
Distributional Patterns of Aquatic Empididae (Diptera) along an Elevational Diversity Gradient in a Low Mountain Range: An Example from Central Europe


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
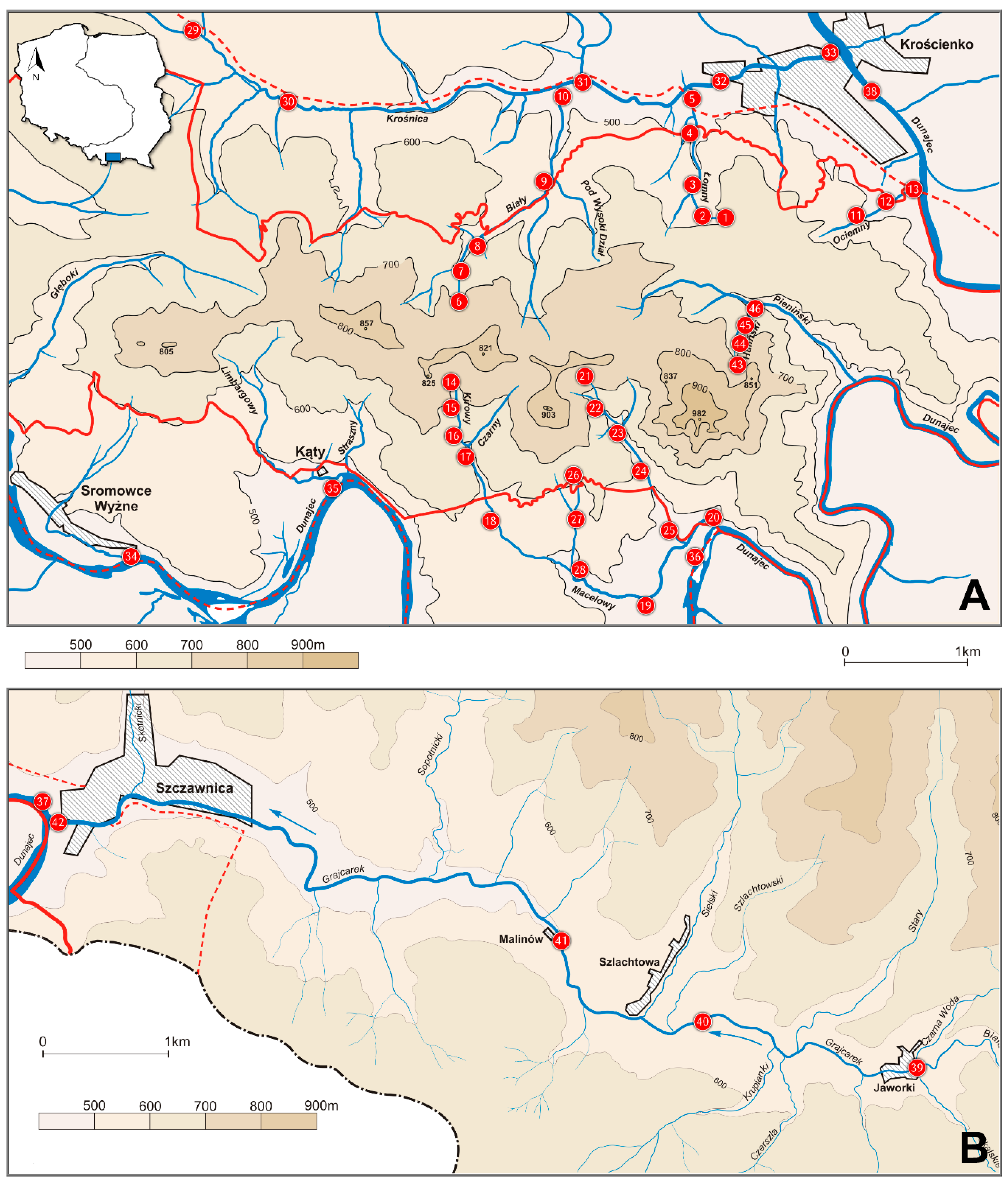
2.1. Study Area and Field Sampling
2.2. Statistical Methods
3. Results
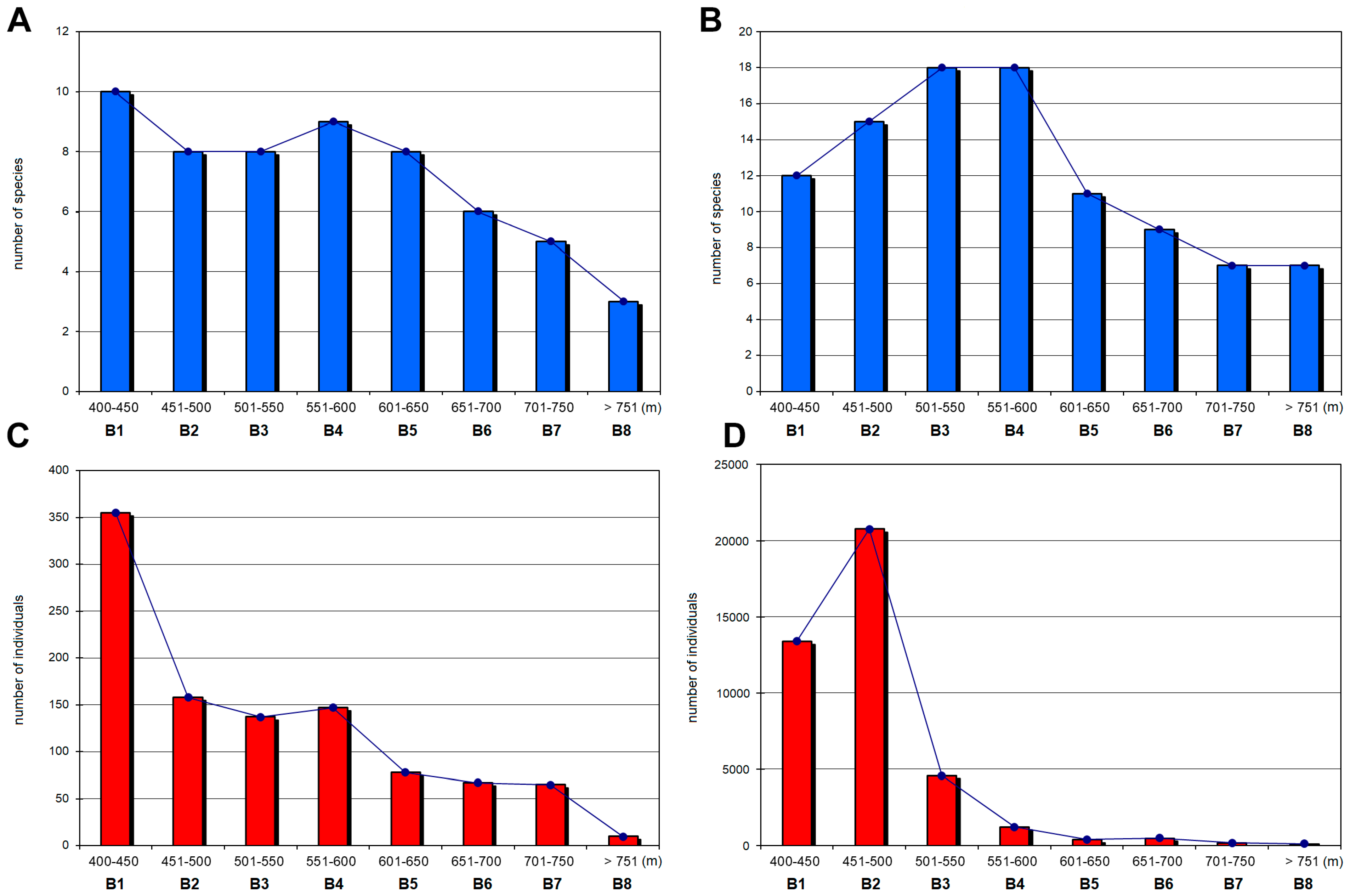
3.1. Species Composition, Species Richness and Diversity along the Elevational Zones
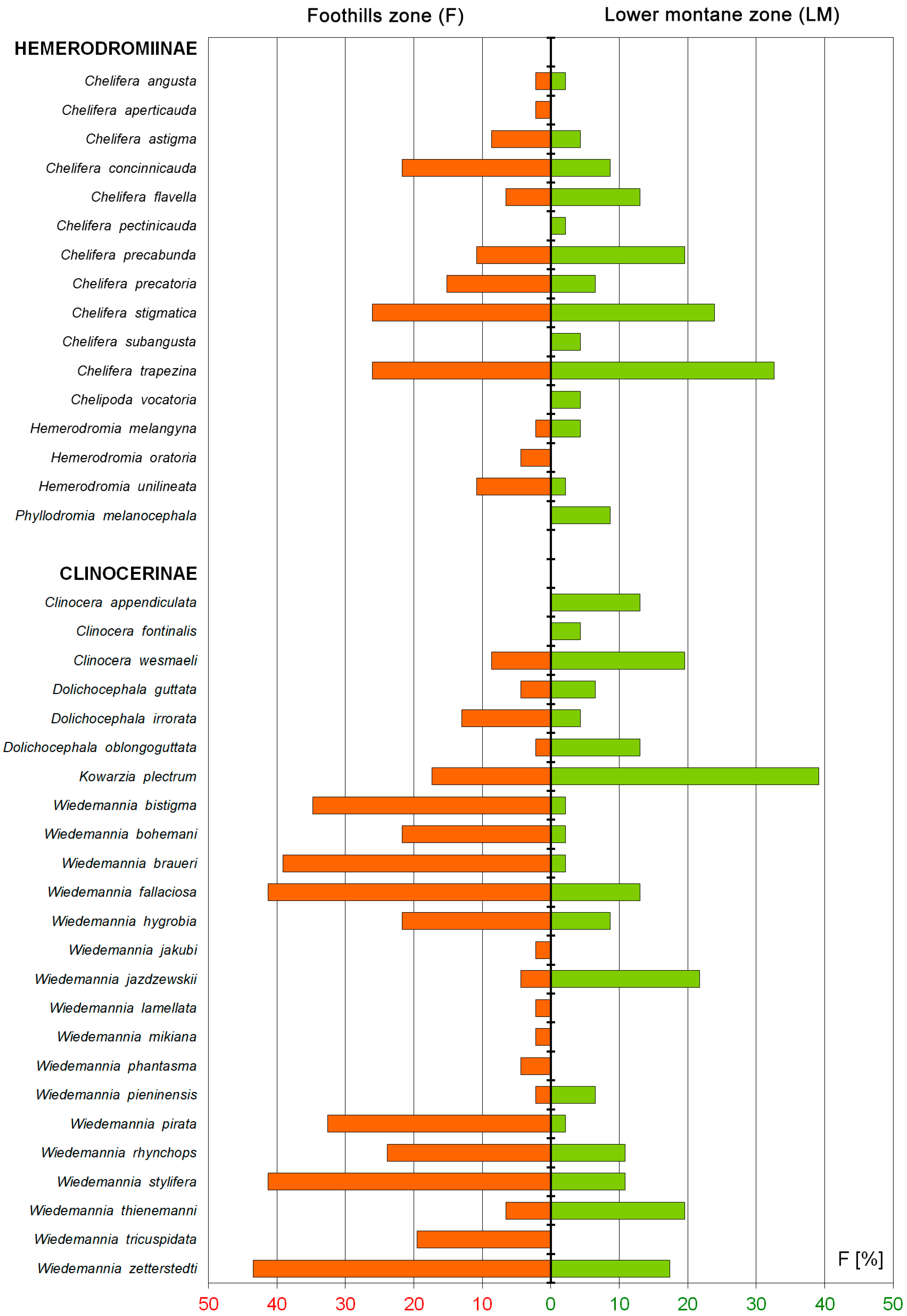
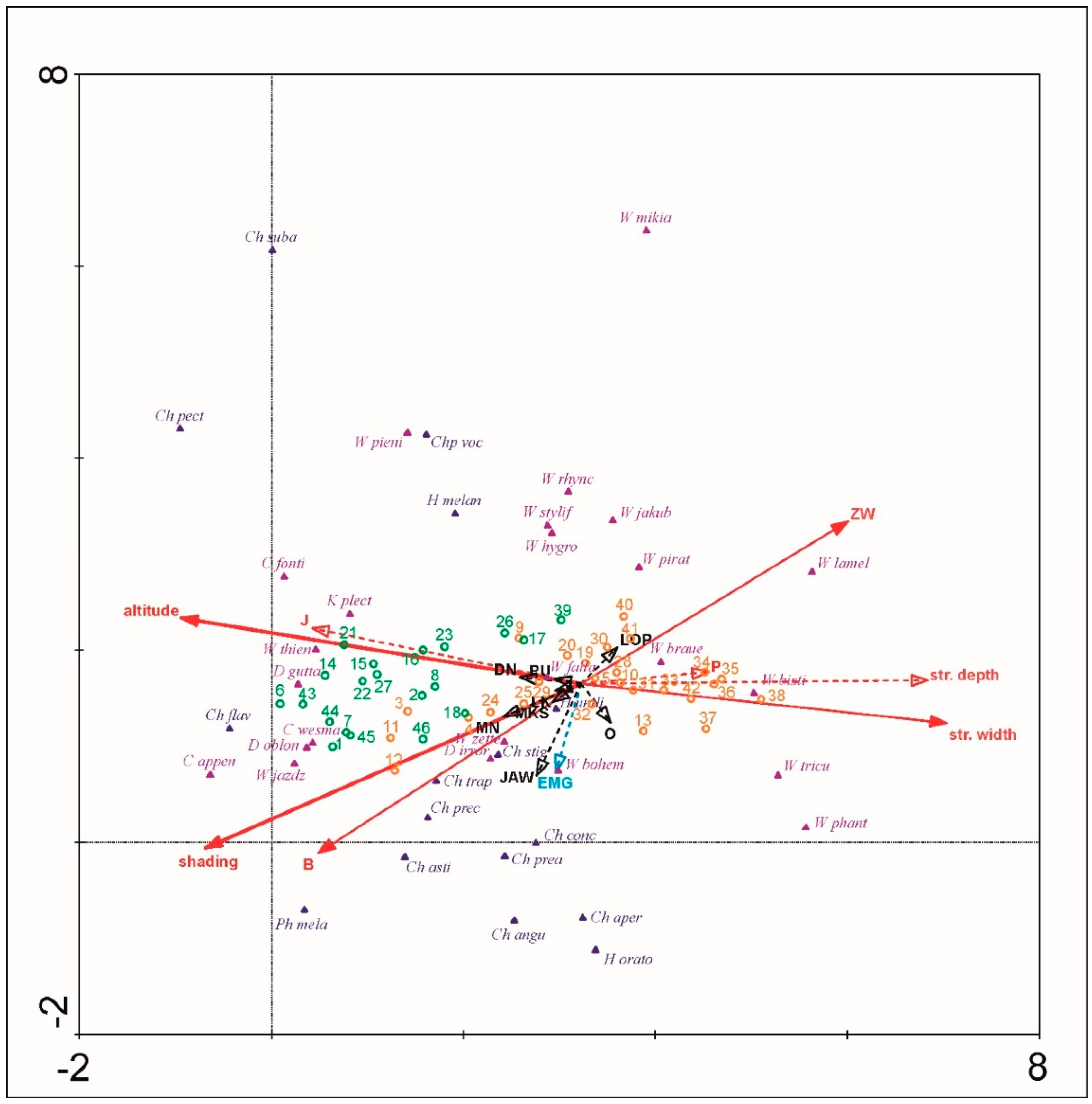
3.2. Biotic and Abiotic Factors vs. Distribution at the Altitudinal Zones
4. Discussion
4.1. Species Richness of the Pieniny Mts. vs. Polish and European Mountain Massifs
4.2. Elevational Patterns of Hemerodromiinae and Clinocerinae
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rahbek, C. The elevational gradient of species richness: A uniform pattern? Ecography 1995, 18, 200–205. [Google Scholar] [CrossRef]
- Rahbek, C. The relationship among area, elevation, and regional species richness in neotropical birds. Am. Nat. 1997, 149, 875–902. [Google Scholar] [CrossRef]
- Terborgh, J. Bird species diversity on an Andean elevational gradient. Ecology 1977, 58, 1007–1019. [Google Scholar] [CrossRef]
- Stevens, G.C. The elevational gradient in altitudinal range: An extension of Rapoport’s latitudinal rule to altitude. Am. Nat. 1992, 140, 893–911. [Google Scholar] [CrossRef]
- Kesller, M. Elevational gradients in species richness and endemism of selected plant groups in the central Bolivian Andes. Plant Ecol. 2000, 149, 181–193. [Google Scholar] [CrossRef]
- Jones, J.I.; Li, W.; Marbely, S. Area, altitude and Aquatic plant diversity. Ecography 2003, 26, 411–420. [Google Scholar] [CrossRef]
- Mallet-Rodrigues, F.; Parrini, R.; Pimentel, L.M.S.; Bessa, R. Altitudinal distribution of birds in a mountainous region in southeastern Brazil. Zoologia 2010, 27, 503–522. [Google Scholar] [CrossRef]
- Bhatt, D.; Joshi, K.K. Bird assemblages in natural and urbanized habitats along elevational gradient in Nainital district (western Himalaya) of Uttarakhand state, India. Curr. Zool. 2011, 57, 318–329. [Google Scholar] [CrossRef]
- Hu, Y.; Jin, K.; Huang, Z.; Ding, Z.; Liang, J.; Pan, X.; Hu, H.; Jiang, Z. Elevational patterns of non-volant small mammal species richness in Gyirong Valley, Central Himalaya: Evaluating multiple spatial and environmental drivers. J. Biogeogr. 2017, 44, 2764–2777. [Google Scholar] [CrossRef]
- Gebrehiwot, K.; Demissew, S.; Woldu, Z.; Fekadu, M.; Desalegn, T.; Teferi, E. Elevational changes in vascular plants richness, diversity, and distribution pattern in Abune Yosef mountain range, Northern Ethiopia. Plant Divers. 2019, 41, 220–228. [Google Scholar] [CrossRef]
- McCoy, E.D. The distribution of insects along elevational gradients. Oikos 1990, 58, 313–332. [Google Scholar] [CrossRef]
- Colwell, R.K.; Hurtt, G.C. Nonbiological gradients in species richness and a spurious Rapoport effect. Am. Nat. 1994, 144, 570–595. [Google Scholar] [CrossRef]
- Olson, D.M. The distribution of leaf litter invertebrates along a Neotropical altitudinal gradient. J. Trop. Ecol. 1994, 10, 129–150. [Google Scholar] [CrossRef]
- Colwell, R.K.; Less, D.C. The mid-domain effect: Geometric constraints on the geography of species richness. Trends Ecol. Evol. 2000, 15, 70–76. [Google Scholar] [CrossRef]
- Colwell, R.K.; Rahbek, C.; Gotelli, N.J. The mid-domain effect and species richness patterns: What have we learned so far? Am. Nat. 2004, 163, E1–E23. [Google Scholar] [CrossRef] [PubMed]
- Colwell, R.K.; Rahbek, C.; Gotelli, N.J. The mid-domain effect: There’s a baby in the bathwater. Am. Nat. 2005, 166, E149–E154. [Google Scholar] [CrossRef][Green Version]
- Rahbek, C. The role of spatial scale and the perception of large-scale species richness patterns. Ecol. Lett. 2005, 8, 224–239. [Google Scholar] [CrossRef]
- Brehm, G.; Colwell, R.K.; Kluge, J. The role of environment and mid-domain effect on moth species richness along a tropical elevational gradient. Glob. Ecol. Biogeogr. 2007, 16, 205–219. [Google Scholar] [CrossRef]
- Choi, S.W.; Am, J.S. Altitudinal distribution of moths (Lepidoptera) in Mt. Jirisan National Park, South Korea. Eur. J. Entomol. 2010, 107, 229–245. [Google Scholar] [CrossRef]
- Herzog, S.K.; Hamel-Leigue, A.C.; Larsen, T.H.; Mann, D.J.; Soria-Auza, R.W.; Gill, B.D.; Edmonds, W.D.; Spector, S. Elevational Distribution and Conservation Biogeography of Phanaeine Dung Beetles (Coleoptera: Scarabaeinae) in Bolivia. PLoS ONE 2013, 8, e64963. [Google Scholar] [CrossRef]
- Le Cesne, M.; Wilson, S.W.; Soulier-Perkins, A. Elevational gradient of Hemiptera (Heteroptera, Auchenorrhyncha) on a tropicalmountain in Papua New Guinea. PeerJ 2015, 3, e978. [Google Scholar] [CrossRef] [PubMed]
- Widhiono, I.; Sudiana, E.; Darsono, D. Diversity of wild bees along elevational gradient in an agricultural area in central Java, Indonesia. Psyche A J. Entomol. 2017, 2017, 2968414. [Google Scholar] [CrossRef]
- Longino, J.T.; Branstetter, M.G. The truncated bell: An enigmatic but pervasive elevational diversity pattern in Middle American ants. Ecography 2018, 41, 1–12. [Google Scholar] [CrossRef]
- Subedi, I.P.; Budha, P.B. Diversity and distribution patterns of ants along elevational gradients. Nepal. J. Zool. 2020, 4, 44–49. [Google Scholar] [CrossRef]
- Sanchez-Rodriguez, J.F.; Baz, A. The effects of elevation on the butterfly communities of a Mediterranean mountain, Sierra de Javalambre, central Spain. J. Lepid. Soc. 1995, 49, 192–207. [Google Scholar]
- Sanders, N.J. Elevational gradients in ant species richness: Area, geometry, and Rapoport’s rule. Ecography 2002, 25, 25–32. [Google Scholar] [CrossRef]
- Sanders, N.J.; Moss, J.; Wagner, D. Patterns of ant species richness along elevational gradients in an arid ecosystem. Glob. Ecol. Biogeogr. 2003, 12, 93–102. [Google Scholar] [CrossRef]
- Beck, J.; Altermatt, F.; Hagmann, R.; Lang, S. Seasonality in the altitude–diversity pattern of Alpine moths. Basic Appl. Ecol. 2010, 11, 714–722. [Google Scholar] [CrossRef]
- Maveety, S.A.; Browne, R.A.; Erwin, T.L. Carabidae diversity along an altitudinal gradient in a Peruvian cloud forest (Coleoptera). ZooKeys 2011, 147, 651–666. [Google Scholar] [CrossRef]
- Bharti, H.; Dharma, Y.P.; Bharti, M.; Pfeiffer, M. Ant species richness, endemicity and functional groups, along an elevational gradient in the Himalayas. Asian Myrmecol. 2013, 5, 79–101. [Google Scholar]
- Yu, X.-D.; Lü, L.; Luo, T.-H.; Zhou, H.-Z. Elevational Gradient in Species Richness Pattern of Epigaeic Beetles and Underlying Mechanisms at East Slope of Balang Mountain in Southwestern China. PLoS ONE 2013, 8, e69177. [Google Scholar] [CrossRef] [PubMed]
- Pizzolotto, R.; Gobbi, M.; Brandmayr, P. Changes in ground beetle assemblages above and below the treeline of the Dolomites after almost 30 years (1980/2009). Ecol. Evol. 2014, 4, 1284–1294. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Reyes, U.J.; Niño-Maldonado, S.; Jones, R.W. Diversity and altitudinal distribution of Chrysomelidae (Coleoptera) in Peregrina Canyon, Tamaulipas, Mexico. ZooKeys 2014, 417, 103–132. [Google Scholar]
- Pizzolotto, R.; Albertini, A.; Gobbi, M.; Brandmayr, P. Habitat diversity analysis along an altitudinal sequence of alpine habitats: The Carabid beetle assemblages as a study model. Period. Biol. 2016, 118, 241–254. [Google Scholar] [CrossRef]
- Scalercio, S.; Bonacci, T.; Mazzei, A.; Pizzolotto, R.; Brandmayr, P. Better up, worse down: Bidirectional consequences of three decades of climate change on a relict population of Erebia cassioides. J. Insect Conserv. 2014, 18, 643–650. [Google Scholar] [CrossRef]
- Chamberlain, D.; Gobbi, M.; Negro, M.; Caprio, E.; Palestrini, C.; Pedrotti, L.; Brandmayr, P.; Pizzolotto, R.; Rolando, A. Trait-modulated decline of carabid beetle occurrence along elevational gradients across the European Alps. J. Biogeogr. 2020, 47, 1030–1040. [Google Scholar] [CrossRef]
- Baz, A.; Cifrián, B.; María Díaz-Aranda, L.; Martín-Vega, M. The distribution of adult blow-flies (Diptera: Calliphoridae) along an altitudinal gradient in Central Spain. Ann. Soc. Entomol. Fr. 2007, 43, 289–296. [Google Scholar] [CrossRef]
- Hackenberger, B.K.; Jarić, D.; Krčmar, S. Distribution of Tabanids (Diptera: Tabanidae) Along a Two-Sided Altitudinal Transect. Environ. Entomol. 2009, 38, 1600–1607. [Google Scholar] [CrossRef]
- Lambkin, C.L.; Boulter, S.L.; Starick, N.T.; Cantrell, B.K.; Bickel, D.J.; Wright, S.G.; Power, N.; Schutze, M.K.; Turco, F.; Nakamura, A.; et al. Altitudinal and seasonal variation in the family-level assemblages of flies (Diptera) in Australian subtropical rainforest: One hundred thousand and counting! Mem. Qld. Mus. Nat. 2011, 55, 315–331. [Google Scholar]
- Mujica, N.; Kroschel, J.; Zebitz, C. Latitudinal and altitudinal distribution and relative importance of the leafminer flies Liriomyza huidobrensis and L. sativae (Diptera: Agromyzidae) in low and highland regions of Peru. Mitt. Dtsch. Ges. Allg. Angew. Entomol. 2012, 18, 297–300. [Google Scholar]
- Rohner, P.T.; Bächli, G.; Pollini Paltrinieri, L.; Duelli, P.; Obrist, M.K.; Jochmann, R.; Blanckenhorn, W.U. Distribution, diversity gradients and Rapoport’s elevational rule in the black scavenger flies of the Swiss Alps (Diptera: Sepsidae). Insect Conserv. Divers. 2015, 8, 367–376. [Google Scholar] [CrossRef]
- Sengupta, J.; Naskar, A.; Maity, A.; Banerjee, D. Effects of selected environmental variable upon the distribution of hover fly (Insecta: Diptera: Syrphidae) along with an altitudinal gradient. Int. J. Adv. Life Sci. Res. 2018, 1, 1–6. [Google Scholar] [CrossRef][Green Version]
- Lamontagne, S.; Donald, D.B.; Schindler, D.W. The distribution of four Chaohorus species (Diptera: Chaoboridae) along an elevation gradient in Canadian Rocky Mountain lakes. Can. J. Zool. 1994, 72, 1531–1537. [Google Scholar] [CrossRef]
- Denke, P.M.; Lloyd, J.E.; Littlefield, J.L. Elevational distribution of mosquitoes in a mountainous area of southeastern Wyoming. J. Am. Mosq. Control Assoc. 1996, 12, 8–16. [Google Scholar]
- Frutiger, A.; Gammeter, S. Faunistics and altitudinal distribution of net-winged midges (Diptera: Blephariceridae) in Switzerland and Liechtenstein. Mitteilungen der Schweizerischen Entomologischen Gesellschaft. Bull. Soc. Entomol. Suisse 1998, 71, 115–124. [Google Scholar]
- Scheibler, E.E.; Roig-Juñent, S.A.; Claps, M.C. Chironomid (Insecta: Diptera) assemblages along an Andean altitudinal gradient. Aquat. Biol. 2014, 20, 169–184. [Google Scholar] [CrossRef]
- Matthews-Bird, F.; Gosling, W.D.; Coe, A.L.; Bush, M.; Mayle, F.E.; Axford, Y.; Brooks, S.J. Environmental controls on the distribution and diversity of lentic Chironomidae (Insecta: Diptera) across an altitudinal gradient in tropical South America. Ecol. Evol. 2016, 6, 91–112. [Google Scholar] [CrossRef]
- Mantilla, J.S.; Moncada, L.I.; Matta, N.E.; Adler, P.H. Distribution of black flies (Diptera: Simuliidae) along an elevational gradient in the Andes Mountains of Colombia during the El Niño Southern Oscillation. Acta Trop. 2018, 183, 162–172. [Google Scholar] [CrossRef]
- Khairul Husna, N.; Noor Nasuha, A.A.; Saiyid Jalaluddin, S.S.; Muhammad Luqman Hakim, A.; Nurul Fatihah, A.L.; Idris, A.B.; Siti Khairiyah, M.H. Diversity and abundance of dipteran species at two different elevations in Gunung Datuk, Negeri Sembilan, Malaysia. Serangga 2018, 23, 194–202. [Google Scholar]
- Ivković, M.; Plant, A. Aquatic insects in the Dinarides: Identifying hotspots of endemism and species richness shaped by geological and hydrological history using Empididae (Diptera). Insect Conserv. Divers. 2015, 8, 302–312. [Google Scholar] [CrossRef]
- Plant, A.R. Diversity of Chelipoda Macquart, 1823 (Diptera: Empididae: Hemerodromiinae) in northern Thailand with discussion of a biodiversity ‘hotspot’ at Doi Inthanon. Raffles Bull. Zool. 2009, 57, 255–277. [Google Scholar]
- Plant, A.R.; Surin, C.; Saokhod, R.; Srisuka, W. Elevational Gradients of Diversity and Species Composition of Hemerodromiinae (Diptera: Empididae) at Doi Inthanon, Thailand: Has Historical Partitioning between Seasonally Dry Lowland and Aseasonal Moist Mountain Forests Contributed to the Biodiversity of Southeast Asia? Trop. Nat. Hist. 2012, 12, 9–20. [Google Scholar]
- Plant, A.R.; Bickel, D.J.; Chatelain, P.; Hauser, M.; Le Cesne, M.; Surin, C.; Saokhod, R.; Nama, S.; Soulier-Perkins, A.; Daugeron, C.; et al. Spatiotemporal dynamics of insect diversity in tropical seasonal forests is linked to season and elevation, a case from northern Thailand. Raffles Bull. Zool. 2018, 66, 382–393. [Google Scholar]
- Plant, A.R.; Bickel, D.J.; Chatelain, P.; Daugeron, C.; Srisuka, W. Composition and organization of highly speciose Empidoidea (Diptera) communities in tropical montane forests of northern Thailand. Zootaxa 2019, 4590, 1–39. [Google Scholar] [CrossRef]
- Plant, A.R.; Bickel, D.J.; Chatelain, P.; Daugeron, C.; Srisuka, W. Endemism, Similarity and Difference in Montane Evergreen Forest Biodiversity Hotspots: Comparing Communities of Empidoidea (Insecta: Diptera) in the Summit Zones of Doi Inthanon and Doi Phahompok, Thailand. Trop. Nat. Hist. 2020, 20, 16–27. [Google Scholar]
- Chatelain, P.; Plant, A.; Soulier, A.; Daugeron, C. Diversity increases with elevation: Empidine dance flies (Diptera, Empididae) challenge a predominant pattern. Biotropica 2018, 50, 633–640. [Google Scholar] [CrossRef]
- Niesiołowski, S. Morfologia, Biologia i Występowanie w Polsce Wodnych Empididae (Diptera, Brachycera); Acta Universitatis Lodzensis, Folia limnologica: Łódź, Polska, 1990; pp. 1–169. [Google Scholar]
- Krysiak, I.; Niesiołowski, S. Wiedemannia pieninensis a New Species of Aquatic Empidid (Diptera, Empididae) from Poland. Aquat. Insects 2004, 26, 143–146. [Google Scholar] [CrossRef]
- Krysiak, I. Wiedemannia jakubi a New Species of Aquatic Empidid (Diptera: Empididae) from Poland. Ann. Zool. 2005, 55, 75–76. [Google Scholar]
- Krysiak, I.; Palaczyk, A.; Wanat, D. Empidoidea (Diptera: Brachycera). In Owady (Insecta) Parku Krajobrazowego Wzniesień Łódzkich; Jaskuła, R., Tończyk, G., Eds.; Dyrekcja Parku Krajobrazowego Wzniesień Łódzkich/Mazowiecko-Świętokrzyskie Towarzystwo Ornitologiczne: Łódź, Polska, 2010; pp. 159–163. [Google Scholar]
- Słowińska-Krysiak, I. New Records of an Extremely Rare Aquatic Empidid Fly Chelifera polonica Wagner and Niesiolowski, 1987 (Diptera: Empididae: Hemerodromiinae) from Poland. J. Entomol. Res. Soc. 2012, 14, 99–103. [Google Scholar]
- Słowińska-Krysiak, I. Chelifera pectinicauda COLLIN, 1927 i Chelifera subangusta COLLIN, 1961—nowe dla Bieszczadów gatunki Hemerodromiinae (Diptera: Empididae). Wiad. Entomol. 2013, 32, 147–150. [Google Scholar]
- Słowińska-Krysiak, I. New Distributional Data for the Rare Polish Empidid Flies (Empididae: Hemerodromiinae, Clinocerinae) in the Pieniny Mountains with Notes on Ecology and Phenology. J. Entomol. Res. Soc. 2014, 16, 127–140. [Google Scholar]
- Słowińska-Krysiak, I. Nowe stanowisko Wiedemannia pieninensis Krysiak et Niesiołowski, 2004 (Empididae: Clinocerinae) w Pieninach. Pienininy Przyroda i Człowiek 2014, 13, 63–65. [Google Scholar]
- Słowińska, I. New records of the rare species Chelifera aperticauda Collin, 1927 (Diptera: Empididae: Hemerodromiinae) from Poland. Dipteron 2017, 33, 121–126. [Google Scholar]
- Słowińska, I. Hemerodromiinae i Clinocerinae (Diptera: Empididae) Małych Pienin. Dipteron 2017, 33, 143–156. [Google Scholar]
- Słowińska, I. First record of the rare species Wiedemannia pieninensis Krysiak et Niesiołowski, 2004 (Diptera: Empididae: Clinocerinae) from the Polish Tatra Mountains. Dipteron 2019, 35, 178–184. [Google Scholar]
- Palaczyk, A.; Słowińska-Krysiak, I. The genus Phaeobalia Mik, 1881 in Poland (Diptera: Empididae: Clinocerinae). Genus 2013, 24, 415–424. [Google Scholar]
- Słowińska-Krysiak, I.; Palaczyk, A. Empidoidea (Diptera: Brachycera) Spalskiego Parku Krajobrazowego. Dipteron 2014, 30, 63–75. [Google Scholar]
- Słowińska, I.; Palaczyk, A. Nowe dane o Hemerodromiinae (Diptera: Empididae) Tatr polskich. Dipteron 2015, 31, 37–49. [Google Scholar]
- Palaczyk, A.; Słowińska, I.; Klasa, A. The Genus Bergenstammia Mik, 1881 (Diptera: Empididae: Clinocerinae) in Poland with Description of Bergenstammia glacialis sp. nov. from the Tatra Mts. Ann. Zool. 2015, 65, 53–64. [Google Scholar] [CrossRef]
- Słowińska, I.; Palaczyk, A. Hemerodromiinae (Diptera: Empididae) Gorców. Dipteron 2016, 32, 67–76. [Google Scholar]
- Oboňa, J.; Baranová, B.; Hrivniak, L.; Kisková, K.; Manko, P.; Słowińska, I. First record of the rare aquatic dance fly Chelifera aperticauda Collin, 1927 (Diptera: Empididae: Hemerodromiinae) from Slovakia. Check List 2016, 12, 1894. [Google Scholar] [CrossRef]
- Słowińska, I.; Zawal, A.; Stryjecki, R.; Michoński, G. First detailed records of water mite larvae (Hydrachnidia: Hydrovolzidae, Hydryphantidae) parasitizing empidid flies (Diptera: Empididae: Clinocerinae). Int. J. Parasitol. Parasites Wildl. 2020, 12, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Ivković, M.; Čorkalo, M.; Tkoč, M. Aquatic dance flies of Czech Republic (Empididae: Clinocerinae and Hemerodromiinae): New records and updated species list. Zootaxa 2020, 4790, 76–90. [Google Scholar] [CrossRef] [PubMed]
- Kondracki, J. Geografia Regionalna Polski; PWN: Warszawa, Poland, 2000; pp. 1–441. [Google Scholar]
- Nyka, J. Pieniny. Przewodnik. In Wydawnictwo Trawers; Wyd. VIII: Latchorzew, Poland, 2000; 304p. [Google Scholar]
- Birkenmajer, K. Aktualne problemy geologiczne Pienin. Pienininy Przyroda i Człowiek 2003, 8, 33–40. [Google Scholar]
- Birkenmajer, K. Geology of the Pieniny Mountains, West Carpathians, Poland. Monogr. Pienin. Mt. 2017, 3, 5–66. [Google Scholar]
- Biesiadka, E. Wodopójki (Hydracarina) Pienin. Fragm. Faun. 1979, 24, 97–173. [Google Scholar] [CrossRef]
- Niemirowski, M. Zarys fizjografii Pienin. Położenie i ukształtowanie. In Przyroda Pienin w Obliczu Zmian. Studia Naturae Seria B; Zarzycki, K., Ed.; Państwowe Wydawnictwo Naukowe: Warszawa-Kraków, Poland, 1982; Volume 30, pp. 17–31. [Google Scholar]
- Kostrakiewicz, L. Hydrografia. In Przyroda Pienin w Obliczu Zmian. Studia Naturae Seria B; Zarzycki, K., Ed.; Państwowe Wydawnictwo Naukowe: Warszawa-Kraków, Poland, 1982; Volume 30, pp. 70–93. [Google Scholar]
- Biesiadka, E. Ogólna charakterystyka faunistyczna środowisk wodnych Pienin. Fragm. Faun. 1979, 24, 283–293. [Google Scholar] [CrossRef]
- Małecka, D.; Humnicki, W. Problemy hydrogeologii i ochrony wód Pienińskiego Parku Narodowego. Pieniny Przyroda iCzłowiek 2002, 7, 49–70. [Google Scholar]
- Witkowski, Z. Fauna Pienińskiego Parku Narodowego, jej zagrożenie i ochrona. Pienininy Przyrida i Człowiek 2003, 8, 71–82. [Google Scholar]
- Razowski, J. Charakterystyka flory i fauny Pienin. In Flora i Fauna Pienin. Monografie Pienińskie; Razowski, J., Ed.; Pieniński Park Narodowy: Krościenko nad Dunajcem, Poland, 2000; Volume 1, pp. 11–21. [Google Scholar]
- Clark, K.R.; Gorley, R.N. PRIMER v5: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2001. [Google Scholar]
- Ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination, Version-4.5; Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Krysiak, I. Muchówki z Podrodzin Hemerodromiinae i Clinocerinae (Diptera, Empididae) Pienin. [Hemerodromiinae and Clinocerinae (Diptera, Empididae) of Pieniny Mountains]. Ph.D. Thesis, University of Łódź, Łódź, Poland, 2005. [Google Scholar]
- Klasa, A.; Palaczyk, A.; Soszyński, B. Muchówki (Diptera) Bieszczadów. Monografie Bieszczadzkie 2000, 8, 305–369. [Google Scholar]
- Palaczyk, A.; Klasa, A. Muchówki (Diptera) masywu Babiej Góry. In Monografia Fauny Babiej Góry; Wołoszyn, B.W., Wołoszyn, D., Celary, W., Eds.; Komitet Ochrony Przyrody Polskiej Akademii Nauk: Kraków, Poland, 2003; pp. 305–357. [Google Scholar]
- Ivković, M.; Zamora-Muñoz, C.; Sainz-Bariaín, M.; Sinclair, B.J. Aquatic Empididae (Diptera: Hemerodromiinae and Clinocerinae) of the Sierra Nevada, Spain, with the description of five new species. Zootaxa 2014, 3786, 541–556. [Google Scholar] [CrossRef]
- Joost, W. Hemerodromia illiesi sp. n. aus Georgien (UdSSR) (Diptera, Empididae). Reichenbachia 1980, 18, 81–83. [Google Scholar]
- Joost, W. Beitrag zur Kenntnis der Hemerodromiinae des Kaukasus (I) (Diptera, Empididae). Reichenbachia 1981, 19, 183–191. [Google Scholar]
- Joost, W. Dolichocepahala monae sp. n. aus Armenien (UdSSR) (Diptera, Empididae). Reichenbachia 1981, 19, 193–195. [Google Scholar]
- Sinclair, B.J.; Shamshev, I.V. Review of Clinocerinae (Diptera: Empididae) from the Caucasus, with description of three new species. Proc. Zool. Inst. Ras 2014, 318, 40–47. [Google Scholar]
- Kustov, S.; Zherebilo, D. New data on the genus Wiedemannia Zetterstedt (Diptera: Empididae) from the Caucasus with description of four new species. Zootaxa 2015, 4032, 351–369. [Google Scholar] [CrossRef]
- Vaillant, F. Quelques Insectes Diptères, à larves aquatiques, du Parc de la Vanoise. Trav. Sci. Parc. Natl. Vanoise 1973, 3, 133–165. [Google Scholar]
- Wolda, H. Altitude, habitat and tropical insect diversity. Biol. J. Linn. Soc. 1987, 30, 313–323. [Google Scholar] [CrossRef]
- Pawłowski, J. Chrząszcze (Coleoptera) Babiej Góry. Acta Zool. Crac. 1967, 2, 419–665. [Google Scholar]
- Mielewczyk, S. Ważki (Odonata) Pienin. Fragm. Faun. 1978, 22, 265–294. [Google Scholar]
- Mielewczyk, S. Pluskwiaki wodne (Heteroptera aquatica et semiaquatica) Pienin. Fragm. Faun. 1978, 22, 295–336. [Google Scholar]
- Dyduch-Falniowska, A. The Gastropods of the Polish Tatra Mountains; Zakład Ochrony Przyrody i Zasobów Naturalnych PAN: Kraków, Poland, 1991; pp. 1–111. [Google Scholar]
- Szczęsny, B.; Wiśniowska, A. Fauna jętek (Ephemeroptera) Babiej Góry. In Monografia Fauny Babiej Góry; Wołoszyn, B.W., Wołoszyn, D., Celary, W., Eds.; Komitet Ochrony Przyrody Polskiej Akademii Nauk: Kraków, Poland, 2003; pp. 123–139. [Google Scholar]
- Szczęsny, B.; Wiśniowska, A. Fauna widelnic (Plecoptera) Babiej Góry. In Monografia Fauny Babiej Góry; Wołoszyn, B.W., Wołoszyn, D., Celary, W., Eds.; Komitet Ochrony Przyrody Polskiej Akademii Nauk: Kraków, Poland, 2003; pp. 141–158. [Google Scholar]
- Plant, A.R. Diversity, distribution and community composition of Empididae, Hybotidae and Brachystomatidae (Insecta: Diptera) in Thailand—How did they arise? Res. Knowl. 2015, 1, 1–14. [Google Scholar]
- MacArthur, R.H. Environmental factors affecting bird species diversity. Am. Nat. 1964, 98, 387–397. [Google Scholar]
- MacArthur, R.H. Patterns of species diversity. Biol. Rev. 1965, 40, 510–533. [Google Scholar] [CrossRef]
- Rosenzweig, M.L.; Winakur, J. Population ecology of desert rodent communities: Habitats and environmental complexity. Ecology 1969, 50, 558–572. [Google Scholar] [CrossRef]
- Allan, D.J. The distributional ecology and diversity of benthic insects in Cement Creek, Colorado. Ecology 1975, 56, 1040–1053. [Google Scholar] [CrossRef]
- Vaillant, F. La répartition des Wiedemannia dans les cours d’eau et leur utilisation comme inidicateurs de zones écologiques (Diptera, Empididae). Ann. Limnol. 1967, 3, 267–293. [Google Scholar] [CrossRef][Green Version]
- Vaillant, F. Quelques Empididae Hemerodromiinae des Pyrénées (Diptera). Ann. Limnol. 1968, 4, 85–93. [Google Scholar] [CrossRef][Green Version]
- Vaillant, F. Diptères Empididae Hemerodromiinae nouveaux ou peu connus de la région paléartique (première partie). Bonn. Zool. Beiträge 1981, 32, 351–408. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Sampling Site | Altitude (m) | GPS Location |
|---|---|---|---|
| 1 | Łonny stream, headspring | 620 | 49°25′46″ N 20°24′56″ E |
| 2 | Łonny stream | 560 | 49°25′51″ N 20°24′46″ E |
| 3 | Łonny stream | 520 | 49°25′53″ N 20°24′44″ E |
| 4 | Łonny stream | 463 | 49°26′11″ N 20°24′45″ E |
| 5 | Łonny stream (outlet to the Krośnica stream) | 440 | 49°26′22″ N 20°24′43″ E |
| 6 | Biały stream, headspring | 750 | 49°25′12″ N 20°23′09″ E |
| 7 | Biały stream | 640 | 49°25′31″ N 20°23′09″ E |
| 8 | Biały stream | 560 | 49°25′38″ N 20°23′14″ E |
| 9 | Biały stream | 515 | 49°25′52″ N 20°23′43″ E |
| 10 | Biały stream (outlet to the Krośnica stream) | 462 | 49°26′18″ N 20°23′45″ E |
| 11 | Ociemny stream, headspring | 510 | 49°25′48″ N 20°25′55″ E |
| 12 | Ociemny stream | 480 | 49°25′49″ N 20°25′59″ E |
| 13 | Ociemny stream (outlet to the Dunajec River) | 445 | 49°25′54″ N 20°26′19″ E |
| 14 | Kirowy stream, headspring | 770 | 49°25′02″ N 20°23′10″ E |
| 15 | Kirowy stream | 680 | 49°24′51″ N 20°23′09″ E |
| 16 | Kirowy stream | 610 | 49°24′45″ N 20°23′10″ E |
| 17 | Macelowy stream | 587 | 49°24′38″ N 20°23′15″ E |
| 18 | Macelowy stream | 560 | 49°24′35″ N 20°23′17″ E |
| 19 | Macelowy stream, Sromowce Niżne village | 460 | 49°24′04″ N 20°24′35″ E |
| 20 | Macelowy stream (outlet to the Dunajec River) | 450 | 49°24′22″ N 20°25′01″ E |
| 21 | Sobczański stream, headspring | 750 | 49°25′01″ N 20°24′08″ E |
| 22 | Sobczański stream | 655 | 49°24′55″ N 20°24′09″ E |
| 23 | Sobczański stream | 590 | 49°24′50″ N 20°24′16″ E |
| 24 | Sobczański stream | 530 | 49°24′38″ N 20°24′31″ E |
| 25 | Sobczański stream, (outlet to the Macelowy stream), Sromowce Niżne village | 455 | 49°24′19″ N 20°24′45″ E |
| 26 | Kotłowy stream, headspring | 610 | 49°24′39″ N 20°24′02″ E |
| 27 | Kotłowy stream | 570 | 49°24′31″ N 20°24′01″ E |
| 28 | Kotłowy stream (outlet to the Macelowy stream) | 490 | 49°24′10″ N 20°23′59″ E |
| 29 | Krośnica stream, Krośnica village | 550 | 49°26′41″ N 20°20′43″ E |
| 30 | Krośnica stream, near Hałuszowa village | 520 | 49°26′13″ N 20°21′47″ E |
| 31 | Krośnica stream, Tylka village | 462 | 49°26′20″ N 20°23′45″ E |
| 32 | Krośnica stream, Krościenko village | 440 | 49°26′23″ N 20°24′49″ E |
| 33 | Krośnica stream (outlet to the Dunajec River), Krościenko village | 415 | 49°26′33″ N 20°25′39″ E |
| 34 | Dunajec River, Sromowce Wyżne village | 475 | 49°24′03″ N 20°21′12″ E |
| 35 | Dunajec River, Kąty village | 465 | 49°24′25″ N 20°22′12″ E |
| 36 | Dunajec River, Sromowce Niżne village | 455 | 49°24′22″ N 20°25′01″ E |
| 37 | Dunajec River, Szczawnica | 430 | 49°26′26″ N 20°27′34″ E |
| 38 | Dunajec River, Krościenko village | 420 | 49°26′27″ N 20°25′50″ E |
| 39 | Małe Pieniny, Grajcarek stream, Jaworki village | 580 | 49°24′26″ N 20°33′14″ E |
| 40 | Małe Pieniny, Grajcarek stream, Szlachtowa village | 540 | 49°24′37″ N 20°31′48″ E |
| 41 | Małe Pieniny, Grajcarek stream | 490 | 49°25′03″ N 20°30′47″ E |
| 42 | Małe Pieniny, Grajcarek stream (outlet to the Dunajec River), Szczawnica | 430 | 49°25′27″ N 20°27′35″ E |
| 43 | Huliński stream | 745 | 49°24′56″ N 20°25′01″ E |
| 44 | Huliński stream | 680 | 49°25′VI″ N 20°25′05″ E |
| 45 | Huliński stream | 630 | 49°25′15″ N 20°25′04″ E |
| 46 | Huliński stream (outlet to the Pieniński stream) | 587 | 49°25′25″ N 20°25′13″ E |
| Altitude (m) | Foothills Zone (F) | Lower Montane Zone (LM) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 400–450 (B1) | 451–500 (B2) | 501–550 (B3) | Total | 551–600 (B4) | 601–650 (B5) | 651–700 (B6) | 701–750 (B7) | >751 (B8) | Total | |
| Number of Hemerodromiinae species | 10 | 8 | 8 | 12 | 9 | 8 | 6 | 5 | 3 | 14 |
| Number of Hemerodromiinae individuals | 355 | 158 | 137 | 650 | 147 | 78 | 67 | 65 | 10 | 367 |
| Chelifera angusta Collin, 1927 | 3 | 3 | 1 | 1 | ||||||
| Chelifera aperticauda Collin, 1927 | 1 | 1 | ||||||||
| Chelifera astigma Collin, 1927 | 1 | 11 | 1 | 13 | 3 | 3 | ||||
| Chelifera concinnicauda Collin, 1927 | 184 | 10 | 11 | 205 | 5 | 1 | 6 | |||
| Chelifera flavella (Zetterstedt, 1838) | 1 | 11 | 12 | 10 | 53 | 8 | 71 | |||
| Chelifera pectinicauda Collin, 1927 | 1 | 1 | ||||||||
| Chelifera precabunda Collin, 1961 | 15 | 4 | 6 | 25 | 18 | 5 | 4 | 5 | 32 | |
| Chelifera precatoria (Fallén, 1816) | 7 | 6 | 13 | 26 | 4 | 2 | 6 | |||
| Chelifera stigmatica (Schiner, 1862) | 59 | 59 | 48 | 166 | 41 | 4 | 3 | 48 | ||
| Chelifera subangusta Collin, 1961 | 1 | 1 | 2 | |||||||
| Chelifera trapezina (Zetterstedt, 1838) | 24 | 47 | 46 | 117 | 64 | 30 | 56 | 3 | 1 | 154 |
| Chelipoda vocatoria (Fallén, 1816) | 1 | 1 | 2 | |||||||
| Hemerodromia melangyna Collin, 1927 | 1 | 1 | 1 | 1 | 2 | |||||
| Hemerodromia oratoria (Fallén, 1816) | 21 | 21 | ||||||||
| Hemerodromia unilineata Zetterstedt, 1842 | 40 | 20 | 60 | 1 | 1 | |||||
| Phyllodromia melanocephala (Fabricius, 1794) | 12 | 26 | 38 | |||||||
| Number of Clinocerinae species | 12 | 15 | 18 | 22 | 18 | 11 | 9 | 7 | 7 | 19 |
| Number of Clinocerinae individuals | 13,387 | 20,751 | 4594 | 38,732 | 1224 | 394 | 497 | 185 | 106 | 2406 |
| Clinocera appendiculata (Zetterstedt, 1838) | 1 | 23 | 5 | 30 | 1 | 60 | ||||
| Clinocera fontinalis (Haliday, 1833) | 1 | 1 | 2 | |||||||
| Clinocera wesmaeli (Macquart, 1835) | 2 | 17 | 19 | 23 | 1 | 5 | 2 | 8 | 39 | |
| Dolichocephala guttata (Haliday, 1833) | 2 | 2 | 1 | 2 | 3 | 6 | ||||
| Dolichocephala irrorata (Fallén, 1816) | 4 | 12 | 16 | 2 | 1 | 3 | ||||
| Dolichocephala oblongoguttata (Dale, 1878) | 1 | 1 | 1 | 3 | 1 | 1 | 6 | |||
| Kowarzia plectrum (Mik, 1880) | 67 | 267 | 334 | 256 | 173 | 439 | 137 | 84 | 1089 | |
| Wiedemannia bistigma (Curtis, 1834) | 5932 | 16,146 | 53 | 22,131 | 6 | 6 | ||||
| Wiedemannia bohemani (Zetterstedt, 1838) | 28 | 99 | 28 | 155 | 2 | 2 | ||||
| Wiedemannia braueri (Mik, 1880) | 3657 | 2542 | 1231 | 7430 | 57 | 57 | ||||
| Wiedemannia fallaciosa (Loew, 1873) | 404 | 583 | 1449 | 2436 | 85 | 13 | 98 | |||
| Wiedemannia hygrobia (Loew, 1858) | 10 | 12 | 105 | 127 | 37 | 2 | 39 | |||
| Wiedemannia jakubi Krysiak, 2005 | 1 | 1 | ||||||||
| Wiedemannia jazdzewskii Niesiolowski, 1987 | 21 | 21 | 21 | 159 | 34 | 2 | 4 | 220 | ||
| Wiedemannia lamellata (Loew, 1869) | 35 | 35 | ||||||||
| Wiedemannia mikiana (Bezzi, 1899) | 5 | 5 | ||||||||
| Wiedemannia phantasma (Mik, 1880) | 5 | 5 | ||||||||
| Wiedemannia pieninensis Krysiak et Niesiolowski, 2004 | 4 | 4 | 1 | 4 | 1 | 6 | ||||
| Wiedemannia pirata (Mik, 1880) | 90 | 40 | 347 | 477 | 31 | 31 | ||||
| Wiedemannia rhynchops (Nowicki, 1868) | 4 | 53 | 104 | 161 | 86 | 86 | ||||
| Wiedemannia stylifera Mik, 1889 | 57 | 119 | 691 | 867 | 298 | 298 | ||||
| Wiedemannia thienemanni Wagner, 1982 | 9 | 9 | 18 | 32 | 3 | 6 | 10 | 7 | 58 | |
| Wiedemannia tricuspidata (Bezzi, 1905) | 3084 | 696 | 3780 | |||||||
| Wiedemannia zetterstedti (Fallén, 1816) | 112 | 346 | 249 | 707 | 284 | 15 | 1 | 300 | ||
| Altitude (m) | Foothills Zone (F) | Lower Montane Zone (LM) | ||||||
|---|---|---|---|---|---|---|---|---|
| 400–450 (B1) | 451–500 (B2) | 501–550 (B3) | 551–600 (B4) | 601–650 (B5) | 651–700 (B6) | 701–750 (B7) | >750 (B8) | |
| Hemerodromiinae | ||||||||
| Species richness | 10 | 8 | 8 | 9 | 8 | 6 | 5 | 3 |
| Shannon-Wiener Index (H’) | 1.52 | 1.6 | 1.57 | 1.495 | 1.49 | 0.69 | 0.71 | 0.64 |
| Simpson’s Index (D) | 0.68 | 0.75 | 0.74 | 0.71 | 0.72 | 0.29 | 0.33 | 0.34 |
| Margalef Index (DMg) | 1.53 | 1.38 | 1.42 | 1.6 | 1.61 | 1.19 | 0.96 | 0.87 |
| Pielou Evenness Index (J’) | 0.66 | 0.77 | 0.76 | 0.68 | 0.72 | 0.38 | 0.44 | 0.58 |
| Clinocerinae | ||||||||
| Species richness | 12 | 15 | 18 | 18 | 11 | 9 | 7 | 7 |
| Shannon-Wiener Index (H’) | 1.28 | 0.86 | 1.87 | 2.03 | 1.28 | 0.53 | 0.87 | 0.81 |
| Simpson’s Index (D) | 0.68 | 0.38 | 0.79 | 0.83 | 0.64 | 0.22 | 0.42 | 0.36 |
| Margalef Index (DMg) | 1.16 | 1.41 | 2.02 | 2.39 | 1.67 | 1.29 | 1.15 | 1.29 |
| Pielou Evenness Index (J’) | 0.52 | 0.32 | 0.65 | 0.7 | 0.53 | 0.24 | 0.45 | 0.42 |
| Alps | Pyrenees | Massif Central | Tatra Mts | Pieniny Mts. | |
|---|---|---|---|---|---|
| HEMERODROMIINAE | |||||
| Chelifera astigma | 350–1500 | 970–1100 | 445–750 | ||
| Chelifera flavella | 700–1700 | 970–1500 | 480–770 | ||
| Chelifera pectinicauda | 730 | 800 | 770 | ||
| Chelifera precabunda | 1500–1700 | 910–1520 | 445–750 | ||
| Chelifera precatoria | 700–1100 | 802–1850 | 1200 | 930–1000 | 440–680 |
| Chelifera stigmatica | 250–1000 | ca. 1460 | 960–1050 | 440–680 | |
| Chelifera trapezina | 700–1400 | 1150–1300 | 900–1500 | 440–770 | |
| Hemerodromia unilineata | 400–1000 | 320 | 440–560 | ||
| Hemerodromia oratoria | 720 | 440–445 | |||
| Phyllodromia melanocephala | 850–1000 | 990–1150 | 560–640 | ||
| CLINOCERINAE | |||||
| Clinocera appendiculata | 800–2560 | 1000–2100 | 900–2040 | 590–770 | |
| Clinocera wesmaeli | ca. 1000 | 1000–1300 | 463–770 | ||
| Dolichocephala guttata | 2300 | 650–1500 | 1100 | 463–750 | |
| Dolichocephala irrorata | 650 | 900–1620 | 440–630 | ||
| Kowarzia plectrum | 1900 | 980–1150 | 445–770 | ||
| Wiedemannia bistigma | 215–724 | 115–350 | 200–470 | 415–580 | |
| Wiedemannia bohemani | 215–1000 | 1600 | 700–875 | 920 | 415–560 |
| Wiedemannia fallaciosa | 80–1130 | 1050 | 415–610 | ||
| Wiedemannia hygrobia | 210–1850 | 1350–2150 | 900–1650 | 415–640 | |
| Wiedemannia lamellata | 386 | 465 | |||
| Wiedemannia phantasma | 230–877 | 420–445 | |||
| Wiedemannia zetterstedti | ca. 1000 | 217–1123 | 415–680 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Słowińska, I.; Jaskuła, R. Distributional Patterns of Aquatic Empididae (Diptera) along an Elevational Diversity Gradient in a Low Mountain Range: An Example from Central Europe. Insects 2021, 12, 165. https://doi.org/10.3390/insects12020165
Słowińska I, Jaskuła R. Distributional Patterns of Aquatic Empididae (Diptera) along an Elevational Diversity Gradient in a Low Mountain Range: An Example from Central Europe. Insects. 2021; 12(2):165. https://doi.org/10.3390/insects12020165
Chicago/Turabian StyleSłowińska, Iwona, and Radomir Jaskuła. 2021. "Distributional Patterns of Aquatic Empididae (Diptera) along an Elevational Diversity Gradient in a Low Mountain Range: An Example from Central Europe" Insects 12, no. 2: 165. https://doi.org/10.3390/insects12020165
APA StyleSłowińska, I., & Jaskuła, R. (2021). Distributional Patterns of Aquatic Empididae (Diptera) along an Elevational Diversity Gradient in a Low Mountain Range: An Example from Central Europe. Insects, 12(2), 165. https://doi.org/10.3390/insects12020165