Simple Summary
Aphids are major crop pests that are feeding on plant sap and transmitting plant viruses, thus inducing high yield losses worldwide. As chemical pesticides are decreasingly used in plant protection, fungi that cause disease to insects (entomopathogenic fungi) are one of the promising alternatives. They are commonly applied by spraying plants to protect them against herbivores. When applied, some fungi penetrate and live within plant tissues, thus helping to internally protect from insect attacks and other plant diseases. The aim of our study was to assess the effects of entomopathogenic fungi (EPF) applied firstly by contact after insect direct spraying, secondly by endophytic plant inoculation, and thirdly by associated both methods assessing the green peach aphid performances. The impact of the presence of endophytic entomopathogenic fungi (EEPF) in plant tissues on virus transmission by aphids was also considered. We found that the EPF Beauveria bassiana killed the green peach aphid and reduced its fecundity regardless of the application method. On fungal-inoculated plants, there was also a high mortality of aphid nymphs and infection by the potato leafroll virus (PLRV) was delayed by about a week with the EEPF treatment compared to fungal-free plants. This study showed that spraying plant leaves with EPF not only has a direct insecticidal effect against insects but could also have beneficial side effects for the plant against viruses.
Abstract
Aphids are major crop pests that transmit more than half of all insect-vectored plant viruses responsible for high yield losses worldwide. Entomopathogenic fungi (EPF) are biological control agents mainly used by foliar application to control herbivores, including sap-sucking pests such as aphids. Their ability to colonize plant tissues and to interact with diverse plant pathogenic microorganisms have been reported. In our study, we evaluated the effectiveness of Beauveria bassiana ((Balsamo-Crivelli) Vuillemin) directly applied by contact or/and indirectly via endophytism in tobacco plants (Nicotiana tabacum L.) against the virus vector Myzus persicae (Sulzer) carrying the Potato leafroll virus (PLRV) or not. We found that both contact treatment and endophytic colonization of leaves significantly increased aphid mortality and decreased the fecundity rate when compared to control plants. In addition, on fungal-colonized leaves, viruliferous aphids were more negatively impacted than virus-free ones and nymph mortality was significantly higher than on fungal-free plants. Furthermore, we assessed PLRV transmission by M. persicae on tobacco plants inoculated with either B. bassiana or Metarhizium acridum ((Driver and Milner) JF Bischoff, Rehner, and Humber) as source or/and recipient plants. Myzus persicae was found to acquire and transmit PLRV regardless of the treatment. Nevertheless, the infection rate of endophytically colonized plants was lower at a seven-day incubation period and had increased to almost 100% after fifteen days. These results suggest that B. bassiana is effective against aphids, both by contact and via endophytism, and both B. bassiana and M. acridum delayed PLRV infection in tobacco.
1. Introduction
Aphids are major crop pests, not only due to their feeding on phloem sap but mainly because they are plant virus vectors that induce high yield decreases, from 20 to 70% in potato crops [1,2]. Aphids transmit more than half of all insect-vectored plant viruses in different modes [3]. Firstly, plant viruses are acquired and transmitted by non- and semi-persistent modes by successive brief stylet punctures in epidermal and mesophyll cells. The viral particles are essentially retained on the stylet tip of vectors, the latter becoming immediately viruliferous for a short duration (from seconds to minutes) [4]. Secondly, a persistent transmission mode requires a longer phloem-feeding duration by vectors for virus acquisition/transmission. The latent period needs to be higher to allow for virus circulation into the vector before it becomes viruliferous [5,6,7]. For example, Potato leafroll virus (PLRV, Luteoviridae, Polerovirus) is most efficiently transmitted by Myzus persicae (Sulzer) in a persistent circulative mode [7,8,9]. To achieve an efficient transmission rate, more than 48 h of acquisition and inoculation access periods (AAP and IAP, respectively) and from 8 to 123 h of latent periods are needed [7,10,11]. However, regardless of the transmission mode, plant viruses need to multiply and move within different plant organs prior to symptom appearance and probably to be acquired by the vector during phloem feeding.
To control the spread of persistently transmitted viruses, especially PLRV, the use of synthetic chemicals against vector populations was considered to be the best solution for a long time [5,12,13,14]. Indeed, vector mortality probably occurs before the end of the virus transmission process, particularly during the latent period [11]. However, due to environmental side-effects, mainly on non-target beneficial organisms, several chemicals are currently prohibited to be used in agriculture [15,16]. Consequently, alternatives methods with less ecological impact are encouraged [17].
Entomopathogenic fungi (EPF) are biological control agents as alternatives to synthetic chemicals to control sap-feeding pests such as aphids [18,19,20,21,22,23,24,25,26]. To improve the effectiveness of this biocontrol agent, researchers are focusing on the relationships between EPF and insects through their shared host plant [21,27,28,29,30,31,32]. Among EPF, Beauveria spp. (Vuill.) and Metarhizium spp. (Sorokin) (Ascomycota: Clavicipitaceae) are two genera of filamentous fungi that are used as the most commercialized fungal biopesticides [33,34,35,36,37]. They are commonly used in inundative treatments via foliar application [38,39,40,41]. Their capability to transcutaneously infect insects is their major asset [39]. Moreover, recent studies have highlighted their ability to be assimilated and to live internally in plant tissues (i.e., endophytically) without any symptom appearance [42,43,44,45,46,47]. Then, they directly interact with plant pathogens and pests, including virus vectors [48,49,50,51].
The effectiveness of several strains of EPF applied by contact has been demonstrated [20,25,52]. However, insect cadavers showing EPF outgrowth as evidence that mortality was caused by fungal infection were sometimes omitted [53] or very low [52,54,55,56]. The latter may have been due to environmental conditions such as temperature and humidity [57,58,59] but also because insect mortality could be due to the indirect effects EPF through host plants. In fact, studies have shown that endophytic entomopathogenic fungi (EEPF) improve plant resistance by increasing the biosynthesis of secondary metabolites in plant tissues that are toxic for insects [32,60,61] and by activating an induced systemic resistance (ISR) in the plant [21,61,62], which can lead to insect mortality. Then, a direct effect of EPF by contact and an indirect effect via endophytism would act synergistically to improve plant fitness. Furthermore, changes in the behavior of virus vectors in response to an EEPF-colonized plant, as well as their infectious status, were reported on host-seeking and plant-feeding behaviors. Indeed, on the one hand, M. persicae discriminated against tomato plants (Solanum lycopersicum L.) colonized with B. bassiana [63] and switched from PLRV-infected plants to healthy potato plants (Solanum tuberosum L.) after virus acquisition [64]. On the other hand, significant changes were detected on electrical penetration graph (EPG) variables, including the intracellular probe (pd) duration and other sequential variables, when Aphis gossypii (Glover) fed on B. bassiana-colonized melon plants (Cucumis melo L.) [50] and M. persicae fed on Potato virus Y (PVY)-infected tobacco [65]. Additionally, M. persicae carrying PLRV or not were found to be both attracted to the tobacco leaves colonized by Beauveria bassiana ((Balsamo-Crivelli) Vuillemin) or Metarhizium acridum ((Driver and Milner) JF Bischoff, Rehner, and Humber) [66]. This was not the desired effect because biological control agents are supposed to protect plants from pests [63,66]. Therefore, estimating the fitness of an insect living on an EEPF-colonized plant and its ability to transmit a phytovirus is decisive for assessing the true role of EPF endophytism. In addition, it was reported that the susceptibility of viruliferous compared to non-viruliferous insects may differ [67].
In this study, we first evaluated the virulence effect of B. bassiana applied directly by contact or indirectly via endophytism through tobacco plants against M. persicae carrying PLRV or not. Secondly, we assessed the transmission rate of PLRV by M. persicae on tobacco plants inoculated with either B. bassiana or M. acridum as recipient plants and fungal sources, respectively.
2. Materials and Methods
2.1. Plants, Insects and Virus
Tobacco seeds (N. tabacum cv. Xanthii) were placed in a germination container with autoclaved potting soil. Seedlings at the three-leaf stage were individually transferred into pots (7 × 7 × 7 cm3) and stored in a growth chamber at 22 ± 1 °C, with 70 ± 10% relative humidity (RH) and a 16 h light period.
A colony of an MpCh4 strain of M. persicae was maintained on tobacco plants in 60 × 60 × 60 cm3 net cages (Bugdorms, MegaView Science Co., Taichung City, Taiwan) and placed in air-conditioned rooms as described above. Plants were replaced every 3 weeks by four-leaf stage seedlings.
Potato leafroll virus was acquired from infected Physalis floridana Rydb. from DSMZ (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Braunschweig, Germany). It was then transferred on tobacco plants by releasing five M. persicae previously virosed on P. floridana for 4 days. Similarly, PLRV was maintained by transferring insects from virus-infected tobacco to new healthy seedlings every 3–4 weeks.
2.2. Entomopathogenic Fungi
The GHA strain of Beauveria bassiana and the IMI330189 strain of M. acridum were isolated from the wettable powders of Botanigard and Green Muscle commercial bioinsecticides, respectively. Thirty-five microliters of the respective product suspended in sterile distilled water with 0.01% Tween® 80 were spread on potato dextrose agar (PDA) with additional chloramphenicol (0.05 g/L) to prevent bacterial contamination. Sealed plates were then placed in a light-free incubator at 25 ± 1 °C for 3 weeks. Spores were harvested by scraping the surface of the agar with a sterile L-shaped spreader and suspended in sterile distilled water with 0.01% Tween® 80. The initial concentration of the spore suspensions was determined using a Neubauer hemocytometer cell. The final suspensions were adjusted to 108 spores/mL and stored at 4 °C. Spore viability was assessed on ready-to-use suspensions by spreading three replicates of 0.1 mL of a diluted suspension at 104 spores/mL on a microscopic slide overlaid with a thin slice of PDA. The germination rate was estimated by counting 100 spores using a 200× magnification microscope after incubation for 24 h at 25 ± 1 °C in the dark. Only suspensions with >96% viability were used within a maximum of 24 h.
2.3. Aphid Fitness Bioassays
Both plants (inoculated with B. bassiana or not) and insects (sprayed with the B. bassiana spore suspension or not) were used with viruliferous and non-viruliferous aphids. Fourteen combinations were tested, as described in Table 1. Metarhizium acridum was tested in this experiment but not accounted for any further due to the low occurrence of endophytically colonized leaves and their distribution among concerned treatments (see Results section).

Table 1.
Different treatment combinations in the insect fitness bioassays. I+: viruliferous, I−: non-viruliferous.
Tobacco plants at the four-leaf stage were inoculated with B. bassiana (n = 6 per treatment) 48 h before the beginning of the experiment. Two basal leaves were sprayed with 2 mL of a B. bassiana suspension using a cosmetic hand spray with a fine mist (0.35 mm nozzle diameter). A plastic zip bag was used to isolate the plant top part in order to prevent any contact with the spores. For fungal-free plants, the fungus suspension was replaced by sterile water with Tween 80 (0.01%). All plants were then covered with a plastic cover (Natureflex™, 160 × 300 mm2) and kept in a climate chamber at 22 ± 1 °C and a 16 h light period.
Aphids that were used in this bioassay were obtained by placing 40–50 adults on five healthy or PLRV-infected plants for 24 h. Then, adults were removed, and nymphs of the same age were maintained on the plants for four days, corresponding to the virus acquisition access period (AAP). Groups of 5 individuals from healthy (n = 48) and PLRV-infected (n = 48) plants were then separately placed in Sterilin® Petri dishes (VWR, Radnor, PA, USA) measuring 35 mm in diameter. Both groups from viruliferous and virus-free aphids were also divided into two groups. First, 24 from each group were sprayed with 1 mL of B. bassiana, while the remaining other 24 were sprayed with 1 mL of sterile distilled water containing 0.01% Tween 80. Finally, 5–10 min after treatment, each 24 aphid batch was further divided into two final groups of twelve. Each of these final sets of twelve aphids was transferred to either a fungal-free plant or a B. basiana-inoculated plant. Five individuals were placed on the two unsprayed leaves of each plant, confined in a clip cage (15 mm diameter and 9.0 mm thickness) [68] made of a polyethylene sleeve for an air-conditioner insulation system (NMC International SA, Weiswampach, Luxembourg).
Every two days for eight days, aphid mortality and fecundity were recorded. Newly deposited nymphs were counted and maintained in clip cages. Aphid fecundity was calculated by dividing the number of newly deposited nymphs by the adult number previously recorded. Adult and nymph cadavers were counted and removed from clip cages. Adult cadavers were transferred into Petri dishes (9 cm diameter) with moist filter paper and incubated at 25 °C for fungal outgrowth examination [52]. At the end of the experiment, all leaves used for the test were immediately sampled and assessed for EEPF colonization. The entire experiment was repeated twice.
2.4. Virus Spread Bioassay
Fifteen plants at the three-leaf stage were infected by confining 5 individuals from PLRV-infected plants in a clip-cage for 5 days. They were then evaluated after 14 days of incubation by the DAS-ELISA (double-antibody sandwich-enzyme-linked immunosorbent assay) (see below). Nine plants with the highest optical density (OD) were selected as virosed source plants. They were separated into 3 groups of three plants. Two groups were inoculated with EPF following the process described above. The first group was inoculated with B. bassiana, the second was inoculated with M. acridum, and the third was sprayed with sterile water with 0.01% Tween 80. Five days post fungal inoculation (dpi), 3 samples from 3 non-sprayed leaves were collected for EEPF examination. At 8 dpi, several adult individuals were released on the source plants (SPs) in order to reproduce. Twenty-four hours later, adults were removed and laid nymphs were maintained for 4 days, corresponding to the virus AAP. Following the EEPF colonization rate, only 1/3 of SPs for the same treatment with the highest EEPF colonization rate (for treated plants) were selected for the bioassays.
Three treatments of recipient plants (RPs) were prepared: B. bassiana (Bp)-inoculated plants, M. acridum (Mp)-inoculated plants, and fungal-free plants (Ffp) as a control. Those plants were at the four-leaf stage. Their two basal leaves were sprayed with the corresponding inoculum or sterile water with Tween 80 at 0.01%. At 5 dpi, two leaves (one treated and one non-treated) were sampled for the confirmation of the EEPF colonization of tissues. Each treatment of RP was divided into 3 subgroups of 12 plants. Each SP group corresponded to 3 RP subgroups according to the combinations (see Table 2).
Table 2.
Different treatment combinations in the virus spread bioassay.
At 7 dpi, five individuals from the SP selected for each treatment were confined in a clip cage on one of the RP leaves that were not exposed to the fungal spray. The IAP was 3 days, after which aphids were removed with a brush. Plants were kept at room temperature, as described above for incubation. Samples were collected on the 7th, 11th, and 15th days of incubation from the top of the plants where the virus concentration was probably higher [69].
The PLRV infection in each plant was assessed by a qualitative DAS-ELISA with a DSMZ kit following the manufacturer’s instructions in triplicate. Plants were considered as infected when the average OD was at least two times high than negative controls. The entire experiment was repeated thrice.
2.5. Confirmation of Endophytic Colonization of Tobacco by EPF
Leaves collected immediately after the fitness bioassays, and at the 5th dpi for virus spread, bioassays were evaluated to confirm EEPF colonization. They were first surface-sterilized with a solution containing 0.5% NaOCl and Tween 80 (0.01%), then sterilized in a solution containing 70% ethanol, and finally rinsed thrice in sterile distilled water. After drying on sterilized paper towels, 6 samples of approximately 1.5 cm2 from each leaf were taken using a sterile scalpel blade. Samples from each leaf were first pressed (adaxial and abaxial sides) on a PDA plate to mark an imprint to determine whether any epiphytic spores remained on the leaf surface and next transferred on a new culture medium to incubate. Three samples of 100 µL of final rinse water from each leaf were plated on the PDA to evaluate the disinfection process. All plates were sealed and placed in a light-free incubator at 25 °C for 10 days. Fungal colonies growing from internal plant tissues were visually examined according to the characteristics described by the authors of [70,71] for B. bassiana and M. acridum, respectively. For insect fitness bioassays, when one tissue from a tested leaf showed fungal growth, the whole leaf was classified as being endophytically colonized [43]. The EEPF colonization rate was calculated for each leaf using Formula (1). In any case, only results from leaves with confirmed EEPF colonization were taken into account.
2.6. Statistical Analysis
Aphid fitness data collected on the eighth day of the experiment were processed using the R software. A multiple linear regression model was fitted to compare treatments with plant type, aphid virosed status, fungal application mode, and interactions between factors. The factor experiment was not significant and was therefore removed from the model. Then, data from two replicates of fitness assays were pooled in the same analysis. Requirements for the validity of the test (normality and homoscedasticity of the residual) were checked before applying the analysis of variance (ANOVA) test to the regression model. For each significant difference, a pairwise comparison was applied using the “lsmeans” function of the lsmeans package [72] with the Tukey fitting method. The “cld” function of the multcomp package [73] was used to set up a compact display of the letters of all pairwise comparisons. In addition, the probability of adult survival was estimated using the Kaplan–Meier method [74], and curves were plotted with “survival” and “survminer” packages [75]. The multivariate Cox proportional hazards [76] were also calculated to analyze the effects of factor combinations on adult survival using same packages.
Correlations between the colonization rate of leaf tissue by EEPF and insect fitness data including mortality, fecundity, and nymph mortality rates were evaluated by the Spearman method using the “cor.test” function of the package stats [77,78].
The proportions of PLRV-infected plants to uninfected plants for each incubation period were assessed by Pearson’s chi-squared test. The pairwise tests of independence for nominal data were applied with the “pairwiseNominalIndependence” and the “cldList” functions of the rcompanion package [79] to determine the symmetry between treatments.
The significance threshold for all tests was set at 0.05%.
3. Results
3.1. Endophytic Colonization of Tobacco Plants
The inoculation technique of B. bassiana and M. acridum showed successful colonization rates of leaves ranging from 16.7 to 100.0%. Then, 62 and 12 individual leaves inoculated to B. bassiana and M. acridum, respectively, were considered as endophytically colonized for insect fitness trials. Due to the low occurrence of endophytically colonized leaves and their distribution over the concerned treatments (Mp:I−:sf = 5, Mp:I−:ms = 2, Mp:I+:sf = 4, and Mp:I+:ms = 0; I+: viruliferous and I−: non-viruliferous), aphid fitness essays with M. acridum were not considered for further analysis. However, in virus transmission trials, 83 and 89 recipient plants inoculated with B. bassiana and M. acridum, respectively, were considered as endophytically colonized based on the detection of fungal outgrowth within any of the leaf sections sampled on that plants.
3.2. Direct and Indirect Effect of EPF on Aphid Mortality
Adult mortality was significantly higher on fungal-inoculated plants compared to fungal-free plants (F = 53.45; p < 0.001; Figure 1A). Additionally, the mortality rate of aphids that were directly sprayed with B. bassiana was significantly higher compared to unsprayed insects (F = 14.45; p < 0.001). Similar observations with aphids with or without virus were obtained (F = 1.73; p = 0.189). In contrast, there was a significant effect on the interaction between the aphid infection status and plant treatment (F = 7.68; p < 0.01). Indeed, on plants colonized with EEPF, mortality in virosed aphids was higher than the virus-free ones. Furthermore, there was a positive correlation between adult mortality and the rate of leaf tissue colonization to EEPF (rho = 0.47; S = 63; p < 0.001). However, neither the endophytic colonization and contact treatment interaction nor the interaction between contact treatment and the aphid virosed state produced significant effects on aphid mortality (F = 0.08 and p = 0.771 vs. F = 3.59 and p = 0.061, respectively).
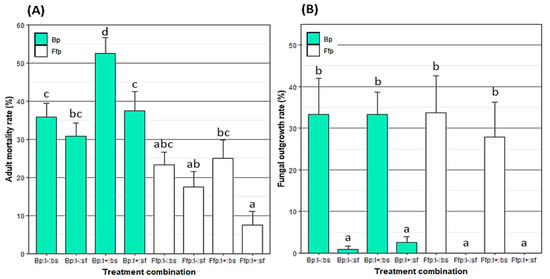
Figure 1.
Mean ± SE of (A) mortality rate (%) of Myzus persicae eight days after exposure to a spray of B. bassiana and/or release on B. bassiana-inoculated plants and (B) fungal outgrowth rate (%) from M. persicae cadavers after ten days of incubation. Treatments were based on combinations of plant type (Bp: B. bassiana-inoculated plant; Ffp: fungal-free plant), insect infectious status (I+: viruliferous; I-: non-viruliferous) and insect spraying (bs: with B. bassiana spore suspension; sf: with spore-free solution). Treatments followed by a different letter differed significantly (p < 0.05). n = Ffp:I−:sf, Ffp:I−:bs, Ffp:I+:sf and Ffp:I+:bs = 24; Bp:I−:sf = 16; Bp:I−:bs = 14; Bp:I+:sf = 15; and Bp:I+:bs = 17.
Direct aphid spraying with a spore suspension was the only significant factor considering the fungal outgrowth on insect cadavers (F = 60.81; p < 0.001). Globally, fungal outgrowth was only observed on the aphid cadavers that were sprayed with B. bassiana (Figure 1B). However, a few cadavers from treatments with plants colonized by EEPF showed fungal outgrowth, but it was not significantly different from those from fungal-free plants (F = 0.27; p = 0.601).
Survival probability according to treatments revealed a significant difference (X2 = 85.2; df = 7; p < 0.001). Aphid survival probability tended to drop between the second and fourth days of EPF spray treatments (Ffp:I−:bs, Ffp:I+:bs, Bp:I−:bs, and Bp:I+:bs; Figure 2). In contrast, treatments made with EEPF-inoculated plants and EPF-unsprayed insects (Bp:I−:sf and Bp:I+:sf) had an accelerated decrease in survival on the sixth and eighth days of observation. Curves related to fungal-free plants were not significantly different except for the Ffp:I+:sf treatment, which had the highest survival probability (92.5%) (Table 3). However, the Bp:I+:bs treatment was the other extreme (47.5%), the only one having reached the median lethal time (LT50) after eight days. Endophytic EPF-inoculation on plants and EPF-spray on aphids were strongly related to mortality risk. Indeed, the fungal-free plant condition was associated with a lower mortality (hazard ratio: HR = 0.41; p < 0.001), while spraying EPF on aphids significantly increased mortality (HR = 1.72; p < 0.001).
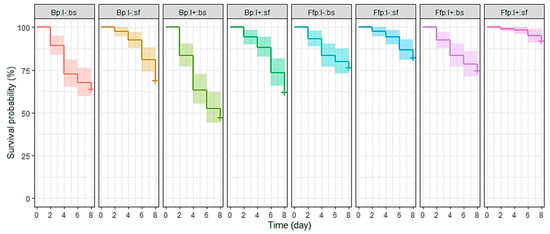
Figure 2.
Survival curves of Myzus persicae for eight days after exposure to a spray of B. bassiana and/or release on B. bassiana-inoculated plants. Treatments were based on combinations of plant type (Bp: B. bassiana-inoculated plant; Ffp: fungal-free plant), insect infectious status (I+: viruliferous; I−: non-viruliferous) and insect spraying (bs: with B. bassiana spore suspension; sf: with spore-free solution). n = Ffp:I−:sf, Ffp:I−:bs, Ffp:I+:sf and Ffp:I+:bs = 120; Bp:I−:sf = 80; Bp:I−:bs = 70; Bp:I+:sf = 75; and Bp:I+:sf = 85.
Table 3.
Pair-wise comparison of Myzus persicae survival curves for eight days after exposure to a spray of B. bassiana and/or release on B. bassiana-inoculated plants. Treatments were based on combinations of plant type (Bp: B. bassiana-inoculated plant; Ffp: fungal-free plant), insect infectious status (I+: viruliferous; I−: non-viruliferous) and insect spraying (bs: with B. bassiana spore suspension; sf: with spore-free solution). n = Ffp:I−:sf, Ffp:I−:bs, Ffp:I+:sf and Ffp:I+:bs = 120; Bp:I−:sf = 80; Bp:I−:bs = 70; Bp:I+:sf = 75; and Bp:I+:sf = 85.
3.3. Direct and Indirect Effect of EPF on Aphid Fecundity and Nymph Mortality
Aphid fecundity analysis per capita was negatively impacted by the presence of EEPF in host plant tissues (F = 228.17; p < 0.001; Figure 3A). In addition, a negative correlation between leaf colonization rate by EEPF and the fecundity per capita was observed (rho = −0.66; S = 20; p < 0.001). Similarly, in treatments where aphids were exposed to a direct contact with B. bassiana, the reproduction rate was significantly decreased (F = 37.36; p < 0.001). Different responses were observed according to aphid virosed status and the B. bassiana colonization of their host plant. The fecundity rate of virosed aphids was significantly lower than virus-free ones (F = 12.06; p < 0.001). Finally, there was no synergistic effect between contact and endophytic plant colonization with EPF (F = 0.45; p = 0.501).
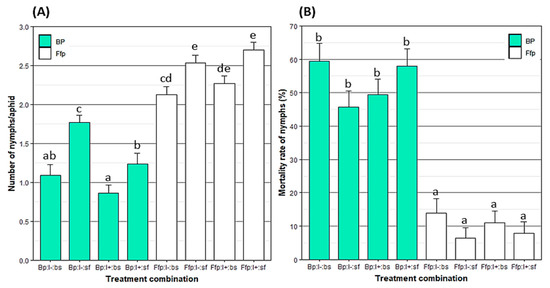
Figure 3.
Mean ± SE of (A) fecundity per capita of Myzus persicae and (B) mortality rate of nymphs laid eight days after release on B. bassiana-inoculated or fungal-free plants. Treatments were based on combinations of plant type (Bp: B. bassiana-inoculated plant; Ffp: fungal-free plant), insect infectious status (I+: viruliferous; I−: non-viruliferous) and insect spraying (bs: with B. bassiana spore suspension; sf: with spore-free solution). Treatments followed by a different letter differ significantly (p < 0.05). n = Ffp:I−:sf, Ffp:I−:bs, Ffp:I+:sf and Ffp:I+:bs = 36; Bp:I−:sf = 31; Bp:I−:bs = 24; Bp:I+:sf = 22; and Bp:I+:sf = 34.
The mortality rate of nymphs laid on endophytically-colonized leaves were significantly higher than those laid on non-treated leaves (F = 194.21; p < 0.001; Figure 3B). In addition, a strong positive correlation was detected between nymph mortality and the B. bassiana colonization rate of leaf tissues (rho = 0.72; S = 331,765; p < 0.001). The direct spraying of B. bassiana spore suspensions did not significantly affect the nymph mortality (F = 1.59; p = 0.208). Similarly, there was no significant effect of insect virosed status (F = 0.003; p = 0.957). In contrast, a significant effect of interaction between direct fungal spraying and aphid virosed status was observed (F = 4.57; p < 0.03).
3.4. Effect of EEPF Colonization on Virus Spread by Aphids
The effect of EEPF on the virus transmission rate was significant for the seven-day (X2 = 50.94; df = 8; p < 0.001) and eleven-day (Χ2 = 40.00; df = 8; p < 0.001) incubation periods (Figure 4A,B). The highest contamination rates were observed at the Ffp–Ffp and Ffp–Mp treatments with 94.4% and 83.3% of infected plants, respectively, on the seventh day of the incubation period. The lowest contamination rates on the seventh day of incubation were observed for the Bp–Bp and Bp–Mp treatments with 33.3% of infected plants. This had gradually increased to 41.7% and 66.7% for Bp–Bp and Bp–Mp, respectively, and reached 100% at the 15th day of incubation. An overall reduction in the PLRV incidence was observed on recipient plants treated with B. bassiana (Mp–Bp and Ffp–Bp) and/or whose aphid vectors were derived from plants inoculated with B. bassiana (Bp–Ffp, Bp–Mp, and Bp–Bp). An intermediate level of contamination was observed on treatments from combinations of fungal-free plants and those inoculated with M. acridum as source or recipient plant (Mp–Ffp, Mp–Mp, and Ffp–Mp).
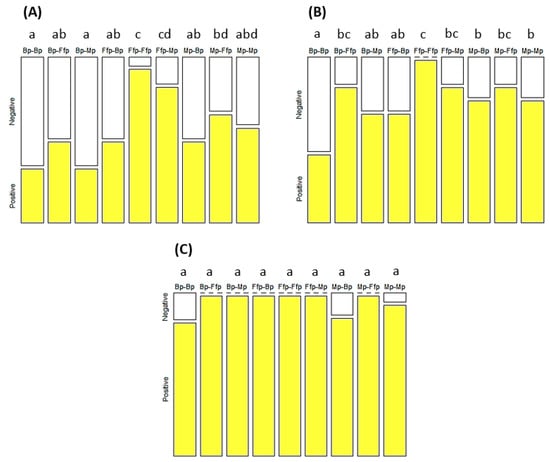
Figure 4.
Percentage of PLRV-infected (positive) and non-infected (negative) tobacco after seven (A), eleven (B), and fifteen (C) days of incubation. Treatments were based on combinations of B. bassiana-inoculated plants (Bp), M. acridum-inoculated plants (Mp), and fungal-free plants (Ffp), as well as a control, as source (two first letters) and recipient (two last letters) plants. Treatments followed by different letters were significantly different (p < 0.05). n = Bp–Ffp, Mp–Ffp, and Ffp–Ffp = 36; Bp–Bp = 32; Bp–Mp = 29; Ffp–Bp = 26; Ffp–Mp = 30; Mp–Bp = 25; and Mp–Mp = 30.
4. Discussion
The evaluation of the EEPF inoculation technique revealed successful colonization of tobacco leaves by B. bassiana and M. acridum from 16.7 to 100.0% of leaf tissues from unsprayed leaves on the same plants as that were treated with a spore suspension. However, there were less M. acridum-colonized leaves for insect fitness trials than those for virus propagation. This was justified by the inclusion of leaves sprayed with spore suspensions for virus propagation trials. The improvement of endophytic colonization-detection techniques could help to properly quantify the extent of plant colonization to EEPF by reducing the risk of false negatives [27].
The effectiveness of B. bassiana as a biological control agent against phloem-feeding insects and especially aphids are well-known and widely documented [21,26,28,30,80,81,82,83,84,85,86]. The most common treatment method use in the field and in the laboratory is the direct contact between the EPF spores and the insect pest [87,88,89,90]. It is also known that the infection of EPF occurs through spore adhesion and germination on the insect cuticle, penetration into the organism by enzymatic activity and mechanical pressure, the invasion of the insect tissues and diffusion of toxins leading to death, and, finally, mycelia growth on the insect cadaver in the saprophytic phase of EPF [91,92,93]. In our study, we showed that the mortality rate was significantly increased and that there was B. bassiana mycelia growth on the aphids that were sprayed with the spore suspension. Additionally, the mortality rate was significantly higher in aphids settled on the plant leaves that were endophytically colonized by B. bassiana. These results are in accordance with several studies such as those from Gurulingappa et al. [94] and Lopez et al. [30], who evaluated A. gossypii survival on cotton (Gossypium hirsutum L.) colonized to B. bassiana, as well as Manoussopoulos et al. [26], who used M. persicae and Chaetosiphon fragaefolii (Cockerell) on strawberry Fragaria × ananassa (Duchesne) plants. These studies provided evidence that B. bassiana was also indirectly effective against aphids via endophytism. Even if Allegrucci et al. [81] did not find significant aphid mortality differences on pepper (Capsicum annuum L.) endophytically colonized by B. bassiana and control plants, aphids more numerously died on B. bassiana-inoculated plants. Such an increased mortality in aphids settled on endophytically colonized plants was previously attributed to a possible improvement in plant resistance conferred by EEPF [27,95]. This hypothesis was supported by our results that showed a positive correlation between adult mortality and leaf EEPF colonization rate. Unlike aphids sprayed with the EPF spore suspension, the mycosis rate was very low (1–3%) in aphid cadavers derived from endophytically colonized leaves. In most published studies concerning the effect of EEPF on insects, mycosis has either not been reported [96,97] or not observed [94,98], except for in stem-borers or leaf mining insects [31,99]. For phloem-feeding pests including aphids, some studies have suggested that EEPF provide the overall insecticidal protection by the biosynthesis of toxic secondary metabolites that could: (1) directly reach insects via plant tissues in which they feed [32,60,61] or (2) induce a systemic resistance by stimulating the plant immune system [21,62] causing feeding deterrence, antibiosis, or global changes in metabolism of the host plant, thus impacting its suitability towards herbivores [43]. Mycelium can also be found in the epidermal cells, palisade parenchyma, intercellular spaces, and vascular elements of the xylem of EEPF-colonized plants [44,100]. This could possibly infect insects, as observed in our study.
The number of nymphs produced per adult was significantly lower with treatments where the insects were sprayed with the spores or/and their plant was endophytically colonized with B. bassiana. Regarding treatment by contact, similar results were obtained by Baverstock et al. [86] with Acyrthosiphon pisum (Harris) infected with Pandora neoaphidis and B. bassiana, as well as by Gurulingappa et al. [32] with A. gossypii sprayed with spores of Lecanicillium lecanii (Zimm.) and B. bassiana. Shrestha et al. [87] also found that daily nymph production and overall fecundity rate in lettuce aphid, Nasonovia ribisnigri (Mosley) were significantly affected when treated with B. bassiana. During pathogenesis, the sublethal level of EPF could affect the insect’s ability to reproduce through physical damage and intoxication, with secondary metabolites affecting insect reproductive system [87]. This was possibly the mechanism of action responsible of the decrease of aphid fecundity in our study. Furthermore, many other studies have shown that the EEPF colonization of plants significantly reduces aphid fecundity [28,30,32,101]. In addition, Jaber and Araj [28] showed that the fecundity rate of M. persicae settled on sweet pepper leaves colonized by B. bassiana and Metarhizium brunneum (Petch) was significantly reduced over two generations. Interestingly, our results showed a strong negative correlation between the aphid fecundity rate and the EEPF colonization leaf rate. This suggested that the presence of B. bassiana decreased M. persicae fecundity. Similarly, Collinson et al. [102] reported a significant decrease in fecundity rates for Diuraphis noxia (Mordvilko) and Aploneura lentisci (Passerini) on perennial ryegrass cv. Alto (Lolium perenne L.) endophytically colonized by Epichloë festucae. The authors associated these results with the presence of the alkaloids lolitrem B, ergovaline, and peramine identified in colonized plants [102]. Beauveria bassiana produces several molecules in vivo and in vitro with antimicrobial, insecticidal, and cytotoxic activities such as bassianin, tennelin, beauvericin, bassianolide, pyridomacrolidin, and pyridovericin [61,103,104,105]. However, future investigations should establish a link between the biosynthesis of alkaloids in planta in sufficient amounts required to affect herbivore fitness and evidence of their impact [61,94].
In our case, it was widely demonstrated that the GHA strain of B. bassiana was effective against aphids, including M. persicae [80,86,87,106,107]. Moreover, there was a strong positive correlation between the mortality rate of nymphs and the EEPF colonization rate of leaves on which nymphs were deposited. In addition, our findings showed that the spore spraying treatment had no impact on nymph survival. This would indicate that the nymphs would not be infected by the EPF. Therefore, there would have been no spore transmission, either vertically or horizontally to the nymphs [85]. The precautions taken during the experiment, in particular removing dead insects regularly from the clip cage, helped to avoid the nymph contamination.
Presently, few studies have provided information regarding the susceptibility of virus vectors to biological control agents, particularly when the vectors are carrying phytovirus compared to virus-free aphids. For instance, Rophalosiphum padi was found to be more susceptible to be parasitized by Aphidius colemani (Viereck) when carrying Barley yellow dwarf virus (BYDV) compared to non-viruliferous individuals [108]. Here, our findings revealed that the adult mortality rate recorded with the Bp:I−:Bb treatment was significantly lower compared to its homologous treatment with viruliferous insects (Bp:I+:Bb). In addition, the survival curve of the latter treatment was significantly lower compared to the others. There was also a higher average fecundity for the Bp:I−:Sf treatment than its homologous treatment with viruliferous individuals (Bp:I+:Sf). To our knowledge, we showed for the first time that the virosed state influenced aphid fitness when exposed to an EPF. Virus transmission by aphids leads to complex chemical reactions that mediate the interactions between the plant, the vector, and the virus [109]. Indeed, plants have developed several mechanisms of genetic, metabolic, and physiological resistance that act in complementary ways to promote their defense against viral infections and insect vector infestations [110]. Aphids produce effector proteins contained in their watery saliva that are injected into plants during phloem-feeding with various effects on plant defense according to host species. Within saliva, some viruses are transmitted with induced impact on plant responses. Depending on the kind of virus, the latter can improve plant quality in order to attract non-viruliferous aphids to facilitate viral propagation [111]. A complete analysis of the chemical ecology in this pathosystem would allow us to determine the responses of each component towards EEPF [109]. Such information, as well as the vectorial competence of the insect settled on EEPF-inoculated plants, is very important to understand the plant–EPF–insect–virus relationship, in particular the role of EPF on virus propagation in the field in a multitrophic approach of biological control toward a diversity of bio-aggressors, including pests and diseases.
Regarding the vectorial competence of aphids for an EEPF-colonized plant, overall, M. persicae was able to acquire and to transmit PLRV to a new plant regardless of the source and recipient plant treatment. Indeed, nearly all kinds of plants were found to be positive for PLRV by an ELISA after fifteen days of incubation. This was in accordance with the findings of Gonzalez et al. [50], where the EPG variables related to the inoculation of persistently and non-persistently transmitted viruses were not altered when A. gossypii was fed on C. melo endophytically colonized by B. bassiana. Additionally, a similar study reported significantly lower rates of the transmission of Cucumber mosaic virus (CMV) and Cucurbit aphid-borne yellows virus (CABYV) by A. gossypii on B. bassiana-colonized plants compared to control plants [50]. In our study, the early evaluation revealed a significantly lower infection rate on recipient plants inoculated with EEPF compared to fungal-free plants, especially at the seventh day of incubation. This potential plant resistance against the virus conferred by EEPF was also reported by Jaber and Salem [112], who found that B. bassiana significantly reduced the incidence and severity of Zucchini yellow mosaic virus (ZYMV) on squash (Cucurbita pepo L.). Similarly, the fungal Hypocrea lixii Pat. significantly reduced the disease level caused by Iris yellow spot virus (IYSV) on onion (Allium cepa L.) [113].
Various mechanisms may justify the reduction of virus incidence on EEPF-colonized plants. This include the activation of the plant’s defense system and the antibiosis effect of EEPF secondary metabolites against viruses [50,61,112,113,114,115]. Accordingly, oosporein, one of the major substances synthesized by B. bassiana, and destruxins synthesized by Metarhizium spp. have antiviral activity [60,104,116]. Destruxins have been reported to be produced in endophytically colonized plant tissues [116], but nothing has been reported about oosporein. In all cases, the interaction of destruxins with plant-viruses in plant tissue should still be demonstrated. Furthermore, space and resource competition between EEPF and a virus could directly affect the location and movement of a virus in plant tissues [50,112,117]. Endophytic strains of B. bassiana and Metarhizium spp. can systemically reach all parts of the plant-colonizing intercellular spaces and vascular xylem elements upwards and downwards from the inoculation point [100,112,118,119]. Then, the intercellular movement of the virus could be delayed or even inhibited by the EEPF [112]. This could possibly explain the low infection rate detected in our study during early plant evaluation. Jaber and Salem [112] found that ZYMV symptom development was delayed after mechanical inoculation on squash colonized with B. bassiana. However, the plant protective duration by EEPF against the virus would have been limited. Indeed, the colonization rate of plant tissue decreased with time [81] depending on several factors, including the inoculation method, fungal isolate, plant species, and environmental conditions [36,46,47]. For instance, in tobacco, tissue colonization by B. bassiana inoculated by the foliar spray method dropped from 100 to 20% twenty days after inoculation [47]. Then, in our study, the fifteenth day of incubation corresponded to the twenty-third day after fungal inoculation. The increase in the virus infection rate after late plant evaluation would have been the result of EEPF decline in plant tissue.
5. Conclusions
This study confirmed the effectiveness of B. bassiana as a biological control agent against aphids and especially M. persicae. The effects of B. bassiana on aphid fitness were observed by using both direct contact by spore spraying and indirect exposure via endophytic setting. There was no interaction between the two modes of application. However, to our knowledge, we report, for the first time, that virosed aphid fitness was negatively impacted compared to virus-free vector when exposed to EPF. In addition, in B. bassiana- and M. acridum-colonized plants, the infection of PLRV on plants was delayed for about one week compared to the control. This suggests a possible plant resistance against virus due to the presence of EEPF in plant tissues. Analyzing the chemical ecology in this pathosystem would allow us to determine the role of EEPF in order for it to be integrated to a crop protection strategy in multitarget pest.
Author Contributions
Conceptualization, J.C.F.-M.; data curation, J.C.F.-M.; funding acquisition, F.F.; methodology, J.C.F.-M. and F.F.; supervision, F.F.; writing—original draft, J.C.F.-M.; writing—review and editing, T.B., and F.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this study are available in this article.
Acknowledgments
The authors are grateful to all those who contributed either way to complete this study. These are Félicien Tosso Dji-ndé, Grégoire Noël and Lucie Tamisier.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lin, F.J.; Bosquée, E.; Liu, Y.J.; Chen, J.L.; Yong, L.; Francis, F. Impact of aphid alarm pheromone release on virus transmission efficiency: When pest control strategy could induce higher virus dispersion. J. Virol. Methods 2016, 235, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Whitworth, J.L.; Nolte, P.; McIntosh, C.; Davidson, R. Effect of Potato virus Y on yield of three potato cultivars grown under different nitrogen levels. Plant Dis. 2006, 90, 73–76. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Capinera, J.L. Green Peach Aphid, Myzus persicae (Sulzer) (Hemiptera: Aphididae). In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 1727–1730. [Google Scholar] [CrossRef]
- Moreno, A.; Tjallingii, W.F.; Fernandez-Mata, G.; Fereres, A. Differences in the mechanism of inoculation between a semi-persistent and a non-persistent aphid-transmitted plant virus. J. Gen. Virol. 2012, 93, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Radcliffe, E.B.; Ragsdale, D.W. Aphid-transmitted potato viruses: The importance of understanding vector biology. Am. J. Potato Res. 2002, 79, 353–386. [Google Scholar] [CrossRef]
- Eskandari, F.; Sylvester, E.S.; Richardson, J. Evidence for lack of propagation of Potato leaf roll virus in its aphid vector, Myzus persicae. Phytopathology 1979, 69, 45. [Google Scholar] [CrossRef]
- Garret, A.; Kerlan, C.; Thomas, D. Ultrastructural study of acquisition and retention of potato leafroll luteovirus in the alimentary canal of its aphid vector, Myzus persicae Sulz. Arch. Virol. 1996, 141, 1279–1292. [Google Scholar] [CrossRef]
- Kotzampigikis, A.T.; Hristova, D.; Tasheva-Terzieva, E. Virus-vector relationship between Potato leafroll virus PLRV and Myzus persicae Sulzer. Bulg. J. Agric. Sci. 2010, 16, 412–421. [Google Scholar]
- Loebenstein, G. Potato leafroll virus (PLRV; Genus Polerovirus; Family Luteoviridae). In Virus and Virus-like Diseases of Potatoes and Production of Seed-Potatoes; Loebenstein, G., Brunt, A.A., Eds.; Kluwer Academic Publishers: Amsterdam, The Netherlands; Springer Netherlands: Dordrecht, The Netherlands, 2001; pp. 69–75. [Google Scholar] [CrossRef]
- Tanaka, S.; Shiota, H. Latent period of Potato leaf roll virus in the green peach aphid (Myzus persicae Sulzer). Jpn. J. Phytopathol. 1970, 36, 106–111. [Google Scholar] [CrossRef]
- Mowry, T.M. Insecticidal reduction of Potato leafroll virus transmission by Myzus persicae. Ann. Appl. Biol. 2005, 146, 81–88. [Google Scholar] [CrossRef]
- Alvarez, J.M.; Srinivasan, R. Evaluation of Hairy Nightshade as an Inoculum Source for Aphid-Mediated Transmission of Potato Leafroll Virus. J. Econ. Entomol. 2005, 98, 1101–1108. [Google Scholar] [CrossRef]
- DiFonzo, C.D.; Ragsdale, D.W.; Radcliffe, E.B. Potato leafroll virus spread in differentially resistant potato cultivars under varying aphid densities. Am. Potato J. 1995, 72, 119–132. [Google Scholar] [CrossRef]
- Agrios, G.N. Plant Pathology, 5th ed.; Elsevier: Burlington, NJ, USA, 2005; Volume 1. [Google Scholar] [CrossRef]
- Shah, P.A.; Pell, J.K. Entomopathogenic fungi as biological control agents. Appl. Microbiol. Biotechnol. 2003, 61, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Peshin, R.; Vasanthakumar, J.; Kalra, R. Diffusion of innovation theory and integrated pest management. In Integrated Pest Managment: Dissemination and Impact; Peshin, R., Dhawan, A.K., Eds.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2009; Volume 2, pp. 1–30. [Google Scholar]
- Vernon, R.S.; van Herk, W.G. Wireworms as Pests of Potato. In Insect Pests of Potato; Elsevier: Amsterdam, The Netherlands, 2013; pp. 103–164. [Google Scholar] [CrossRef]
- Khan, S. Bioassay and enzymatic comparison of six entomopathogenic fungal isolates for virulence or toxicity against green peach aphids Myzus persicae. Afr. J. Biotechnol. 2012, 11, 14193–14203. [Google Scholar] [CrossRef]
- Roy, H.E.; Baverstock, J.; Chamberlain, K.; Pell, J.K. Do aphids infected with entomopathogenic fungi continue to produce and respond to alarm pheromone? Biocontrol. Sci. Technol. 2005, 15, 859–866. [Google Scholar] [CrossRef]
- Seye, F.; Bawin, T.; Boukraa, S.; Zimmer, J.Y.; Ndiaye, M.; Delvigne, F.; Francis, F. Effect of entomopathogenic Aspergillus strains against the pea aphid, Acyrthosiphon pisum (Hemiptera: Aphididae). Appl. Entomol. Zool. 2014, 49, 453–458. [Google Scholar] [CrossRef]
- Clifton, E.H.; Jaronski, S.T.; Coates, B.S.; Hodgson, E.W.; Gassmann, A.J. Effects of endophytic entomopathogenic fungi on soybean aphid and identification of Metarhizium isolates from agricultural fields. PLoS ONE 2018, 13, e0194815. [Google Scholar] [CrossRef] [PubMed]
- Hatano, E.; Baverstock, J.; Kunert, G.; Pell, J.K.; Weisser, W.W. Entomopathogenic fungi stimulate transgenerational wing induction in pea aphids, Acyrthosiphon pisum (Hemiptera: Aphididae). Ecol. Entomol. 2012, 37, 75–82. [Google Scholar] [CrossRef]
- Vu, V.H.; Hong, S., II; Kim, K. Selection of entomopathogenic fungi for aphid control. J. Biosci. Bioeng. 2007, 104, 498–505. [Google Scholar] [CrossRef]
- Koch, K.A. The Role of Entomopathogenic Fungi in the Management of Soybean Aphid. Ph.D. Thesis, University of Minnesota, Minneapolis, MN, USA, 2011. [Google Scholar]
- Saranya, S.; Ushakumari, R.; Jacob, S.; Philip, B.M. Efficacy of different entomopathogenic fungi against cowpea aphid, Aphis craccivora (Koch). J. Biopestic. 2010, 3, 138–142. [Google Scholar]
- Manoussopoulos, Y.; Mantzoukas, S.; Lagogiannis, I.; Goudoudaki, S.; Kambouris, M. Effects of three strawberry entomopathogenic fungi on the prefeeding behavior of the aphid Myzus persicae. J. Insect Behav. 2019, 32, 99–108. [Google Scholar] [CrossRef]
- González-Mas, N.; Sánchez-Ortiz, A.; Valverde-García, P.; Quesada-Moraga, E. Effects of endophytic entomopathogenic ascomycetes on the life-history traits of Aphis gossypii Glover and its interactions with melon plants. Insects 2019, 10, 165. [Google Scholar] [CrossRef] [PubMed]
- Jaber, L.R.; Araj, S.-E. Interactions among endophytic fungal entomopathogens (Ascomycota: Hypocreales), the green peach aphid Myzus persicae Sulzer (Homoptera: Aphididae), and the aphid endoparasitoid Aphidius colemani Viereck (Hymenoptera: Braconidae). Biol. Control 2018, 116, 53–61. [Google Scholar] [CrossRef]
- González-Mas, N.; Medina, M.C.; Sánchez, F.G.; Moraga, E.Q. Bottom—Up effects of endophytic Beauveria bassiana on multitrophic interactions between the cotton aphid, Aphis gossypii, and its natural enemies in melon. J. Pest Sci. 2019, 92, 1271–1281. [Google Scholar] [CrossRef]
- Lopez, D.C.; Zhu-Salzman, K.; Ek-Ramos, M.J.; Sword, G.A. The entomopathogenic fungal endophytes Purpureocillium lilacinum (formerly Paecilomyces lilacinus) and Beauveria bassiana negatively affect cotton aphid reproduction under both greenhouse and field conditions. PLoS ONE 2014, 9, e103891. [Google Scholar] [CrossRef]
- Vidal, S.; Jaber, L.R. Entomopathogenic fungi as endophytes: Plant–endophyte–herbivore interactions and prospects for use in biological control. Curr. Sci. 2015, 109, 46–54. [Google Scholar] [CrossRef]
- Gurulingappa, P.; McGee, P.A.; Sword, G. Endophytic Lecanicillium lecanii and Beauveria bassiana reduce the survival and fecundity of Aphis gossypii following contact with conidia and secondary metabolites. Crop Prot. 2011, 30, 349–353. [Google Scholar] [CrossRef]
- Skinner, M.; Parker, B.L.; Kim, J.S. Chapter 10—Role of Entomopathogenic Fungi in Integrated Pest Management Margaret; Elsevier Inc.: Amsterdam, The Netherlands, 2014. [Google Scholar] [CrossRef]
- Jaronski, S.T. Chapter 11—Mass Production of Entomopathogenic Fungi: State of the Art. In Mass Production of Beneficial Organisms; Morales-Ramos, J.A., Rojas, M.G., Shapiro-Ilan, D.I., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 357–413. [Google Scholar] [CrossRef]
- Chandler, D. Chapter 5—Basic and Applied Research on Entomopathogenic Fungi; Elsevier Inc.: Amsterdam, The Netherlands, 2017. [Google Scholar] [CrossRef]
- Klieber, J.; Reineke, A. The entomopathogen Beauveria bassiana has epiphytic and endophytic activity against the tomato leaf miner Tuta absoluta. J. Appl. Entomol. 2016, 140, 580–589. [Google Scholar] [CrossRef]
- Lacey, L.A. Chapter 1—Entomopathogens Used as Microbial Control Agents; Elsevier Inc.: Amsterdam, The Netherlands, 2017. [Google Scholar] [CrossRef]
- Pedrini, N.; Crespo, R.; Juárez, M.P. Biochemistry of insect epicuticle degradation by entomopathogenic fungi. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 146, 124–137. [Google Scholar] [CrossRef]
- Bawin, T.; Seye, F.; Boukraa, S.; Zimmer, J.-Y.; Delvigne, F.; Francis, F. La lutte contre les moustiques (Diptera: Culicidae): Diversité des approches et application du contrôle biologique. Can. Entomol. 2015, 147, 476–500. [Google Scholar] [CrossRef]
- Jaronski, S.T.; Mascarin, G.M. Mass Production of Fungal Entomopathogens. In Microbial Control of Insect and Mite Pests; Elsevier: Amsterdam, The Netherlands, 2017; pp. 141–155. [Google Scholar] [CrossRef]
- Jaronski, S.T.; Jackson, M.A. Mass Production of Entomopathogenic Hypocreales, 2nd ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2012. [Google Scholar] [CrossRef]
- Guesmi-Jouini, J.; Garrido-Jurado, I.; López-Díaz, C.; Ben Halima-Kamel, M.; Quesada-Moraga, E. Establishment of fungal entomopathogens Beauveria bassiana and Bionectria ochroleuca (Ascomycota: Hypocreales) as endophytes on artichoke Cynara scolymus. J. Invertebr. Pathol. 2014, 119, 1–4. [Google Scholar] [CrossRef]
- Rondot, Y.; Reineke, A. Endophytic Beauveria bassiana in grapevine Vitis vinifera (L.) reduces infestation with piercing-sucking insects. Biol. Control 2018, 116, 82–89. [Google Scholar] [CrossRef]
- Elliot, S.L.; Sabelis, M.W.; Janssen, A.; Van der Geest, L.P.S.; Beerling, E.A.M.; Fransen, J. Can plants use entomopathogens as bodyguards? Ecol. Lett. 2000, 3, 228–235. [Google Scholar] [CrossRef]
- Vega, F.E.; Posada, F.; Catherine Aime, M.; Pava-Ripoll, M.; Infante, F.; Rehner, S.A. Entomopathogenic fungal endophytes. Biol. Control 2008, 46, 72–82. [Google Scholar] [CrossRef]
- Biswas, C.; Dey, P.; Satpathy, S.; Satya, P. Establishment of the fungal entomopathogen Beauveria bassiana as a season long endophyte in jute (Corchorus olitorius) and its rapid detection using SCAR marker. BioControl 2012, 57, 565–571. [Google Scholar] [CrossRef]
- Russo, M.L.; Pelizza, S.A.; Cabello, M.N.; Stenglein, S.A.; Scorsetti, A.C. Endophytic colonisation of tobacco, corn, wheat and soybeans by the fungal entomopathogen Beauveria bassiana (Ascomycota, Hypocreales). Biocontrol Sci. Technol. 2015, 25, 475–480. [Google Scholar] [CrossRef]
- Rúa, M.A.; McCulley, R.L.; Mitchell, C.E. Fungal endophyte infection and host genetic background jointly modulate host response to an aphid-transmitted viral pathogen. J. Ecol. 2013, 101, 1007–1018. [Google Scholar] [CrossRef]
- Lehtonen, P.T.; Helander, M.; Siddiqui, S.A.; Lehto, K.; Saikkonen, K. Endophytic fungus decreases plant virus infections in meadow ryegrass (Lolium pratense). Biol. Lett. 2006, 2, 620–623. [Google Scholar] [CrossRef] [PubMed][Green Version]
- González-Mas, N.; Quesada-Moraga, E.; Plaza, M.; Fereres, A.; Moreno, A. Changes in feeding behaviour are not related to the reduction in the transmission rate of plant viruses by Aphis gossypii (Homoptera: Aphididae) to melon plants colonized by Beauveria bassiana (Ascomycota: Hypocreales). Biol. Control 2019, 130, 95–103. [Google Scholar] [CrossRef]
- Guy, P.L. Barley yellow dwarf viruses in Japanese pasture grasses and lack of correlation with the presence of fungal endophytes. Plant Pathol. 1993, 42, 1–5. [Google Scholar] [CrossRef]
- Shan, L.T.; Feng, M.G. Evaluation of the biocontrol potential of various Metarhizium isolates against green peach aphid Myzus persicae (Homoptera: Aphididae). Pest Manag. Sci. 2010, 66, 669–675. [Google Scholar] [CrossRef]
- Sookar, P.; Bhagwant, S.; Awuor Ouna, E. Isolation of entomopathogenic fungi from the soil and their pathogenicity to two fruit fly species (Diptera: Tephritidae). J. Appl. Entomol. 2008, 132, 778–788. [Google Scholar] [CrossRef]
- Baverstock, J.; Clark, S.J.; Alderson, P.G.; Pell, J.K. Intraguild interactions between the entomopathogenic fungus Pandora neoaphidis and an aphid predator and parasitoid at the population scale. J. Invertebr. Pathol. 2009, 102, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Quintela, E.D.; Mascarin, G.M.; da Silva, R.A.; Barrigossi, J.A.F.; da Martins, J.F.S. Enhanced susceptibility of Tibraca limbativentris (Heteroptera: Pentatomidae) to Metarhizium anisopliae with sublethal doses of chemical insecticides. Biol. Control 2013, 66, 56–64. [Google Scholar] [CrossRef]
- Mbata, G.N.; Shapiro-Ilan, D. The potential for controlling Pangaeus bilineatus (Heteroptera: Cydnidae) using a combination of entomopathogens and an insecticide. J. Econ. Entomol. 2013, 106, 2072–2076. [Google Scholar] [CrossRef] [PubMed]
- Klinger, E. Susceptibility of Adult Colorado Potato Beetle (Leptinotarsa decemlineata) to the Fungal Entomopathogen Beauveria bassiana. Master’s Thesis, The University of Maine, Orono, ME, USA, 2003. [Google Scholar]
- Inglis, G.D.; Johnson, D.L.; Goettel, M.S. Effects of temperature and thermoregulation on mycosis by Beauveria bassiana in grasshoppers. Biol. Control 1996, 7, 131–139. [Google Scholar] [CrossRef]
- Fargues, J. Étude des conditions d’infection des larves de doryphore, Leptinotarsa decemlineata say, par Beauveria bassiana (Bals.) Vuill. [Fungi imperfecti]. Entomophaga 1972, 17, 319–337. [Google Scholar] [CrossRef]
- Liu, B.; Tzeng, Y. Development and applications of destruxins: A review. Biotechnol. Adv. 2012, 30, 1242–1254. [Google Scholar] [CrossRef]
- Jaber, L.R.; Ownley, B.H. Can we use entomopathogenic fungi as endophytes for dual biological control of insect pests and plant pathogens? Biol. Control 2017, 107, 50–59. [Google Scholar] [CrossRef]
- Van Wees, S.C.; Van der Ent, S.; Pieterse, C.M. Plant immune responses triggered by beneficial microbes. Curr. Opin. Plant Biol. 2008, 11, 443–448. [Google Scholar] [CrossRef]
- Aragón, S.M. How Entomopathogenic Endophytic Fungi Modulate Plant-Insect Interactions. Ph.D. Thesis, Georg-August-University Goettingen, Göttingen, Germany, 2016. [Google Scholar]
- Rajabaskar, D.; Bosque-Pérez, N.A.; Eigenbrode, S.D. Preference by a virus vector for infected plants is reversed after virus acquisition. Virus Res. 2014, 186, 32–37. [Google Scholar] [CrossRef]
- Liu, J.; Liu, Y.; Donkersley, P.; Dong, Y.; Chen, X.; Zang, Y.; Xu, P.; Ren, G. Preference of the aphid Myzus persicae (Hemiptera: Aphididae) for tobacco plants at specific stages of potato virus Y infection. Arch. Virol. 2019, 164, 1567–1573. [Google Scholar] [CrossRef] [PubMed]
- Fingu-Mabola, J.C.; Martin, C.; Bawin, T.; Verheggen, F.J.; Francis, F. Does the infectious status of aphids influence their preference towards healthy, virus-infected and endophytically colonized plants? Insects 2020, 11, 435. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, R.L.; Moscardini, V.F.; Gontijo, P.C.; Sâmia, R.R.; Marucci, R.C.; Budia, F.; Carvalho, G.A. Life history parameters and feeding preference of the green lacewing Ceraeochrysa cubana fed with virus-free and potato leafroll virus-infected Myzus persicae. BioControl 2016, 61, 671–679. [Google Scholar] [CrossRef]
- Haas, J.; Lozano, E.R.; Poppy, G.M. A simple, light clip-cage for experiments with aphids. Agric. For. Entomol. 2018, 20, 589–592. [Google Scholar] [CrossRef]
- van den Heuvel, J.F.J.M.; Boerma, T.M.; Peters, D. Transmission of Potato leafroll virus from plants and artificial diets by Myzus persicae. Phytopathology 1991, 81, 150–154. [Google Scholar] [CrossRef]
- Humber, R.A. Fungi. In Manual of Techniques in Insect Pathology; Elsevier: Amsterdam, The Netherlands, 1997; pp. 153–185. [Google Scholar] [CrossRef]
- Driver, F.; Milner, R.J.; Trueman, J.W.H. A taxonomic revision of Metarhizium based on a phylogenetic analysis of rDNA sequence data. Mycol. Res. 2000, 104, 134–150. [Google Scholar] [CrossRef]
- Lenth, R. V Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Kaplan, E.L.; Meier, P. Nonparametric estimation from incomplete observations. J. Am. Stat. Assoc. 1958, 53, 457. [Google Scholar] [CrossRef]
- Therneau, T.M.; Grambsch, P.M. Modeling Survival Data: Extending the Cox Model; Statistics for Biology and Health; Springer New York: New York, NY, USA, 2000. [Google Scholar] [CrossRef]
- Cox, D.R. Regression models and life-tables. J. R. Stat. Soc. Ser. B 1972, 34, 187–220. [Google Scholar] [CrossRef]
- Best, D.J.; Roberts, D.E. Algorithm AS 89: The Upper Tail Probabilities of Spearman’s Rho. J. R. Stat. Soc. 1975, 24, 377–379. [Google Scholar] [CrossRef]
- Hollander, M.; Wolfe, D.A.; Chicken, E. Nonparametric Statistical Methods, Wiley Series in Probability and Statistics; Wiley: Hoboken, NJ, USA, 2015. [Google Scholar] [CrossRef]
- Mangiafico, S.S. Summary and Analysis of Extension Program Evaluation in R; Version 1.18.1.; Rutgers Co.: New Brunswick, NJ, USA, 2016. [Google Scholar]
- Al-alawi, M.S.; Obeidat, M. Selection of Beauveria bassiana (Balsamo) Vuillemin isolates for management of Myzus persicae (sultzar) (hom.: Aphidae) based on virulence and growth related characteristics. Am. J. Agric. Biol. Sci. 2014, 9, 94–100. [Google Scholar] [CrossRef]
- Allegrucci, N.; Velazquez, M.S.; Russo, M.L.; Vianna, M.F.; Abarca, C.; Scorsetti, A.C. Establishment of the entomopathogenic fungus Beauveria bassiana as an endophyte in Capsicum annuum and its effects on the aphid pest Myzus persicae (Homoptera: Aphididae). Rev. Biol. Trop. 2020, 68, 1084–1094. [Google Scholar] [CrossRef]
- Akbari, S.; Ali Safavi, S.; Ghosta, Y. Efficacy of Beauveria bassiana (Blas.) Vuill. against cabbage aphid Brevicoryne brassicae L. (Hem.: Aphididae) in laboratory condition. Arch. Phytopathol. Plant Prot. 2014, 47, 1454–1458. [Google Scholar] [CrossRef]
- Abd El-Salam, A.M.E.; El-Hawary, F.M.A. Lethal and pathogenic effects of Beauveria bassiana and Lecanicillium lecanii on the adult and nymph of Aphis craccivora Koch. Arch. Phytopathol. Plant Prot. 2011, 44, 57–66. [Google Scholar] [CrossRef]
- Knudsen, G.R.; Schotzko, D.J. Spatial simulation of epizootics caused by Beauveria bassiana in russian wheat aphid populations. Biol. Control 1999, 16, 318–326. [Google Scholar] [CrossRef]
- Dara, S.K.; Dara, S.S. Endophytic colonization and pest management potential of Beauveria bassiana in strawberries. J. Berry Res. 2013, 3, 203–211. [Google Scholar] [CrossRef]
- Baverstock, J.; Roy, H.E.; Clark, S.J.; Alderson, P.G.; Pell, J.K. Effect of fungal infection on the reproductive potential of aphids and their progeny. J. Invertebr. Pathol. 2006, 91, 136–139. [Google Scholar] [CrossRef]
- Shrestha, G.; Enkegaard, A.; Steenberg, T. Laboratory and semi-field evaluation of Beauveria bassiana (Ascomycota: Hypocreales) against the lettuce aphid, Nasonovia ribisnigri (Hemiptera: Aphididae). Biol. Control 2015, 85, 37–45. [Google Scholar] [CrossRef]
- Rashki, M.; Kharazi-pakdel, A.; Allahyari, H.; van Alphen, J.J.M. Interactions among the entomopathogenic fungus, Beauveria bassiana (Ascomycota: Hypocreales), the parasitoid, Aphidius matricariae (Hymenoptera: Braconidae), and its host, Myzus persicae (Homoptera: Aphididae). Biol. Control 2009, 50, 324–328. [Google Scholar] [CrossRef]
- Bayındır Erol, A.; Abdelaziz, O.; Birgücü, A.K.; Senoussi, M.M.; Oufroukh, A.; Karaca, İ. Effects of some entomopathogenic fungi on the aphid species, Aphis gossypii Glover (Hemiptera: Aphididae). Egypt. J. Biol. Pest Control 2020, 30, 3–6. [Google Scholar] [CrossRef]
- Sahayaraj, K.; Namachivayam, S. Field evaluation of three entomopathogenic fungi on groundnut pests. Tropicultura 2011, 29, 143–147. [Google Scholar]
- Kaushal, K.S.; Ajoy, K.C.; Priyanka, K. Entomopathogenic Fungi. In Ecofriendly Pest Management for Food Security; Omkar, Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 475–505. [Google Scholar] [CrossRef]
- Toledo, A.V.; de Remes Lenicov, A.M.M.; López Lastra, C.C. Pathogenicity of fungal isolates (Ascomycota: Hypocreales) against Peregrinus maidis, Delphacodes kuscheli (Hemiptera: Delphacidae), and Dalbulus maidis (Hemiptera: Cicadellidae), vectors of corn diseases. Mycopathologia 2007, 163, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Bawin, T.; Seye, F.; Boukraa, S.; Zimmer, J.Y.; Raharimalala, F.N.; Ndiaye, M.; Compere, P.; Delvigne, F.; Francis, F. Histopathological effects of Aspergillus clavatus (Ascomycota: Trichocomaceae) on larvae of the southern house mosquito, Culex quinquefasciatus (Diptera: Culicidae). Fungal Biol. 2016, 120, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Gurulingappa, P.; Sword, G.A.; Murdoch, G.; McGee, P.A. Colonization of crop plants by fungal entomopathogens and their effects on two insect pests when in planta. Biol. Control 2010, 55, 34–41. [Google Scholar] [CrossRef]
- Jallow, M.F.A.; Dugassa-Gobena, D.; Vidal, S. Influence of an endophytic fungus on host plant selection by a polyphagous moth via volatile spectrum changes. Arthropod. Plant. Interact. 2008, 2, 53–62. [Google Scholar] [CrossRef]
- Reddy, N.P.; Ali Khan, A.P.; Devi, U.K.; Sharma, H.C.; Reineke, A. Treatment of millet crop plant (Sorghum bicolor) with the entomopathogenic fungus (Beauveria bassiana) to combat infestation by the stem borer, Chilo partellus Swinhoe (Lepidoptera: Pyralidae). J. Asia Pac. Entomol. 2009, 12, 221–226. [Google Scholar] [CrossRef][Green Version]
- Quesada-Moraga, E.; Munoz-Ledesma, F.J.; Santiago-Alvarez, C. Systemic protection of Papaver somniferum L. against Iraella luteipes (Hymenoptera: Cynipidae) by an endophytic strain of Beauveria bassiana (Ascomycota: Hypocreales). Environ. Entomol. 2009, 38, 723–730. [Google Scholar] [CrossRef]
- Akutse, K.S.; Maniania, N.K.; Fiaboe, K.K.M.; Berg, J.V.A.N.D.E.N.; Ekesi, S. Endophytic colonization of Vicia faba and Phaseolus vulgaris (Fabaceae) by fungal pathogens and their effects on the life- history parameters of Liriomyza huidobrensis (Diptera: Agromyzidae). Fungal Ecol. 2013, 6, 293–301. [Google Scholar] [CrossRef]
- Powell, W.A.; Klingeman, W.E.; Ownley, B.H.; Gwinn, K.D. Evidence of endophytic Beauveria bassiana in seed-treated tomato plants acting as a systemic entomopathogen to larval Helicoverpa zea (Lepidoptera: Noctuidae). J. Entomol. Sci. 2009, 44, 391–396. [Google Scholar] [CrossRef]
- Wagner, B.L.; Lewis, L.C. Colonization of corn, Zea mays, by the entomopathogenic fungus Beauveria bassiana. Appl. Environ. Microbiol. 2000, 66, 3468–3473. [Google Scholar] [CrossRef]
- Meister, B.; Krauss, J.; Härri, S.A.; Victoria Schneider, M.; Müller, C.B. Fungal endosymbionts affect aphid population size by reduction of adult life span and fecundity. Basic Appl. Ecol. 2006, 7, 244–252. [Google Scholar] [CrossRef]
- Collinson, N.P.; Mann, R.C.; Giri, K.; Malipatil, M.; Kaur, J.; Spangenberg, G.; Valenzuela, I. Novel bioassay to assess antibiotic effects of fungal endophytes on aphids. PLoS ONE 2020, 15, e0228813. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, F.; Vidal, S.; Rautenbach, F.; Lewu, F.; Nchu, F. Effects of Beauveria bassiana (Hypocreales) on plant growth and secondary metabolites of extracts of hydroponically cultivated chive (Allium schoenoprasum L. [Amaryllidaceae]). Heliyon 2019, 5, e03038. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, G. Review on safety of the entomopathogenic fungi Beauveria bassiana and Beauveria brongniartii. Biocontrol Sci. Technol. 2007, 17, 553–596. [Google Scholar] [CrossRef]
- Liu, H.; Xie, L.; Wang, J.; Guo, Q.; Yang, S.; Liang, P.; Wang, C.; Lin, M.; Xu, Y.; Zhang, L. The stress-responsive and host-oriented role of nonribosomal peptide synthetases in an entomopathogenic fungus, Beauveria bassiana. J. Microbiol. Biotechnol. 2017, 27, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Prince, G.; Chandler, D. Susceptibility of Myzus persicae, Brevicoryne brassicae and Nasonovia ribisnigri to fungal biopesticides in laboratory and field experiments. Insects 2020, 11, 55. [Google Scholar] [CrossRef]
- Jandricic, S.E.; Filotas, M.; Sanderson, J.P.; Wraight, S.P. Pathogenicity of conidia-based preparations of entomopathogenic fungi against the greenhouse pest aphids Myzus persicae, Aphis gossypii, and Aulacorthum solani (Hemiptera: Aphididae). J. Invertebr. Pathol. 2014, 118, 34–46. [Google Scholar] [CrossRef]
- de Oliveira, C.F.; Long, E.Y.; Finke, D.L. A negative effect of a pathogen on its vector? A plant pathogen increases the vulnerability of its vector to attack by natural enemies. Oecologia 2014, 174, 1169–1177. [Google Scholar] [CrossRef][Green Version]
- Mauck, K.E.; De Moraes, C.M.; Mescher, M.C. Biochemical and physiological mechanisms underlying effects of Cucumber mosaic virus on host-plant traits that mediate transmission by aphid vectors. Plant Cell Environ. 2014, 37, 1427–1439. [Google Scholar] [CrossRef]
- Souza, P.F.N.; Garcia-Ruiz, H.; Carvalho, F.E.L. What proteomics can reveal about plant–virus interactions? Photosynthesis-related proteins on the spotligh. Physiol. Behav. 2017, 176, 139–148. [Google Scholar] [CrossRef]
- Mauck, K.E.; De Moraes, C.M.; Mescher, M.C. Effects of pathogens on sensory-mediated interactions between plants and insect vectors. Curr. Opin. Plant Biol. 2016, 32, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Jaber, L.R.; Salem, N.M. Endophytic colonisation of squash by the fungal entomopathogen Beauveria bassiana (Ascomycota: Hypocreales) for managing Zucchini yellow mosaic virus in cucurbits. Biocontrol Sci. Technol. 2014, 24, 1096–1109. [Google Scholar] [CrossRef]
- Muvea, A.M.; Subramanian, S.; Maniania, N.K.; Poehling, H.M.; Ekesi, S.; Meyhöfer, R. Endophytic colonization of onions induces resistance against viruliferous thrips and virus replication. Front. Plant Sci. 2018, 871, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ownley, B.H.; Gwinn, K.D.; Vega, F.E. Endophytic fungal entomopathogens with activity against plant pathogens: Ecology and evolution. BioControl 2010, 55, 113–128. [Google Scholar] [CrossRef]
- Vega, F.E. The use of fungal entomopathogens as endophytes in biological control: A review. Mycologia 2018, 110, 4–30. [Google Scholar] [CrossRef] [PubMed]
- Golo, P.S.; Gardner, D.R.; Grilley, M.M.; Takemoto, J.Y.; Krasnoff, S.B.; Pires, M.S.; Fernandes, E.K.K.; Bittencourt, V.R.E.P.; Roberts, D.W. Production of destruxins from Metarhizium spp. fungi in artificial medium and in endophytically colonized cowpea plants. PLoS ONE 2014, 9, e104946. [Google Scholar] [CrossRef]
- Loerenstein, G. Localization and induced resistance in virus-infected plants. Annu. Rev. Phytopathol. 1972, 10, 177–206. [Google Scholar] [CrossRef]
- Behie, S.W.; Jones, S.J.; Bidochka, M.J. Plant tissue localization of the endophytic insect pathogenic fungi Metarhizium and Beauveria. Fungal Ecol. 2015, 13, 112–119. [Google Scholar] [CrossRef]
- Quesada-Moraga, E.; Landa, B.B.; Muñoz-Ledesma, J.; Jiménez-Diáz, R.M.; Santiago-Álvarez, C. Endophytic colonisation of opium poppy, Papaver somniferum, by an entomopathogenic Beauveria bassiana strain. Mycopathologia 2006, 161, 323–329. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).