Relicts from Glacial Times: The Ground Beetle Pterostichus adstrictus Eschscholtz, 1823 (Coleoptera: Carabidae) in the Austrian Alps


 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Sampling
2.2. Species Determination and Morphological Comparison
2.3. Molecular Characterization
3. Results
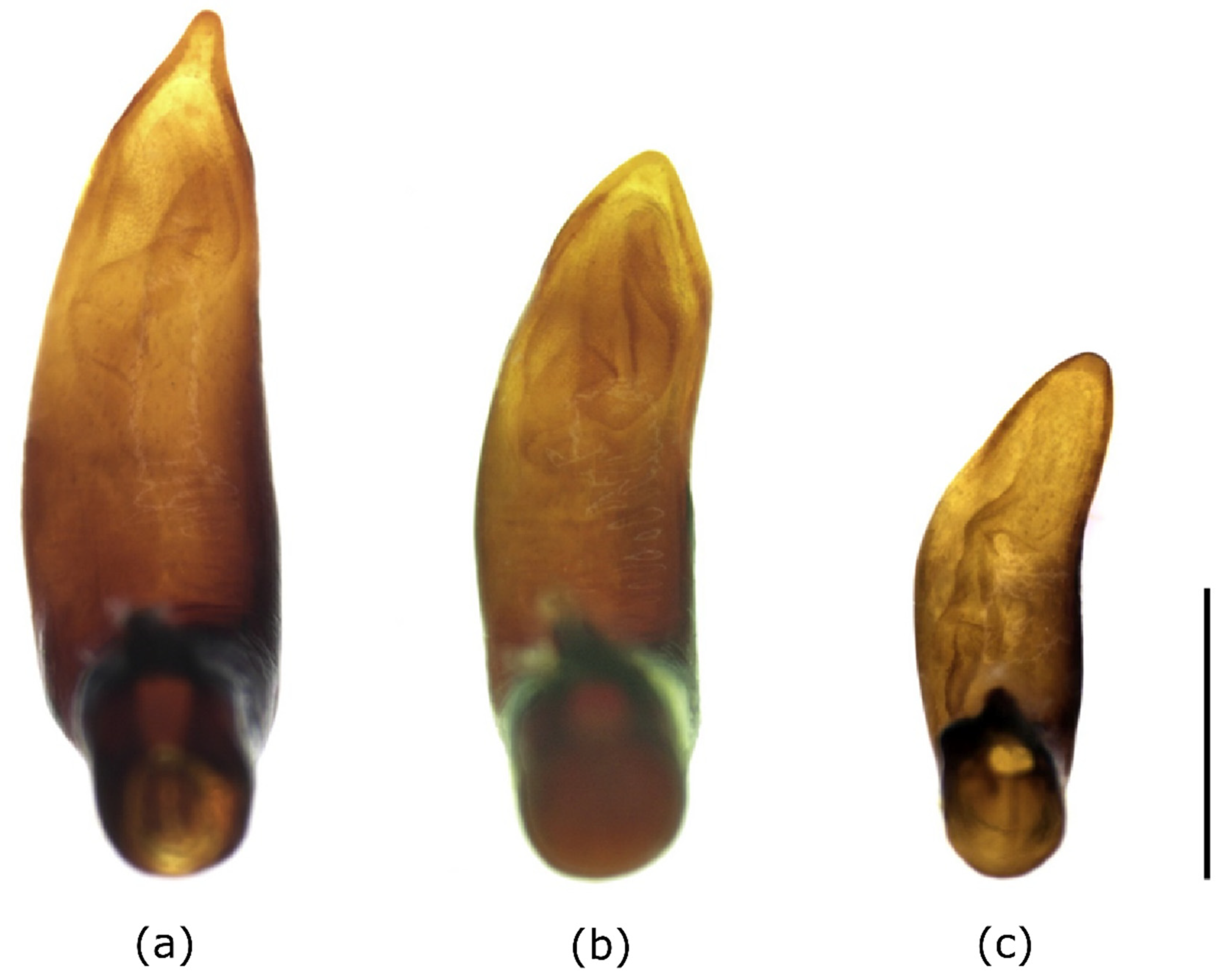
3.1. Morphological Characterization of Austrian P. adstrictus in Comparison to Asian Populations
3.2. Morphological Differentiation and Determination of the European Bothriopterus
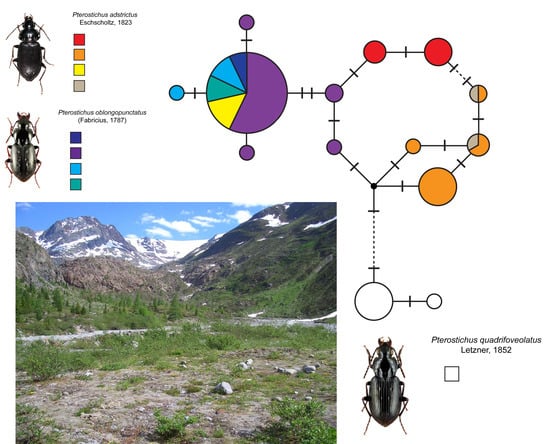
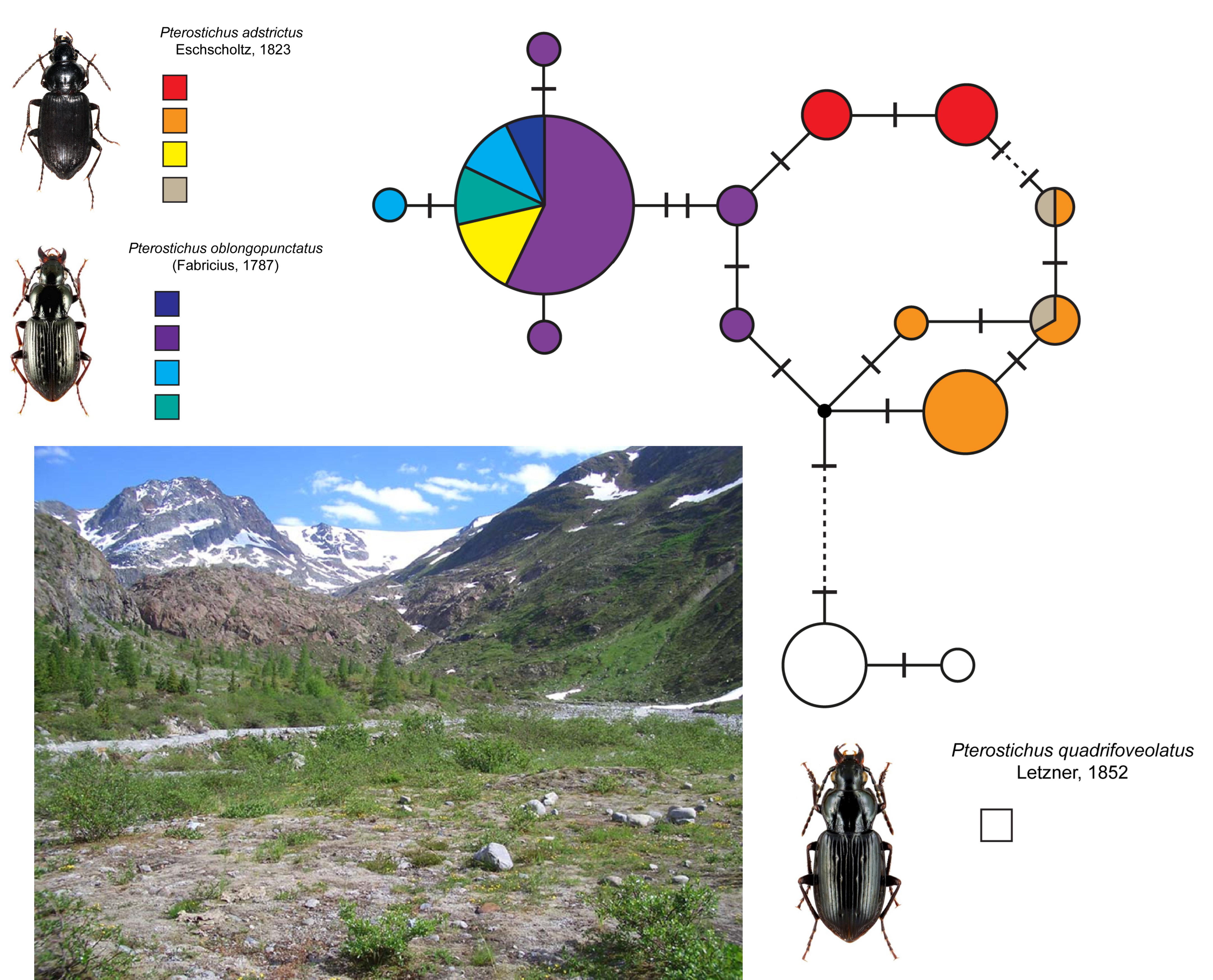
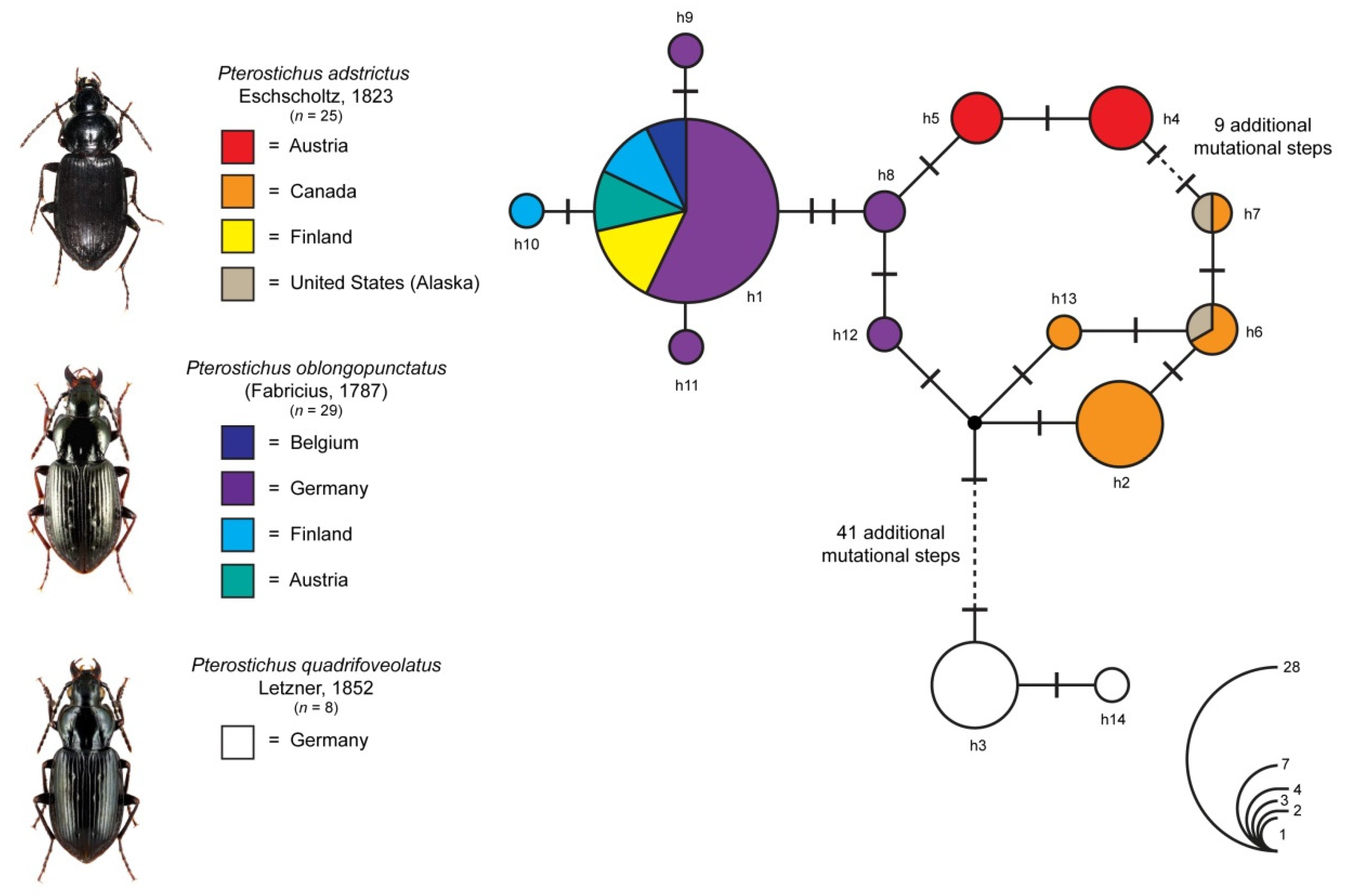
3.3. Analysis of the DNA Barcode Data
3.4. Occurrence and Habitat Use of P. adstrictus in the Ötztal Alps
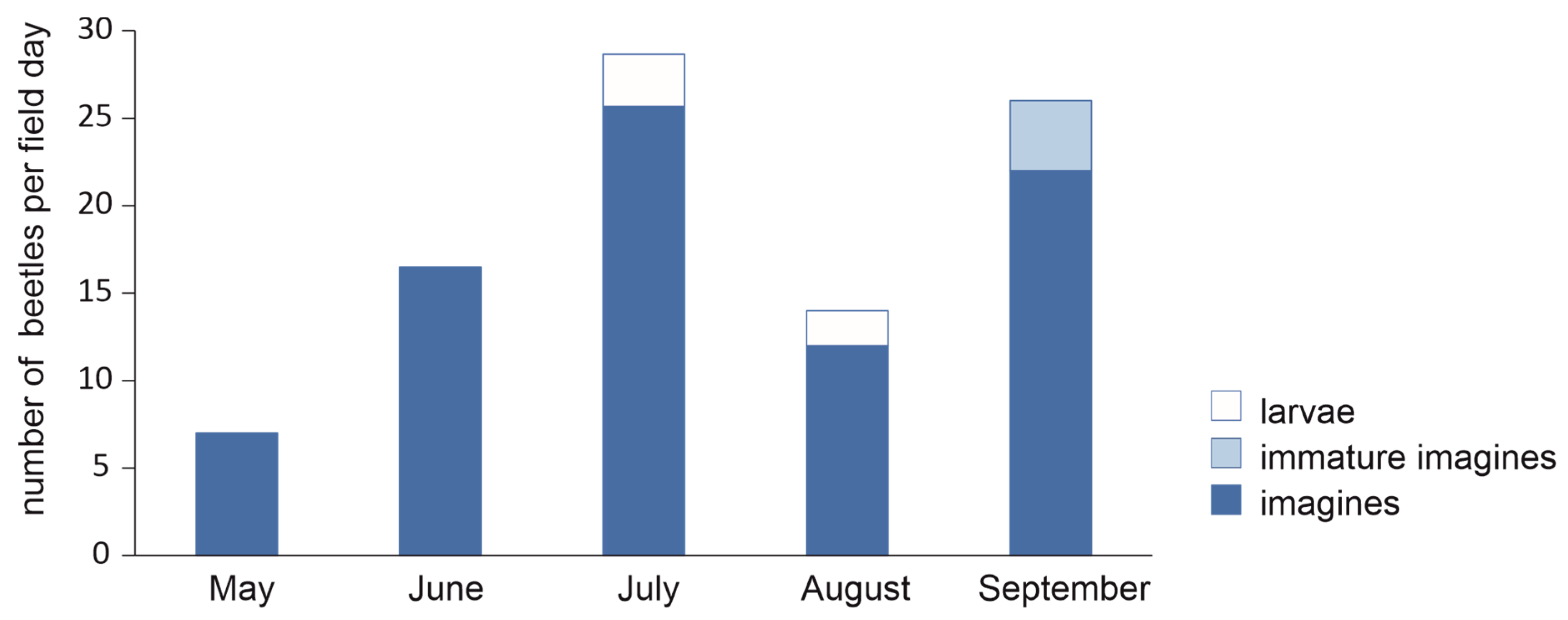
3.5. Seasonality
3.6. Accompanying Ground Beetle Fauna
4. Discussion
4.1. Morphological Characterization
4.2. DNA Barcode Analysis and Origin of Alpine Populations
4.3. Faunistic Interpretation and Historic Data
4.4. Habitat Preference and Physiological Adaptations
4.5. Accompanying Ground Beetle Fauna
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holdhaus, K. Die Spuren der Eiszeit in der Tierwelt Europas. Abh. Zool.-Bot. Ges. Wien. 1954, 18, 1–493. [Google Scholar]
- Hewitt, G. The genetic legacy of the Quaternary ice ages. Nature 2000, 405, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Shafer, A.B.A.; Cullingham, C.I.; Côté, S.D.; Coltman, D.W. Of glaciers and refugia: A decade of study sheds new light on the phylogeography of northwestern North America. Mol. Ecol. 2010, 19, 4589–4621. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, T. Molekulare Biogeographie. Gene in Raum und Zeit, 1st ed.; Haupt Verlag: Bern, Switzerland, 2020. [Google Scholar]
- De Lattin, G. Grundriss der Zoogeographie; Gustav Fischer: Jena, Germany, 1967. [Google Scholar]
- Mani, M.S. The Boreo-Alpine Insects. In Ecology and Biogeography of High-Altitude Insects; Mani, M.S., Ed.; Springer: Dordrecht, The Netherlands, 1968; pp. 313–335. [Google Scholar]
- Muster, C.; Maddison, W.P.; Uhlmann, S.; Berendonk, T.U.; Vogler, A.P. Arctic-Alpine distributions—Metapopulations on a continental scale? Am. Nat. 2009, 173, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Stevanović, V.; Vukojičić, S.; Šinžar-Sekulić, J.; Lazarević, M.; Tomović, G.; Tan, K. Distribution and diversity of Arctic-Alpine species in the Balkans. Plant Syst. Evol. 2009, 283, 219. [Google Scholar] [CrossRef]
- Franz, H. Die Nordost-Alpen im Spiegel ihrer Landtierwelt. Eine Gebietsmonographie. Band III, Coleoptera 1. Teil; Wagner: Innsbruck, Austria; München, Germany, 1970. [Google Scholar]
- Paill, W.; Kahlen, M. Coleoptera (Käfer). In Endemiten—Kostbarkeiten in Österreichs Pflanzen-und Tierwelt; Rabitsch, W., Essl, F., Eds.; Naturwissenschaftlicher Verein und Umweltbundesamt GmbH: Klagenfurt, Austria; Wien, Austria, 2009; pp. 627–783. [Google Scholar]
- Holdhaus, K.; Lindroth, C.H. Die europäischen Koleopteren mit boreoalpiner Verbreitung. Ann. Naturhist. Mus. 1939, 50, 123–293. [Google Scholar]
- Luka, H.; Marggi, W.; Huber, C.; Gonseth, Y.; Nagel, P. Carabidae Ecology—Atlas. Fauna Helv. 2009, 24, 1–677. [Google Scholar]
- Trautner, J.; Fritze, M.-A.; Hannig, K.; Kaiser, M. Verbreitungsatlas der Laufkäfer Deutschlands; Books on Demand: Norderstedt, Germany, 2014. [Google Scholar]
- Paill, W. Patrobini. Adephaga 1: Carabidae (Laufkäfer). In Die Käfer Mitteleuropas, 2nd ed.; Corrected Reprint; Freude, H., Harde, K.W., Lohse, G.A., Klausnitzer, B., Eds.; Spektrum Verlag: Heidelberg/Berlin, Germany, 2006; Volume 2, pp. 211–215. [Google Scholar]
- Bousquet, Y. Tribe Pterostichini. Archostemata-Myxophaga-Adephaga. In Catalogue of Palaearctic Coleoptera; Revised and Updated Edition; Löbl, I., Löbl, D., Eds.; Brill: Leiden, The Netherlands; Boston, MA, USA, 2017; Volume 1, pp. 675–755. [Google Scholar]
- Sundukov, Y.N.; Makarov, K.V. Ground beetles (Coleoptera: Carabidae) of the Russian Far East: Additions and corrections to the Catalogue of Palaearctic Coleoptera, Volume 1 (2017). Zool. Bespozvon. 2019, 16, 283–304. [Google Scholar] [CrossRef]
- Bousquet, Y. Catalogue of Geadephaga (Coleoptera, Adephaga) of America, north of Mexico. ZooKeys 2012, 245, 1–1722. [Google Scholar] [CrossRef]
- Lindroth, C.H. The ground-beetles (Carabidae, excl. Cicindelinae) of Canada and Alaska. Part 4. Opusc. Entom. 1966, 29, 409–648. [Google Scholar]
- Martikainen, P.; Kouki, J.; Heikkala, O. The effects of green tree retention and subsequent prescribed burning on ground beetles (Coleoptera: Carabidae) in boreal pine-dominated forests. Ecography 2006, 29, 659–670. [Google Scholar] [CrossRef]
- Phillips, I.D.; Cobb, T.P.; Spence, J.R.; Brigham, R.M. Salvage logging, edge effects, and Carabid beetles: Connections to conservation and sustainable forest management. Environ. Entomol. 2006, 35, 950–957. [Google Scholar] [CrossRef]
- Pantoja, A.; Sikes, D.S.; Hagerty, A.M.; Emmert, S.Y.; Rondon, S.I. Ground beetle (Coleoptera: Carabidae) assemblages in the Conservation Reserve Program crop rotation systems in interior Alaska. J. Entomol. Soc. B. C. 2013, 110, 6–18. [Google Scholar]
- Lindroth, C.H. Die fennoskandischen Carabidae, eine tiergeographische Studie, I, spezieller Teil. Göteborgs Kungliga Vetenskapsoch Vitterhets-Samhälles Handlingar B 1945, 4, 1–709. [Google Scholar]
- Luff, M.L. Provisional Atlas of the Ground Beetles (Coleoptera, Carabidae) of Britain; Biological Records Centre: Huntingdon, UK, 1998. [Google Scholar]
- Anderson, R.; McFerran, D.; Cameron, A. The Ground Beetles of Northern Ireland (Coleoptera-Carabidae); Ulster Museum: Belfast, Ireland, 2000. [Google Scholar]
- Raupach, M.J.; Hannig, K.; Moriniere, J.; Hendrich, L. A DNA barcode library for ground beetles of Germany: The genus Pterostichus Bonelli, 1810 and allied taxa (Insecta: Coleoptera: Carabidae). Zookeys 2020, 980, 93–117. [Google Scholar] [CrossRef] [PubMed]
- Hilger, L. Quantification and regionalization of geomorphic processes using spatial models and high-resolution topographic data: A sediment budget of the Upper Kauner Valley, Ötztal Alps. Ph.D. Thesis, Catholic University of Eichstätt-Ingolstadt, Eichstätt, Germany, 2017. [Google Scholar]
- Müller-Motzfeld, G. Adephaga 1: Carabidae (Laufkäfer). In Die Käfer Mitteleuropas; Freude, H., Harde, K.W., Lohse, G.A., Klausnitzer, B., Eds.; Spektrum: Heidelberg/Berlin, Germany, 2006; Volume 2, pp. 1–512. [Google Scholar]
- Lindroth, C.H. Coleoptera, Carabidae—Handbooks for the Identification of British insects IV(2); Royal Entomological Society: Saint Albans, UK, 1974. [Google Scholar]
- Bousquet, Y. Illustrated Identification Guide to Adults and Larvae of Northeastern North. American Ground Beetles (Coleoptera: Carabidae); Pensoft: Sofia, Bulgaria, 2010. [Google Scholar]
- Löbl, I.; Löbl, D. Catalogue of Palaearctic Coleoptera; Revised and Updated Edition; Volume 1, Archostemata-Myxophaga-Adephaga; Brill: Leiden, The Netherlands, 2017; pp. 1–1442. [Google Scholar]
- Persohn, M. Faunistische Notizen. 599. Pterostichus adstrictus Eschscholtz, 1823-eine Ergänzung zur Kenntnis der mitteleuropäischen Laufkäfer-Fauna (Col., Carabidae). Entomol. Nachr. Ber. 1996, 40, 252. [Google Scholar]
- Marggi, W. Pterostichini. In Die Käfer Mitteleuropas, 2nd ed.; Corrected, Reprint; Volume 2 Adephaga 1: Carabidae, (Laufkäfer); Freude, H., Harde, K.W., Lohse, G.A., Klausnitzer, B., Eds.; Spektrum Verlag: Heidelberg/Berlin, Germany, 2006; pp. 216–251. [Google Scholar]
- Luff, M. The Carabidae (Ground Beetles) of Britain and Ireland. In Handbooks for the Identification of British Insects, 2nd ed.; Royal Entomological Society: Saint Albans, UK, 2007; Volume 4, Part 2. [Google Scholar]
- Lagisz, M. Changes in morphology of the ground beetle Pterostichus oblongopunctatus F. (Coleoptera; Carabidae) from vicinities of a zinc and lead smelter. Environ. Toxicol. Chem. 2008, 27, 1744–1747. [Google Scholar] [CrossRef]
- Guéorguiev, B.V. Pterostichus Bonelli, 1810. In Ground Beetles of (Carabidae) of Greece; Arndt, E., Schnitter, P.-H., Sfenthourakis, S., Wrase, D., Eds.; Pensoft: Sofia, Bulgaria, 2011; pp. 253–259. [Google Scholar]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; de Waard, J.R. Biological identifications through DNA barcodes. Proc. Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef]
- Hernández-Triana, L.M.; Prosser, S.W.; Rodríguez-Perez, M.A.; Chaverri, L.G.; Hebert, P.D.N.; Ryan Gregory, T. Recovery of DNA barcodes from blackfly museum specimens (Diptera: Simuliidae) using primer sets that target a variety of sequence lengths. Mol. Ecol. Res. 2014, 14, 508–518. [Google Scholar] [CrossRef]
- Duftner, N.; Koblmüller, S.; Sturmbauer, C. Evolutionary relationships of the Limnochromini, a tribe of benthic deepwater cichlid fish endemic to Lake Tanganyika, East Africa. J. Mol. Evol. 2005, 60, 277–289. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. A DNA-based registry for all animal species: The Barcode Index Number (BIN) system. PLoS ONE 2013, 8, e66213. [Google Scholar] [CrossRef] [PubMed]
- Hendrich, L.; Morinière, J.; Haszprunar, G.; Hebert, P.D.N.; Hausmann, A.; Köhler, F.; Balke, M. A comprehensive DNA barcode database for Central European beetles with a focus on Germany: Adding more than 3,500 identified species to BOLD. Mol. Ecol. Res. 2015, 15, 795–818. [Google Scholar] [CrossRef] [PubMed]
- Pentinsaari, M.; Hebert, P.D.N.; Mutanen, M. Barcoding beetles: A regional survey of 1872 species reveals high identification success and unusually deep interspecific divergences. PLoS ONE 2014, 9, e108651. [Google Scholar] [CrossRef] [PubMed]
- Raupach, M.J.; Astrin, J.J.; Hannig, K.; Peters, M.K.; Stoeckle, M.Y.; Wägele, J.W. Molecular species identifications of Central European ground beetles (Coleoptera: Carabidae) using nuclear rDNA expansion segments and DNA barcodes. Front. Zool. 2010, 7, 26. [Google Scholar] [CrossRef] [PubMed]
- Sikes, D.S.; Bowser, M.; Morton, J.M.; Bickford, C.; Meierotto, S.; Hildebrandt, K. Building a DNA barcode library of Alaska‘s non-marine arthropods. Genome 2017, 60, 248–259. [Google Scholar] [CrossRef]
- Woodcock, T.S.; Boyle, E.; Roughley, R.E.; Kevan, P.G.; Labbee, R.N.; Smith, A.B.T.; Goulet, H.; Steinke, D.; Adamowicz, S.J. The diversity and biogeography of the Coleoptera of Churchill: Insights from DNA barcoding. BMC Ecol. 2013, 13, 40. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1660. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Goulet, H. Biology and relationships of Pterostichus adstrictus and Pterostichus pensylvanicus. Quaest. Entomol. 1974, 10, 3–33. [Google Scholar]
- Lorenz, W. CarabCat: Global Database of Ground Beetles (version October 2017). In Species 2000 and ITIS Catalogue of Life; Roskov, Y., Ower, G., Orrell, T., Nicolson, D., Bailly, N., Kirk, P.M., Bourgoin, T., DeWalt, R.E., Decock, W., Nieukerken, E., et al., Eds.; Available online: www.catalogueoflife.org/col (accessed on 16 April 2020).
- Kryzhanovskij, O.L.; Belousov, I.A.; Kabak, I.I.; Kataev, B.M.; Makarov, K.V.; Shilenkov, V.G. A Checklist of the Ground-Beetles of Russia and Adjacent Lands (Insecta, Coleoptera, Carabidae); Pensoft: Sofia, Bulgaria; Moscow, Russia, 1995. [Google Scholar]
- Lindroth, C.H. The carabid beetles of Newfoundland. Opusc. Entom. 1955, 12, 160. [Google Scholar]
- Den Boer, P.J.; Szyszko, J.; Vermeulen, R. Spreading the risk of extinction by genetic diversity in populations of the carabid beetle Pterostichus oblongopunctatus F. (Coleoptera, Carabidae). Neth. J. Zool. 1993, 43, 242–259. [Google Scholar] [CrossRef]
- Schwerk, A.; Jaskuła, R. Rare patterns of dorsal puncture in Pterostichus oblongopunctatus (Coleoptera: Carabidae). PeerJ 2018, 6, e4657. [Google Scholar] [CrossRef] [PubMed]
- Van Husen, D. LGM and late-glacial fluctuations in the Eastern Alps. Quat. Int. 1997, 38, 109–118. [Google Scholar] [CrossRef]
- Seguinot, J.; Ivy-Ochs, S.; Jouvet, G.; Huss, M.; Funk, M.; Preusser, F. Modelling last glacial cycle ice dynamics in the Alps. Cryosphere 2018, 12, 3265–3285. [Google Scholar] [CrossRef]
- Tribsch, A. Areas of endemism of vascular plants in the Eastern Alps in relation to Pleistocene glaciation. J. Biogeogr. 2004, 31, 747–760. [Google Scholar] [CrossRef]
- Schönswetter, P.; Stehlik, I.; Holderegger, R.; Tribsch, A. Molecular evidence for glacial refugia of mountain plants in the European Alps. Mol. Ecol. 2005, 14, 3547–3555. [Google Scholar] [CrossRef]
- Wachter, G.A.; Papadopoulou, A.; Muster, C.; Arthofer, W.; Knowles, L.L.; Steiner, F.M.; Schlick-Steiner, B.C. Glacial refugia, recolonization patterns and diversification forces in Alpine-endemic Megabunus harvestmen. Mol. Ecol. 2016, 25, 2904–2919. [Google Scholar] [CrossRef]
- Schönswetter, P.; Schneeweiss, G.M. Is the incidence of survival in interior Pleistocene refugia (nunataks) underestimated? Phylogeography of the high mountain plant Androsace alpina (Primulaceae) in the European Alps revisited. Ecol. Evol. 2019, 9, 4078–4086. [Google Scholar] [CrossRef] [PubMed]
- Brower, A.V.Z. Rapid morphological radiation and convergence among races of the butterfly Heliconius erato inferred from patterns of mitochondrial DNA evolution. Proc. Natl. Acad. Sci. USA 1994, 91, 6491–6495. [Google Scholar] [CrossRef] [PubMed]
- Pentinsaari, M.; Salmela, H.; Mutanen, M.; Roslin, T. Molecular evolution of a widely-adopted taxonomic marker (COI) across the animal tree of life. Sci. Rep. 2016, 6, 35275. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.Y.W.; Phillips, M.J.; Cooper, A.; Drummond, A.J. Time dependency of molecular rate estimates and systematic overestimation of recent divergence times. Mol. Biol. Evol. 2005, 22, 1561–1568. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.Y.W.; Lanfear, R.; Bromham, L.; Phillips, M.J.; Soubrier, J.; Rodrigo, A.G.; Cooper, A. Time-dependent rates of molecular evolution. Mol. Ecol. 2011, 20, 3087–3101. [Google Scholar] [CrossRef] [PubMed]
- Gratton, P.; Konopiński, M.K.; Sbordoni, V. Pleistocene evolutionary history of the Clouded Apollo (Parnassius mnemosyne): Genetic signatures of climate cycles and a ‘time-dependent’ mitochondrial substitution rate. Mol. Ecol. 2008, 17, 4248–4262. [Google Scholar] [CrossRef]
- Heliölä, J.; Koivula, M.; Niemelä, J. Distribution of carabid beetles (Coleoptera, Carabidae) across boreal forest-clearcut ecotone. Conserv. Biol. 2001, 15, 370–377. [Google Scholar] [CrossRef]
- Nevado, B.; Koblmüller, S.; Sturmbauer, C.; Snoeks, J.; Usano-Alemany, J.; Verheyen, E. Complete mitochondrial DNA replacement in a Lake Tanganyika cichlid fish. Mol. Ecol. 2009, 18, 4240–4255. [Google Scholar] [CrossRef]
- Koblmüller, S.; Nevado, B.; Makasa, L.; Van Steenberge, M.; Vanhove, M.P.M.; Verheyen, E.; Sturmbauer, C.; Sefc, K.M. Phylogeny and phylogeography of Altolamprologus: Ancient introgression and recent divergence in a rock-dwelling Lake Tanganyika cichlid genus. Hydrobiologia 2015, 791, 35–50. [Google Scholar] [CrossRef]
- Good, J.M.; Vanderpool, D.; Keeble, S.; Bi, K. Negligible nuclear introgression despite complete mitochondrial capture between two species of chipmunks. Evolution 2015, 69, 1961–1972. [Google Scholar] [CrossRef]
- Hertach, T.; Puissant, S.; Gogala, M.; Trilar, T.; Hagmann, R.; Baur, H.; Kunz, G.; Wade, E.J.; Loader, S.P.; Simon, C.; et al. Complex within a complex: Integrative taxonomy reveals hidden diversity in Cicadetta brevipennis (Hemiptera: Cicadidae) and unexpected relationships with a song divergent relative. PLoS ONE 2016, 11, e0165562. [Google Scholar] [CrossRef] [PubMed]
- Sota, T. Radiation and reticulation: Extensive introgressive hybridization in the carabid beetles Ohomopterus inferred from mitochondrial gene genealogy. Popul. Ecol. 2002, 44, 145–156. [Google Scholar] [CrossRef]
- Takami, Y.; Suzuki, H. Morphological, genetic and behavioural analyses of a hybrid zone between the ground beetles Carabus lewisianus and C. albrechti (Coleoptera, Carabidae): Asymmetrical introgression caused by movement of the zone? Biol. J. Linnean Soc. 2005, 86, 79–94. [Google Scholar] [CrossRef]
- Zanella, L. Biodiversity of the endangered coastal beetle Scarites laevigatus: Is the northern Adriatic population a geographical subspecies or a case of introgressive hybridisation? (Coleoptera: Carabidae). Fragm. Entomol. 2018, 50, 149–160. [Google Scholar] [CrossRef]
- Kosuda, S.; Sasakawa, K.; Ikeda, H. Directional mitochondrial introgression and character displacement due to reproductive interference in two closely related Pterostichus ground beetle species. J. Evol. Biol. 2016, 26, 1121–1130. [Google Scholar] [CrossRef]
- Gredler, V.M. Die Käfer von Tirol Nach Ihrer Horizontalen Und Vertikalen Verbreitung; Eberle: Bozen, Italy, 1863. [Google Scholar]
- Ammann, J.; Knabl, H. Die Käferfauna des Oetztals (Tirol). Coleopterol. Rundsch. 1912, 3, 36–61. [Google Scholar]
- Steinböck, O. Die Nunatak-Fauna der Venter Berge. In Das Venter Tal; Bruckmann, F., Ed.; Sektion Mark Brandenburg des deutschen Alpenvereins: Munich, Germany, 1939; pp. 63–73. [Google Scholar]
- Wörndle, A. Die Käfer von Nordtirol. Schlern-Schriften 1950, 64, 1–388. [Google Scholar]
- Janetschek, H. Tierische Successionen auf hochalpinem Neuland. Nach Untersuchungen am Hintereis-, Niederjoch- und Gepatschferner in den Ötztal Alpen. Schlern-Schriften 1949, 76, 1–215. [Google Scholar]
- Janetschek, H. Die beiden großen Alpenexkursionen. d) Die Alpen von Zell am See bis Bregenz. In Exkursionsführer zum XIth International Congress of Entomology; Stehlicek and Pühringer: Vienna, Austria, 1960; pp. 115–191. [Google Scholar]
- Janetschek, H.; Meyer, E.; Schatz, H.; Schatz, I. Ökologische Untersuchungen an Wirbellosen im Raum Gurgl unter Berücksichtigung anthropogener Einflüsse. In MaB-Projekt Obergurgl; Patzelt, G., Ed.; Wagner: Innsbruck, Austria, 1987; Volume 10, pp. 281–315. [Google Scholar]
- De Zordo, I. Ökologische Untersuchungen an Wirbellosen des zentralalpinen Hochgebirges (Obergurgl, Tirol). III. Lebenszyklen und Zönotik von Coleopteren. Alpin.-Biol. Stud. 1979, 11, 131. [Google Scholar]
- Thaler, K. Fragmenta Faunistica Tirolensia—VI. Ber. Naturwiss.-Med. Ver. Innsb. 1984, 71, 97–118. [Google Scholar]
- Schatz, I. Life strategy of an alpine carabid: Pterostichus jurinei (Coleoptera, Carabidae). In Carabid Beetles: Ecology and Evolution; Desender, K., Dufrene, M., Loreau, M., Luff, M.L., Maelfait, J.-P., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1994; pp. 213–217. [Google Scholar]
- Troger, H.; Janetschek, H.; Meyer, E.; Schatz, W. Schlüpfabundanz von Insekten (Diptera/Coleoptera/Hymenoptera) im zentralalpinen Hochgebrige (Tirol: Ötztal). Entomol. Gen. 1994, 18, 241–260. [Google Scholar] [CrossRef]
- Kopf, T.; Link, A.; Schatz, I. Käfer (Coleoptera)—307 Arten. Ber. Naturwiss.-Med. Ver. Innsb. 2005, 92, 275–281. [Google Scholar]
- Link, A.; Schuh, R.; Kopf, T. Zikaden (Auchenorrhyncha)—2 Arten, Wanzen (Heteroptera)—20 Arten, Käfer Coleoptera—151 Arten, Schnaken (Diptera: Tipulidae)—4 Arten. Veröffentlichungen des Tiroler Landesmuseums Ferdinandeum 2007, 87, 73–170. [Google Scholar]
- Kaufmann, R. Invertebrate succession on an alpine glacier foreland. Ecology 2001, 82, 2261–2278. [Google Scholar] [CrossRef]
- Kaufmann, R.; Juen, A. Habitat use and niche segregation of the genus Nebria (Coleoptera: Carabidae) in the Austrian Alps. Mitt. Schweiz. Entomol. Ges. 2002, 74, 237–254. [Google Scholar]
- Raso, L.; Sint, D.; Mayer, R.; Plangg, S.; Recheis, T.; Brunner, S.; Kaufmann, R.; Traugott, M. Intraguild predation in pioneer predator communities of alpine glacier forelands. Mol. Ecol. 2014, 23, 3744–3754. [Google Scholar] [CrossRef] [PubMed]
- Brandstetter, C.M.; Kapp, A.; Schabel, F. Die Laufkäfer von Vorarlberg und Liechtenstein, 1. Band (Carabidae); Erster Vorarlberger Coleopterologischer Verein: Bürs, Austria, 1993. [Google Scholar]
- Kofler, A. Zur Laufkäferfauna im Bezirk Lienz: Osttirol (Österreich) (Coleoptera: Carabidae). Ber. Naturwiss.-Med. Ver. Innsb. 2005, 92, 189–220. [Google Scholar]
- Marggi, W. Faunistik der Sandlaufkäfer und Laufkäfer der Schweiz (Cicindelidae and Carabidae), Coleoptera, Teil 1. Doc. Faun. Helv. 1992, 13, 477. [Google Scholar]
- Märkel, F.; Kiesenwetter, H. Bericht über eine entomologische Excursion in die Kärnthner Alpen im Jahre 1847. Stett. Ent. Zeit. 1848, 9, 210–221, 277–285, 314–320. [Google Scholar]
- Pacher, D. Ueber die Käfer in den Umgebungen von Sagritz und Heiligenblut. Jahrbuch Des Naturhistorischen Landesmuseums Von Kärnten 1853, 2, 30–52. [Google Scholar]
- Liegel, E. Nachträge zum „Verzeichnis der Käfer Deutschlands“ aus der kärntnerischen Fauna. Dtsch. Entomol. Z. 1890, 1, 205–208. [Google Scholar]
- Franz, H. Die Landtierwelt der mittleren Hohen Tauern. Denkschr. oest. Akad. Wiss. Mathem. Naturw. Kl. 1943, 107, 552. [Google Scholar]
- Franz, H. Erster Nachtrag zur Landtierwelt der mittleren Hohen Tauern. Akad. Wiss. Wien. Math.-Nat. Kl. 1949, 158, 1–77. [Google Scholar]
- Mandl, K.; Schönmann, R. Catalogus Faunae Austriae. Teil XVa: Coleoptera, Carabidae II; Österreichische Akademie der Wissenschaften: Vienna, Austria, 1978; pp. 1–58. [Google Scholar]
- Holdhaus, K.; Prossen, T. Verzeichnis der bisher in Kärnten beobachteten Käfer. Carinthia II 1900, 90, 102–121. [Google Scholar]
- Paill, W.; Schnitter, P.-H. Rote Liste der Laufkäfer Kärntens (Carabidae). Naturschutz in Kärnten 1999, 15, 369–412. [Google Scholar]
- Lindroth, C.H. The Carabidae (Coleoptera) of Fennoscandia and Denmark. Fauna Entomol. Scand. 1986, 15, 1–497. [Google Scholar]
- Lindroth, C.H.; Andersson, H.; Bödvarsson, H.; Richter, S.H. Surtsey, Iceland—The development of a new fauna, 1963–1970, Terrestrial Arthropods. Entomol. Scand. 1973, 5, 1–280. [Google Scholar]
- Voronin, A.G. On sector related change of biotopes by some ground beetle species (Coleoptera, Trachypachidae, Carabidae) in the forest zone of the Palearctic. Russ. J. Ecol. 2011, 42, 344–346. [Google Scholar] [CrossRef]
- Larochelle, A.; Larivière, M.-C. A Natural History of the Ground-Beetles (Coleoptera: Carabidae) of America North of Mexico; Faunistica Pensoft Publishers: Sofia, Bulgaria; Moscow, Russia, 2003; Volume 27, p. 583. [Google Scholar]
- Saint-Germain, M.; Larrivée, M.; Drapeau, P.; Fahrig, L.; Buddle, C.M. Short-term response of ground beetles (Coleoptera: Carabidae) to fire and logging in a spruce-dominated boreal landscape. For. Ecol. Manag. 2005, 212, 118–126. [Google Scholar] [CrossRef]
- Gongalsky, K.B.; Midtgaard, F.; Overgaard, H.J. Effects of prescribed forest burning on carabid beetles (Coleoptera: Carabidae): A case study in southeastern Norway. Entomol. Fennica 2006, 17, 325–333. [Google Scholar] [CrossRef]
- Gandhi, K.J.K.; Gilmore, D.W.; Katovich, S.A.; Mattson, W.J.; Zasada, J.C.; Seybold, S.J. Catastrophic windstorm and fuel-reduction treatments alter ground beetle (Coleoptera: Carabidae) assemblages in a North American sub-boreal forest. For. Ecol. Manag. 2008, 256, 1104–1123. [Google Scholar] [CrossRef]
- Koivulaa, M.J.; Venn, S.; Hakolaa, P.; Niemelä, J. Responses of boreal ground beetles (Coleoptera, Carabidae) to different logging regimes ten years post harvest. For. Ecol. Manag. 2019, 436, 27–38. [Google Scholar] [CrossRef]
- Butterfield, J. Carabid community succession during the forestry cycle in conifer plantations. Ecography 1997, 20, 614–625. [Google Scholar] [CrossRef]
- Jacobs, J.M.; Spence, J.R.; Langor, D.W. Influence of boreal forest succession and dead wood qualities on saproxylic beetles. Agric. For. Entomol. 2007, 9, 3–16. [Google Scholar] [CrossRef]
- Den Boer, P.J. Dispersal power and survival. Carabids in a cultivated countryside. Misc. Papers Landbouwhogeschool Wageningen 1977, 14, 1–190. [Google Scholar]
- Luff, M. The Carabidae (Coleoptera) larvae of Fennoscandia and Denmark. Fauna Entomol. Scand. 1993, 27, 186. [Google Scholar]
- Andersen, J. The life cycles of carabid beetles (Coleoptera, Carabidae) in dry, open habitats north of 69°N, Northern Norway. Nor. J. Entomol. 2013, 60, 140–158. [Google Scholar]
- Paarmann, W. Temperature and photoperiodic influence on developmental stages and adults of the subarctic carabid beetle Pterostichus adstrictus (Coleoptera). In Carabid Beetles: Ecology and Evolution; Desender, K., Dufrene, M., Loreau, M., Luff, M.L., Maelfait, J.-P., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2014; pp. 201–205. [Google Scholar]
- Kopf, T. Die Laufkäferfauna (Coleoptera: Carabidae) des Schlerngebietes (Südtirol, Italien) mit Angaben zu den Artengemeinschaften ausgewählter Lebensräume. Gredleriana 2008, 8, 341–366. [Google Scholar]
- Magistretti, M. Coleoptera Cicindelidae, Carabidae. Fauna d’Italia 8; Calderini: Bologna, Italy, 1965; p. 512. [Google Scholar]
- Balkenohl, M. Coleoptera Westfalica: Familia Carabidae, Subfamiliae Scaritinae et Broscinae. Abh. Westf. Mus. Naturk. 1988, 50, 3–28. [Google Scholar]
- Gobbi, M.; Fontaneto, D.; Bragalanti, N.; Pedrotti, l.; Lencioni, V. Carabid beetle (Coleoptera: Carabidae) richness and functional traits in relation to differently managed grasslands in the Alps. Ann. Soc. Entomol. Fr. 2015, 51, 52–59. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Date | Origin | Collector | Altitude |
|---|---|---|---|
| 11.08.2009 | AUT, North Tyrol, Fissladbachtal | Christian Mairhuber | 1990–2230 m |
| 16.07.2009 | AUT, North Tyrol, Griestal near Mandarfen | Wolfgang Paill | 1840–2000 m |
| 17.06.2009, 18.06.2009, 13.07.2009, 14.07.2009, 15.07.2009, 11.08.2009, 25.08.2009, 01.09.2009, 17.06.–14.07.2009, 10.05.2010, 22.09.2010, 16.07.2016, 13.08.2016 | AUT, North Tyrol, Kaunertal | Thomas Frieß, Barbara-Amina Gereben-Krenn, Christian Mairhuber, Wolfgang Paill | 1870–2270 m |
| 11.08.2009 | AUT, North Tyrol, Kaiserbergtal | Wolfgang Paill | 2160–2520 m |
| 11.08.2009, 09.06.2010, 13.07.2010, 21.07.2010, 18.08.2007, 19.08.2010, 21.09.2010, 09.06.-13.07.2010, 13.07.-19.08.2010, 19.08.-21.09.2010 | AUT, North Tyrol, Platzertal | Thomas Frieß, Wolfgang Paill | 2100–2520 m |
| 12.08.2009 | AUT, North Tyrol, Radurschlbachtal | Thomas Frieß, Christian Mairhuber | 2210–2280 m |
| 12.08.2009 | AUT, North Tyrol, Rifflbachtal | Wolfgang Paill | 2230–2410 m |
| 13.08.2009 | AUT, North Tyrol, Rofental | Christian Mairhuber, Wolfgang Paill | 2340–2390 m |
| 18.06.2009, 15.07.2009, 26.08.2009 | AUT, North Tyrol, Taschachtal | Christian Mairhuber, Wolfgang Paill | 1970–2110 m |
| 12.08.2009 | ITA, South Tyrol, Langtauferertal | Wolfgang Paill | 1890–2020 m |
| P. adstrictus | P. oblongopunctatus | P. quadrifoveolatus | |
|---|---|---|---|
| Number of the foveolate punctures on 3rd elytral interval | mostly 5 or 6 (4–8) | mostly 4 (2–9) | mostly 3 (2–4) |
| Base of the pronotum from inner impression to posterior angle | almost straight or moderately angled forwards laterally | almost straight | angled forwards laterally |
| Side border of the pronotum | moderately wide, widened towards posterior angle | narrow, not widened towards posterior angle | moderately wide, not widened towards posterior angle |
| Vertex (head behind the eyes) | without punctures | without punctures | with punctures |
| Color of the elytra | black, with bronzed luster, in females dull | black, with bronzed to greenish luster | black, with bronzed luster |
| Color of the tibiae | blackish, almost as dark as femora | reddish-brownish, distinctly paler than femora | blackish, almost as dark as femora |
| Color of the palpi | Blackish | reddish-brownish | blackish |
| 1st antennal segment | as long as 3rd segment | as long as 3rd segment | clearly shorter than 3rd segment |
| Shape of the aedeagus in lateral view | less evenly arcuate, apex long | more evenly arcuate, apex short | more evenly arcuate, apex long |
| Shape of the aedeagus in ventral view | aedeagus almost straight, apex acuminate | aedeagus weakly bent, apex almost obtuse | aedeagus clearly bent, apex obtuse |
| Size of the aedeagus in lateral view | big (about 3 mm) | big (about 3 mm) | small (about 2 mm) |
| Size of the membranous wings | relative long (ratio wing to elytra >1.3) | relative short (ratio wing to elytra <1.3) | relative long (ratio wing to elytra >1.3) |
| Total body length (data from literature) | 10.4–13 mm | 9.5–13.0 mm (mean: 11.4) | 8.5–11.9 mm (mean: 10.3) |
| P. adstrictus (Austria) | P. adstrictus (North America) | P. adstrictus (Finland)/ P. oblongopunctatus (Austria/Belgium/ Finland/Germany) | P. quadrifoveolatus (Germany) | |
|---|---|---|---|---|
| P. adstrictus (Austria) | 0–0.0015 | |||
| P. adstrictus (North America) | 0.0169–0.0217 | 0–0.003 | ||
| P. adstrictus (Finland)/ P. oblongopunctatus (Austria/Belgium/ Finland/Germany) | 0.0015–0.0077 | 0.0201–0.028 | 0–0.0061 | |
| P. quadrifoveolatus (Germany) | 0.0720–0.0753 | 0.0702–0.0737 | 0.0702–0.0707 | 0–0.0015 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paill, W.; Koblmüller, S.; Friess, T.; Gereben-Krenn, B.-A.; Mairhuber, C.; Raupach, M.J.; Zangl, L. Relicts from Glacial Times: The Ground Beetle Pterostichus adstrictus Eschscholtz, 1823 (Coleoptera: Carabidae) in the Austrian Alps. Insects 2021, 12, 84. https://doi.org/10.3390/insects12010084
Paill W, Koblmüller S, Friess T, Gereben-Krenn B-A, Mairhuber C, Raupach MJ, Zangl L. Relicts from Glacial Times: The Ground Beetle Pterostichus adstrictus Eschscholtz, 1823 (Coleoptera: Carabidae) in the Austrian Alps. Insects. 2021; 12(1):84. https://doi.org/10.3390/insects12010084
Chicago/Turabian StylePaill, Wolfgang, Stephan Koblmüller, Thomas Friess, Barbara-Amina Gereben-Krenn, Christian Mairhuber, Michael J. Raupach, and Lukas Zangl. 2021. "Relicts from Glacial Times: The Ground Beetle Pterostichus adstrictus Eschscholtz, 1823 (Coleoptera: Carabidae) in the Austrian Alps" Insects 12, no. 1: 84. https://doi.org/10.3390/insects12010084
APA StylePaill, W., Koblmüller, S., Friess, T., Gereben-Krenn, B.-A., Mairhuber, C., Raupach, M. J., & Zangl, L. (2021). Relicts from Glacial Times: The Ground Beetle Pterostichus adstrictus Eschscholtz, 1823 (Coleoptera: Carabidae) in the Austrian Alps. Insects, 12(1), 84. https://doi.org/10.3390/insects12010084