−5 | 1.48 × 10−4 | 1.494 | 12 | 0.160 | |
| SUR | 0.0011 | 0.0008 | −0.822 | 12 | 0.426 | |
| ABCG1 | 0.0106 | 0.0095 | −0.960 | 12 | 0.355 | |
| ABCA3 | 0.0328 | 0.0269 | −1.148 | 12 | 0.273 | |
| L259 | 0.0137 | 0.0099 | −0.469 | 12 | 0.647 | |
| MDR49 | 0.0173 | 0.0173 | 0.005 | 12 | 0.995 | |
| CP6A9 | 0.0102 | 0.0059 | −0.443 | 12 | 0.665 | |
| CP313 | 0.0391 | 0.0374 | −0.115 | 12 | 0.909 | |
| CP134 | 0.0308 | 0.0270 | −0.314 | 12 | 0.758 | |
| CP4D8 | 0.0078 | 0.0038 | −0.534 | 12 | 0.602 | |
| CP6G1 | 0.1161 | 0.0278 | −1.224 | - | 0.264 | |
| C12E1 | 0.0827 | 0.0687 | −0.418 | 12 | 0.682 | |
| CP6T1A | 0.0114 | 0.0048 | −0.594 | - | 0.563 | |
| CP6T1B | 0.0103 | 0.0047 | −0.572 | - | 0.577 | |
| C12B2 | 0.0103 | 0.0047 | −0.572 | 12 | 0.576 | |
| C12B1 | 0.01033 | 0.0046 | −0.572 | 12 | 0.577 | |
| CP306 | 0.0062 | 0.0039 | −0.419 | 12 | 0.682 | |
| CP304A | 0.0100 | 0.0061 | −1.214 | - | 0.259 | |
| C6A14 | 0.5602 | 0.0365 | −1.250 | 12 | 0.234 | |
| CP4S3 | 0.0381 | 0.0216 | −1.021 | 12 | 0.327 | |
| CP132 | 0.0249 | 0.0213 | −0.294 | 12 | 0.773 | |
| CP316 | 0.0012 | 0.0008 | −1.456 | 12 | 0.170 | |
| CP304B | 0.0172 | 0.0101 | −0.831 | - | 0.456 | |
| CP6G2 | 0.0084 | 0.0046 | −0.490 | 12 | 0.632 | |
| C-Larvae tissue Unpicked | 48 h | 120 h | ||||
| GST D1 | 0.0961 | 0.1353 | −1.589 | 12 | 0.138 | |
| GST T1 | 0.4367 | 0.7766 | −1.993 | 7 | 0.086 | |
| GST T7 | 0.0024 | 0.0108 | −0.447 | - | 0.701 | |
| EST F | 0.0066 | 0.0162 | −1.853 | - | 0.073 | |
| EST 1 | 2.76 × 10−4 | 7.13 × 10−5 | 5.654 | 10 | 0.0001 * | 48 h |
| SUR | 0.0008 | 0.0011 | −1.081 | 12 | 0.301 | |
| ABCG1 | 0.0028 | 0.0106 | −7.336 | 12 | <0.0001 * | 120 h |
| ABCA3 | 0.0180 | 0.0328 | −3.976 | 12 | 0.001 * | 120 h |
| L259 | 0.0036 | 0.0137 | −2.619 | - | 0.009 * | 120 h |
| MDR49 | 0.0063 | 0.0173 | −1.853 | - | 0.073 | |
| CP6A9 | 0.0009 | 0.0102 | −1.597 | - | 0.110 | |
| CP313 | 0.0113 | 0.0391 | −2.265 | 6 | 0.061 | |
| CP134 | 0.0015 | 0.0308 | −2.668 | 6 | 0.036 * | 120 h |
| CP4D8 | 0.0004 | 0.0078 | −1.118 | 6 | 0.306 | |
| CP6G1 | 0.0017 | 0.1161 | −3.003 | - | 0.003 * | 120 h |
| C12E1 | 0.0161 | 0.0827 | −2.491 | 6 | 0.046 * | 120 h |
| CP6T1A | 0.0003 | 0.0114 | −2.108 | - | 0.040 * | 120 h |
| CP6T1B | 0.0005 | 0.0103 | −2.236 | - | 0.029 * | 120 h |
| C12B2 | 0.0207 | 0.0716 | −3.879 | 7 | 0.005 * | 120 h |
| C12B1 | 0.0010 | 0.0096 | −2.747 | - | 0.007 * | 120 h |
| CP306 | 0.0019 | 0.0062 | −0.971 | 6 | 0.368 | |
| CP304A | 0.0003 | 0.0100 | −3.002 | - | 0.003 * | 120 h |
| C6A14 | 0.0083 | 0.0560 | −3.903 | 6 | 0.007 * | 120 h |
| CP4S3 | 0.0053 | 0.0381 | −2.875 | - | 0.004 * | 120 h |
| CP132 | 0.0128 | 0.0249 | −1.264 | 6 | 0.251 | |
| CP316 | 0.0007 | 0.0012 | −1.729 | 12 | 0.109 | |
| CP304B | 0.0006 | 0.0172 | −2.264 | - | 0.018 * | 120 h |
| CP6G2 | 0.0007 | 0.0084 | −1.164 | 6 | 0.288 | |
| D-Larvae tissue Picked | 48 h | 120 h | ||||
| GST D1 | 0.0835 | 0.1594 | −1.981 | - | 0.053 | |
| GST T1 | 0.4036 | 1.0905 | −0.319 | - | 0.749 | |
| GST T7 | 0.0017 | 0.0051 | −1.214 | - | 0.225 | |
| EST F | 0.0071 | 0.0138 | −1.550 | 6 | 0.168 | |
| EST 1 | 0.0002 | 0.0001 | 1.766 | 7 | 0.117 | |
| SUR | 0.0006 | 0.0008 | −1.331 | 6 | 0.227 | |
| ABCG1 | 0.0024 | 0.0095 | −9.787 | 8 | <0.0001 * | 120 h |
| ABCA3 | 0.0174 | 0.0269 | −2.243 | 7 | 0.058 | |
| L259 | 0.0037 | 0.0099 | −1.089 | - | 0.277 | |
| MDR49 | 0.0069 | 0.0173 | −1.510 | 6 | 0.177 | |
| CP6A9 | 0.0007 | 0.0059 | −0.703 | - | 0.482 | |
| CP313 | 0.0091 | 0.0374 | −2.970 | 6 | 0.023 * | 120 h |
| CP134 | 0.0010 | 0.0270 | −5.289 | 6 | 0.001 * | 120 h |
| CP4D8 | 0.0003 | 0.0038 | −1.995 | 6 | 0.357 | |
| CP6G1 | 0.0009 | 0.0278 | −3.130 | - | 0.002 * | 120 h |
| C12E1 | 0.0153 | 0.0687 | −2.630 | 6 | 0.038 * | 120 h |
| CP6T1A | 0.0002 | 0.0048 | −1.853 | - | 0.064 | |
| CP6T1B | 0.0003 | 0.0047 | −1.981 | - | 0.055 | |
| C12B2 | 0.0194 | 0.0548 | −3.612 | 6 | 0.010 * | 120 h |
| C12B1 | 0.0009 | 0.0061 | −2.492 | - | 0.013 * | 120 h |
| CP306 | 0.0018 | 0.0039 | −0.708 | 6 | 0.504 | |
| CP304A | 0.0003 | 0.0061 | −2.489 | - | 0.015 * | 120 h |
| C6A14 | 0.0075 | 0.0365 | −2.942 | 6 | 0.025 * | 120 h |
| CP4S3 | 0.0060 | 0.0216 | −2.432 | 6 | 0.048 * | 120 h |
| CP132 | 0.0175 | 0.0213 | −0.452 | 6 | 0.665 | |
| CP316 | 0.0005 | 0.0008 | −1.585 | 7 | 0.155 | |
| CP304B | 0.0005 | 0.0101 | −2.236 | - | 0.029 * | 120 h |
| CP6G2 | 0.0005 | 0.0046 | −1.052 | 6 | 0.333 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roohigohar, S.; Clarke, A.R.; Strutt, F.; van der Burg, C.A.; Prentis, P.J. Fruit Fly Larval Survival in Picked and Unpicked Tomato Fruit of Differing Ripeness and Associated Gene Expression Patterns. Insects 2022, 13, 451. https://doi.org/10.3390/insects13050451
Roohigohar S, Clarke AR, Strutt F, van der Burg CA, Prentis PJ. Fruit Fly Larval Survival in Picked and Unpicked Tomato Fruit of Differing Ripeness and Associated Gene Expression Patterns. Insects. 2022; 13(5):451. https://doi.org/10.3390/insects13050451
Chicago/Turabian StyleRoohigohar, Shirin, Anthony R. Clarke, Francesca Strutt, Chloé A. van der Burg, and Peter J. Prentis. 2022. "Fruit Fly Larval Survival in Picked and Unpicked Tomato Fruit of Differing Ripeness and Associated Gene Expression Patterns" Insects 13, no. 5: 451. https://doi.org/10.3390/insects13050451
APA StyleRoohigohar, S., Clarke, A. R., Strutt, F., van der Burg, C. A., & Prentis, P. J. (2022). Fruit Fly Larval Survival in Picked and Unpicked Tomato Fruit of Differing Ripeness and Associated Gene Expression Patterns. Insects, 13(5), 451. https://doi.org/10.3390/insects13050451