
First Evaluation of Field Evolved Resistance to Commonly Used Insecticides in House Fly Populations from Saudi Arabian Dairy Farms


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection and Rearing of House Fly Populations
2.2. Insecticides
2.3. Adult Diet Incorporation Bioassay
2.4. Data Analysis
3. Results
3.1. Resistance to Pyrethroids
3.2. Resistance to Organophosphates
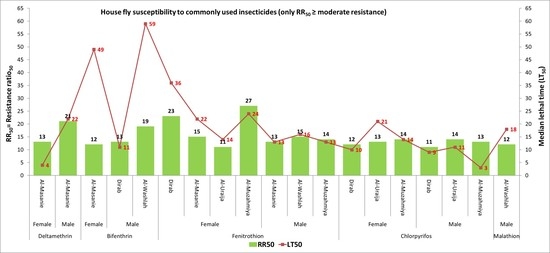
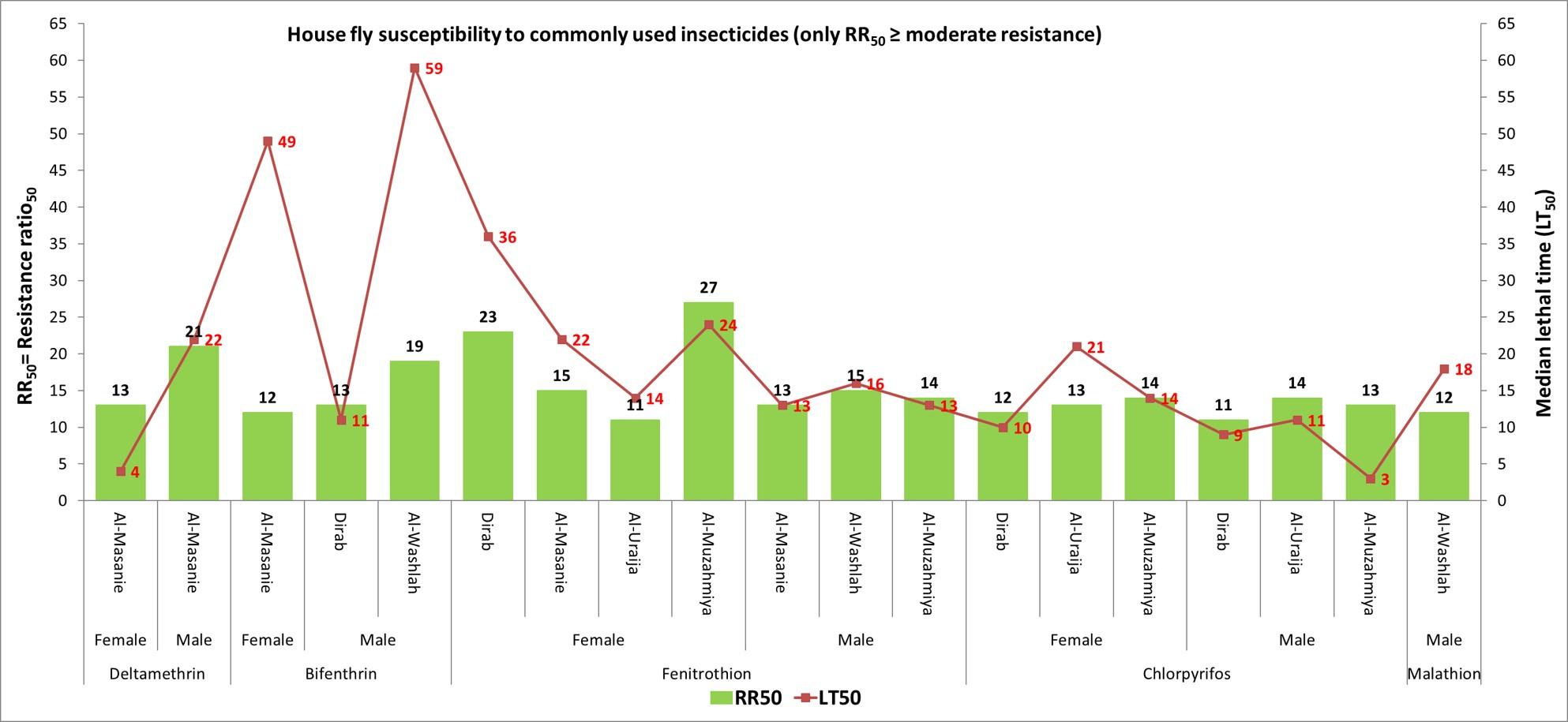
3.3. LT50 of Pyrethroids and Organophosphates
4. Discussion
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abbas, N.; Ijaz, M.; Shad, S.A.; Binyameen, M. Assessment of Resistance Risk to Fipronil and Cross Resistance to Other Insecticides in the Musca domestica L. (Diptera: Muscidae). Vet. Parasitol. 2016, 223, 71–76. [Google Scholar] [CrossRef]
- Ma, Z.; Li, J.; Zhang, Y.; Shan, C.; Gao, X. Inheritance Mode and Mechanisms of Resistance to Imidacloprid in the House Fly Musca domestica (Diptera: Muscidae) from China. PLoS ONE 2017, 12, e0189343. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, P.E.; Nunez, S.C.; Mann, R.S.; Geden, C.J.; Scharf, M.E. Nicotinoid and Pyrethroid Insecticide Resistance in Houseflies (Diptera: Muscidae) Collected from Florida Dairies. Pest Manag. Sci. 2010, 66, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Alzahrani, S.; Ajlan, A.; Hajjar, M.J. Resistance of Field Strains of House Fly Musca domestica L. to Three Selected Synthetic Pyrethroid Insecticides in Riyadh City, Saudi Arabia. Arab J. Plant Prot. 2015, 33, 66–71. [Google Scholar]
- Sarwar, M. Insects Effecting by Annoyance to Peoples Relating to the Public Health Concerns. Am. J. Clin. Neurol. Neurosurg. 2015, 1, 175–181. [Google Scholar]
- Fasanella, A.; Scasciamacchia, S.; Garofolo, G.; Giangaspero, A.; Tarsitano, E.; Adone, R. Evaluation of the House Fly Musca domestica as a Mechanical Vector for an Anthrax. PLoS ONE 2010, 5, e12219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nayduch, D.; Burrus, R.G. Flourishing in Filth: House Fly–Microbe Interactions across Life History. Ann. Entomol. Soc. Am. 2017, 110, 6–18. [Google Scholar] [CrossRef]
- Abbas, N.; Ali Shad, S.A.; Ismail, M. Resistance to Conventional and New Insecticides in House Flies (Diptera: Muscidae) from Poultry Facilities in Punjab, Pakistan. J. Econ. Entomol. 2015, 108, 826–833. [Google Scholar] [CrossRef]
- Khan, H.A.A.; Akram, W.; Fatima, A. Resistance to Pyrethroid Insecticides in House Flies, Musca domestica L., (Diptera: Muscidae) Collected from Urban Areas in Punjab, Pakistan. Parasitol. Res. 2017, 116, 3381–3385. [Google Scholar] [CrossRef] [PubMed]
- Freeman, J.C.; Ross, D.H.; Scott, J.G. Insecticide Resistance Monitoring of House Fly Populations from the United States. Pestic. Biochem. Physiol. 2019, 158, 61–68. [Google Scholar] [CrossRef]
- Wang, J.N.; Hou, J.; Wu, Y.Y.; Guo, S.; Liu, Q.M.; Li, T.Q.; Gong, Z.Y. Resistance of House Fly, Musca domestica L. (Diptera: Muscidae), to Five Insecticides in Zhejiang Province, China: The Situation in 2017. Can. J. Infect. Dis. Med. Microbiol. 2019, 2019, 4851914. [Google Scholar] [CrossRef] [Green Version]
- Soares, M.A.; Passos, L.C.; Campos, M.R.; Collares, L.J.; Desneux, N.; Carvalho, G.A. Side Effects of Insecticides Commonly Used Against Tuta absoluta on the Predator Macrolophus basicornis. J. Pest Sci. 2019, 92, 1447–1456. [Google Scholar] [CrossRef]
- Ippolito, A.; Kattwinkel, M.; Rasmussen, J.J.; Schäfer, R.B.; Fornaroli, R.; Liess, M. Modeling Global Distribution of Agricultural Insecticides in Surface Waters. Environ. Pollut. 2015, 198, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Saeed, R.; Abbas, N.; Mehmood, Z. Emamectin Benzoate Resistance Risk Assessment in Dysdercus Koenigii: Cross-Resistance and Inheritance Patterns. Crop Prot. 2020, 130, 105069. [Google Scholar] [CrossRef]
- Saeed, R.; Abbas, N. Realized Heritability, Inheritance and Cross-Resistance Patterns in Imidacloprid-Resistant Strain of Dysdercus Koenigii (Fabricius) (Hemiptera: Pyrrhocoridae). Pest Manag. Sci. 2020, 76, 2645–2652. [Google Scholar] [CrossRef]
- Abbas, N.; Khan, H.A.A.; Shad, S.A. Resistance of the House Fly Musca domestica (Diptera: Muscidae) to Lambda-Cyhalothrin: Mode of Inheritance, Realized Heritability, and Cross-Resistance to Other Insecticides. Ecotoxicology 2014, 23, 791–801. [Google Scholar] [CrossRef]
- Khan, H.A.A. Characterization of Permethrin Resistance in a Musca domestica strain: Resistance Development, Cross-Resistance Potential and Realized Heritability. Pest Manag. Sci. 2019, 75, 2969–2974. [Google Scholar] [CrossRef] [PubMed]
- Roditakis, E.; Skarmoutsou, C.; Staurakaki, M. Toxicity of Insecticides to Populations of Tomato Borer Tuta absoluta (Meyrick) from Greece. Pest Manag. Sci. 2013, 69, 834–840. [Google Scholar] [CrossRef] [PubMed]
- Abbas, N.; Shad, S.A.; Shah, R.M. Resistance Status of Musca domestica L. Populations to Neonicotinoids and Insect Growth Regulators in Pakistan Poultry Facilities. Pak. J. Zool. 2015, 47, 1663–1671. [Google Scholar]
- Shah, R.M.; Alam, M.; Ahmad, D.; Waqas, M.; Ali, Q.; Binyamin, M.; Shad, S.A. Toxicity of 25 Synthetic Insecticides to the Field Population of Culex quinquefasciatus Say. Parasitol. Res. 2016, 115, 4345–4351. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Huang, J.; Yuan, J. Status and Preliminary Mechanism of Resistance to Insecticides in a Field Strain of House Fly (Musca domestica, L). Rev. Bras. Entomol. 2018, 62, 311–314. [Google Scholar] [CrossRef]
- LeOra, S. Poloplus, a User’s Guide to Probit or Logit Analysis; LeOra Software: Berkeley, CA, USA, 2003. [Google Scholar]
- Abbott, W.S. A Method of Computing the Effectiveness of an Insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Litchfield, J.T.; Wilcoxon, F. A Simplified Method of Evaluating Dose-Effect Experiments. J. Pharmacol. Exp. Ther. 1949, 96, 99–113. [Google Scholar]
- Torres-Vila, L.M.; Rodrıguez-Molina, M.C.; Lacasa-Plasencia, A.; Bielza-Lino, P. Insecticide Resistance of Helicoverpa armigera to Endosulfan, Carbamates and Organophosphates: The Spanish Case. Crop Prot. 2002, 21, 1003–1013. [Google Scholar] [CrossRef]
- Abbas, N.; Hafez, A.M. Resistance to Insect Growth Regulators and Age-Stage, Two-Sex Life Table in Musca domestica from Different Dairy Facilities. PLoS ONE 2021, 16, e0248693. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Mota-Sanchez, D.; Whalon, M.E.; Hollingworth, R.M.; Carrière, Y. Defining Terms for Proactive Management of Resistance to Bt Crops and Pesticides. J. Econ. Entomol. 2014, 107, 496–507. [Google Scholar] [CrossRef] [PubMed]
- Akıner, M.M.; Çağlar, S.S. Monitoring of Five Different Insecticide Resistance Status in Turkish House Fly Musca domestica L. (Diptera: Muscidae) Populations and the Relationship between Resistance and Insecticide Usage Profile. Turkiye Parazitol. Derg. 2012, 36, 87–91. [Google Scholar] [CrossRef]
- Abbas, N.; Shad, S.A. Assessment of Resistance Risk to Lambda-Cyhalothrin and Cross-Resistance to Four Other Insecticides in the House Fly, Musca domestica L. (Diptera: Muscidae). Parasitol. Res. 2015, 114, 2629–2637. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.A.A.; Akram, W.; Shad, S.A. Resistance to Conventional Insecticides in Pakistani Populations of Musca domestica L. (Diptera: Muscidae): A Potential Ectoparasite of Dairy Animals. Ecotoxicology 2013, 22, 522–527. [Google Scholar] [CrossRef]
- Smith, L.B.; Kasai, S.; Scott, J.G. Pyrethroid Resistance in Aedes aegypti and Aedes albopictus: Important Mosquito Vectors of Human Diseases. Pestic. Biochem. Physiol. 2016, 133, 1–12. [Google Scholar] [CrossRef]
- Scott, J.G.; Yoshimizu, M.H.; Kasai, S. Pyrethroid Resistance in Culex pipiens Mosquitoes. Pestic. Biochem. Physiol. 2015, 120, 68–76. [Google Scholar] [CrossRef]
- Protopopoff, N.; Matowo, J.; Malima, R.; Kavishe, R.; Kaaya, R.; Wright, A.; West, P.A.; Kleinschmidt, I.; Kisinza, W.; Mosha, F.W.; et al. High Level of Resistance in the Mosquito Anopheles gambiae to Pyrethroid Insecticides and Reduced Susceptibility to Bendiocarb in North-Western Tanzania. Malar. J. 2013, 12, 149. [Google Scholar] [CrossRef] [Green Version]
- Ali Khan, H.A.A.; Akram, W.; Lee, S. Resistance to Selected Pyrethroid Insecticides in the Malaria Mosquito, Anopheles stephensi (Diptera: Culicidae), from Punjab, Pakistan. J. Med. Entomol. 2018, 55, 735–738. [Google Scholar] [CrossRef] [PubMed]
- Al-Sarar, A.S. Insecticide Resistance of Culex pipiens (L.) Populations (Diptera: Culicidae) from Riyadh City, Saudi Arabia: Status and Overcome. Saudi J. Biol. Sci. 2010, 17, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Khan, H.A.A.; Akram, W.; Shehzad, K.; Shaalan, E.A. First Report of Field Evolved Resistance to Agrochemicals in Dengue Mosquito, Aedes albopictus (Diptera: Culicidae), from Pakistan. Parasit. Vectors 2011, 4, 146. [Google Scholar] [CrossRef] [Green Version]
- Saddiq, B.; Shad, S.A.; Khan, H.A.A.; Aslam, M.; Ejaz, M.; Afzal, M.B.S. Resistance in the Mealybug Phenacoccus solenopsis Tinsley (Homoptera: Pseudococcidae) in Pakistan to Selected Organophosphate and Pyrethroid Insecticides. Crop Prot. 2014, 66, 29–33. [Google Scholar] [CrossRef]
- Geden, C.J. Status of Biopesticides for Control of House Flies. J. Biopest. 2012, 5, 1–11. [Google Scholar]
- Kavran, M.; Zgomba, M.F.; Ignjatovic-Cupina, A.M.; Lazic, S.D.; Petric, D.V. Choice of Optimal Biocide Combination to Control Flies (Diptera: Muscidae). Ann. Agric. Environ. Med. 2015, 22, 243–246. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, A.A.; Ahmed, F.A.; Kadhim, J.H.; Salman, A.M. Susceptibility of Adult and Larval Stages of Housefly, Musca domestica to Entomopathogenic Fungal biopesticides. Biocontrol Sci. Technol. 2021, 31, 1016–1026. [Google Scholar] [CrossRef]

{kind=link}
| Insecticide | Population | Year | 1 N | Slope ± SE | χ2 | p | 2 LC50 | 3 FL (95%) | 4 RR |
|---|---|---|---|---|---|---|---|---|---|
| Alpha-cypermethrin | Susceptible | - | 180 | 1.4 ± 0.3 | 0.8 | 0.8 | 42 | 29–61 | 1 |
| Dirab | 2019 | 180 | 2.4 ± 0.4 | 6.8 | 0.1 | 90 | 46–230 | 2 | |
| Al-Masanie | 2019 | 180 | 0.9 ± 0.3 | 0.6 | 0.9 | 160 | 93–557 | 4 | |
| Al-Uraija | 2019 | 180 | 1.2 ± 0.3 | 1.4 | 0.7 | 136 | 88–289 | 3 | |
| Al-Washlah | 2019 | 180 | 2.5 ± 0.5 | 2.3 | 0.5 | 89 | 71–114 | 2 | |
| Al-Muzahmiya | 2019 | 180 | 1.8 ± 0.3 | 2.6 | 0.5 | 86 | 64–119 | 2 | |
| Deltamethrin | Susceptible | - | 180 | 1.3 ± 0.3 | 0.6 | 0.4 | 71 | 48–116 | 1 |
| Dirab | 2019 | 180 | 0.9 ± 0.3 | 0.1 | 1.0 | 205 | 59–354 | 3 | |
| Al-Masanie | 2019 | 180 | 1.8 ± 0.3 | 1.8 | 0.6 | 889 | 667–1283 | 13 | |
| Al-Uraija | 2019 | 180 | 1.1 ± 0.3 | 0.7 | 0.9 | 322 | 170–497 | 5 | |
| Al-Washlah | 2019 | 180 | 2.4 ± 0.3 | 7.5 | 0.1 | 698 | 371–1698 | 10 | |
| Al-Muzahmiya | 2019 | 180 | 1.4 ± 0.3 | 1.3 | 0.7 | 398 | 262–566 | 6 | |
| Bifenthrin | Susceptible | - | 180 | 1.0 ± 0.3 | 0.2 | 0.7 | 139 | 87–265 | 1 |
| Dirab | 2019 | 180 | 1.5 ± 0.3 | 4.9 | 0.2 | 975 | 697–1570 | 7 | |
| Al-Masanie | 2019 | 180 | 1.1 ± 0.3 | 0.4 | 0.9 | 1638 | 984–4944 | 12 | |
| Al-Uraija | 2019 | 180 | 1.0 ± 0.3 | 0.7 | 0.9 | 651 | 340–3949 | 5 | |
| Al-Washlah | 2019 | 180 | 1.1 ± 0.3 | 0.7 | 0.9 | 552 | 342–926 | 4 | |
| Al-Muzahmiya | 2019 | 180 | 1.6 ± 0.3 | 2.7 | 0.4 | 1025 | 737–1643 | 7 | |
| Cyfluthrin | Susceptible | - | 180 | 1.6 ± 0.3 | 1.0 | 0.5 | 123 | 89–177 | 1 |
| Dirab | 2019 | 180 | 1.7 ± 0.3 | 5.7 | 0.1 | 580 | 428–800 | 5 | |
| Al-Masanie | 2019 | 180 | 1.2 ± 0.3 | 2.9 | 0.4 | 473 | 304–722 | 4 | |
| Al-Uraija | 2019 | 180 | 1.5 ± 0.3 | 5.1 | 0.2 | 490 | 345–690 | 4 | |
| Al-Washlah | 2019 | 180 | 0.8 ± 0.3 | 2.2 | 0.5 | 1107 | 605–4966 | 9 | |
| Al-Muzahmiya | 2019 | 180 | 0.8 ± 0.3 | 0.8 | 0.8 | 304 | 90–558 | 3 | |
| Cypermethrin | Susceptible | - | 180 | 1.3 ± 0.3 | 0.7 | 0.5 | 70 | 42–104 | 1 |
| Dirab | 2019 | 180 | 1.1 ± 0.3 | 2.0 | 0.6 | 211 | 82–341 | 3 | |
| Al-Masanie | 2019 | 180 | 2.0 ± 0.3 | 1.3 | 0.7 | 406 | 303–530 | 6 | |
| Al-Uraija | 2019 | 180 | 1.3 ± 0.3 | 0.5 | 0.9 | 404 | 257–591 | 6 | |
| Al-Washlah | 2019 | 180 | 1.3 ± 0.3 | 0.7 | 0.9 | 571 | 380–885 | 8 | |
| Al-Muzahmiya | 2019 | 180 | 1.1 ± 0.3 | 0.2 | 1.0 | 80 | 10–162 | 1 |
| Insecticide | Population | Year | 1 N | Slope ± SE | χ2 | p | 2 LC50 | 3 FL (95%) | 4 RR |
|---|---|---|---|---|---|---|---|---|---|
| Alpha-cypermethrin | Susceptible | - | 180 | 2.2 ± 0.3 | 3.6 | 0.9 | 35 | 19–56 | 1 |
| Dirab | 2019 | 180 | 2.2 ± 0.3 | 6.6 | 0.1 | 82 | 40–120 | 2 | |
| Al-Masanie | 2019 | 180 | 1.2 ± 0.3 | 4.5 | 0.2 | 241 | 146–709 | 7 | |
| Al-Uraija | 2019 | 180 | 0.8 ± 0.3 | 0.1 | 0.9 | 74 | 39–162 | 2 | |
| Al-Washlah | 2019 | 180 | 3.3 ± 0.4 | 2.6 | 0.5 | 59 | 49–71 | 2 | |
| Al-Muzahmiya | 2019 | 180 | 1.0 ± 0.3 | 2.1 | 0.5 | 19 | 5–33 | 0.5 | |
| Deltamethrin | Susceptible | - | 180 | 1.3 ± 0.3 | 0.5 | 0.2 | 47 | 31–69 | 1 |
| Dirab | 2019 | 180 | 0.8 ± 0.3 | 0.2 | 1.0 | 114 | 9–236 | 2 | |
| Al-Masanie | 2019 | 180 | 1.6 ± 0.3 | 1.6 | 0.6 | 983 | 704–1579 | 21 | |
| Al-Uraija | 2019 | 180 | 1.3 ± 0.3 | 0.3 | 0.9 | 133 | 48–215 | 3 | |
| Al-Washlah | 2019 | 180 | 1.3 ± 0.3 | 0.8 | 0.8 | 299 | 168–443 | 6 | |
| Al-Muzahmiya | 2019 | 180 | 1.3 ± 0.3 | 1.3 | 0.7 | 97 | 25–173 | 2 | |
| Bifenthrin | Susceptible | - | 180 | 1.2 ± 0.3 | 0.2 | 0.6 | 86 | 52–133 | 1 |
| Dirab | 2019 | 180 | 0.9 ± 0.3 | 4.1 | 0.3 | 1083 | 600–4376 | 13 | |
| Al-Masanie | 2019 | 180 | 1.7 ± 0.3 | 2.3 | 0.5 | 470 | 339–643 | 6 | |
| Al-Uraija | 2019 | 180 | 1.7 ± 0.4 | 2.3 | 0.5 | 510 | 343–1085 | 6 | |
| Al-Washlah | 2019 | 180 | 0.7 ± 0.3 | 0.3 | 1.0 | 1591 | 786–2088 | 19 | |
| Al-Muzahmiya | 2019 | 180 | 1.3 ± 0.3 | 0.8 | 0.8 | 829 | 568–1401 | 10 | |
| Cyfluthrin | Susceptible | - | 180 | 1.6 ± 0.3 | 3.1 | 0.6 | 85 | 43–154 | 1 |
| Dirab | 2019 | 180 | 1.1 ± 0.3 | 5.2 | 0.2 | 432 | 247–695 | 5 | |
| Al-Masanie | 2019 | 180 | 1.4 ± 0.3 | 1.2 | 0.8 | 345 | 215–496 | 4 | |
| Al-Uraija | 2019 | 180 | 1.6 ± 0.3 | 0.3 | 1.0 | 358 | 245–489 | 4 | |
| Al-Washlah | 2019 | 180 | 1.9 ± 0.3 | 4.9 | 0.2 | 355 | 244–672 | 4 | |
| Al-Muzahmiya | 2019 | 180 | 1.3 ± 0.3 | 0.3 | 1.0 | 121 | 38–204 | 1 | |
| Cypermethrin | Susceptible | - | 180 | 1.6 ± 0.3 | 1.9 | 0.1 | 53 | 34–73 | 1 |
| Dirab | 2019 | 180 | 0.9 ± 0.3 | 0.2 | 1.0 | 74 | 4–166 | 1 | |
| Al-Masanie | 2019 | 180 | 0.9 ± 0.3 | 0.2 | 1.0 | 72 | 4–161 | 1 | |
| Al-Uraija | 2019 | 180 | 1.5 ± 0.3 | 1.3 | 0.7 | 201 | 108–292 | 4 | |
| Al-Washlah | 2019 | 180 | 0.8 ± 0.3 | 0.9 | 0.8 | 175 | 28–329 | 3 | |
| Al-Muzahmiya | 2019 | 180 | 2.9 ± 0.6 | 1.3 | 0.7 | 122 | 71–162 | 2 |
| Insecticide | Population | Year | 1 N | Slope ± SE | χ2 | p | 2 LC50 | 3 FL (95%) | 4 RR |
|---|---|---|---|---|---|---|---|---|---|
| Fenitrothion | Susceptible | - | 180 | 1.0 ± 0.3 | 1.8 | 0.6 | 37 | 19–61 | 1 |
| Dirab | 2019 | 180 | 1.3 ± 0.3 | 2.0 | 0.6 | 849 | 587–1418 | 23 | |
| Al-Masanie | 2019 | 180 | 2.0 ± 0.3 | 6.6 | 0.1 | 548 | 241–1386 | 15 | |
| Al-Uraija | 2019 | 180 | 1.5 ± 0.3 | 0.7 | 0.9 | 410 | 279–575 | 11 | |
| Al-Washlah | 2019 | 180 | 1.1 ± 0.3 | 0.9 | 0.8 | 241 | 104–381 | 7 | |
| Al-Muzahmiya | 2019 | 180 | 1.9 ± 0.3 | 1.7 | 0.6 | 990 | 749–1425 | 27 | |
| Chlorpyrifos | Susceptible | - | 180 | 1.8 ± 0.3 | 3.9 | 0.3 | 26 | 9–46 | 1 |
| Dirab | 2019 | 180 | 1.1 ± 0.3 | 1.6 | 0.7 | 302 | 146–475 | 12 | |
| Al-Masanie | 2019 | 180 | 1.2 ± 0.3 | 0.7 | 0.9 | 50 | 9–106 | 2 | |
| Al-Uraija | 2019 | 180 | 1.1 ± 0.3 | 0.4 | 0.9 | 347 | 189–537 | 13 | |
| Al-Washlah | 2019 | 180 | 1.7 ± 0.4 | 1.3 | 0.7 | 120 | 52–145 | 5 | |
| Al-Muzahmiya | 2019 | 180 | 1.5 ± 0.3 | 0.4 | 0.7 | 352 | 231–494 | 14 | |
| Malathion | Susceptible | - | 180 | 2.4 ± 0.3 | 0.5 | 0.1 | 79 | 52–144 | 1 |
| Dirab | 2019 | 180 | 1.5 ± 0.3 | 0.9 | 0.8 | 267 | 160–380 | 3 | |
| Al-Masanie | 2019 | 180 | 1.2 ± 0.3 | 0.9 | 0.8 | 266 | 141–399 | 3 | |
| Al-Uraija | 2019 | 180 | 1.5 ± 0.3 | 2.7 | 0.4 | 375 | 249–525 | 5 | |
| Al-Washlah | 2019 | 180 | 1.9 ± 0.3 | 1.3 | 0.7 | 736 | 555–1023 | 9 | |
| Al-Muzahmiya | 2019 | 180 | 1.3 ± 0.3 | 5.1 | 0.2 | 680 | 468–1071 | 9 |
| Insecticide | Population | Year | 1 N | Slope ± SE | χ2 | p | 2 LC50 | 3 FL (95%) | 4 RR |
|---|---|---|---|---|---|---|---|---|---|
| Fenitrothion | Susceptible | - | 180 | 1.9 ± 0.3 | 4.6 | 0.2 | 32 | 13–60 | 1 |
| Dirab | 2019 | 180 | 1.9 ± 0.3 | 4.4 | 0.2 | 280 | 104–490 | 9 | |
| Al-Masanie | 2019 | 180 | 2.3 ± 0.3 | 2.8 | 0.4 | 421 | 325–536 | 13 | |
| Al-Uraija | 2019 | 180 | 1.2 ± 0.3 | 1.6 | 0.7 | 139 | 44–234 | 4 | |
| Al-Washlah | 2019 | 180 | 1.1 ± 0.2 | 1.8 | 0.6 | 466 | 322–659 | 15 | |
| Al-Muzahmiya | 2019 | 180 | 3.3 ± 0.4 | 6.6 | 0.1 | 444 | 255–799 | 14 | |
| Chlorpyrifos | Susceptible | - | 180 | 1.7 ± 0.3 | 5.6 | 0.1 | 18 | 10–25 | 1 |
| Dirab | 2019 | 180 | 1.7 ± 0.3 | 1.3 | 0.7 | 195 | 115–272 | 11 | |
| Al-Masanie | 2019 | 180 | 1.6 ± 0.3 | 1.4 | 0.7 | 127 | 56–192 | 7 | |
| Al-Uraija | 2019 | 180 | 1.9 ± 0.3 | 5.4 | 0.2 | 259 | 159–486 | 14 | |
| Al-Washlah | 2019 | 180 | 3.6 ± 0.4 | 0.2 | 0.9 | 93 | 32–125 | 5 | |
| Al-Muzahmiya | 2019 | 180 | 1.6 ± 0.3 | 2.9 | 0.4 | 236 | 141–332 | 13 | |
| Malathion | Susceptible | - | 180 | 1.6 ± 0.3 | 0.4 | 0.9 | 46 | 33–65 | 1 |
| Dirab | 2019 | 180 | 2.1 ± 0.3 | 1.2 | 0.8 | 219 | 149–290 | 5 | |
| Al-Masanie | 2019 | 180 | 2.1 ± 0.4 | 0.3 | 0.9 | 121 | 61–173 | 3 | |
| Al-Uraija | 2019 | 180 | 1.3 ± 0.3 | 0.3 | 0.9 | 157 | 65–246 | 3 | |
| Al-Washlah | 2019 | 180 | 1.5 ± 0.3 | 2.0 | 0.6 | 542 | 385–772 | 12 | |
| Al-Muzahmiya | 2019 | 180 | 2.1 ± 0.3 | 2.8 | 0.4 | 385 | 292–495 | 8 |
| Population | Conc. ppm | 1 LT50 (h) | 2 FL (95%) | Slope (SE) | Conc. ppm | 1 LT50 (h) | 2 FL (95%) | Slope (SE) |
|---|---|---|---|---|---|---|---|---|
| Alpha-cypermethrin | Deltamethrin | |||||||
| Dirab | 256 | 14 | 12–17 b | 5.8 (1.2) | 2048 | 3 | 1–4 c | 1.2 (0.2) |
| Al-Masanie | 256 | 29 | 19–52 a | 1.4 (0.3) | 2048 | 22 | 15–33 a | 1.7 (0.4) |
| Al-Uraija | 256 | 33 | 21–68 a | 1.3 (0.3) | 2048 | 5 | 3–8 bc | 1.3 (0.2) |
| Al-Washlah | 256 | 9 | 6–13 bc | 2.7 (0.5) | 2048 | 9 | 6–14 b | 1.1 (0.2) |
| Al-Muzahmiya | 256 | 3 | 1–7 c | 2.0 (0.7) | 2048 | 3 | 1–6 bc | 1.1 (0.2) |
| Bifenthrin | Cypermethrin | |||||||
| Dirab | 2048 | 11 | 6–18 b | 1.2 (0.2) | 2048 | 3 | 1–6 ab | 1.0 (0.2) |
| Al-Masanie | 2048 | 8 | 5–13 b | 1.3 (0.2) | 2048 | 1 | 0–3 ab | 0.8 (0.2) |
| Al-Uraija | - | - | - | - | 2048 | 7 | 3–13 a | 0.9 (0.2) |
| Al-Washlah | 2048 | 59 | 28–461 a | 0.8 (0.2) | 2048 | 3 | 0–7 ab | 0.7 (0.2) |
| Al-Muzahmiya | 2048 | 17 | 10–29 ab | 1.2 (0.2) | 2048 | 1 | 0–2 b | 1.1 (0.2) |
| Cyfluthrin | Fenitrothion | |||||||
| Dirab | 2048 | 1 | 0–3 a | 0.8 (0.2) | 2048 | 14 | 9–17 ab | 3.8 (0.8) |
| Al-Masanie | 2048 | 0.3 | 0–2 a | 0.4 (0.2) | 2048 | 13 | 8–17 ab | 2.3 (0.5) |
| Al-Uraija | 2048 | 0.4 | 0–2 a | 0.6 (0.2) | 2048 | 6 | 3–10 b | 1.3 (0.2) |
| Al-Washlah | 2048 | 0.6 | 0–2 a | 0.8 (0.2) | 2048 | 16 | 11–20 a | 2.5 (0.6) |
| Al-Muzahmiya | 2048 | 1 | 0–2 a | 1.0 (0.2) | 2048 | 13 | 9–16 ab | 3.5 (0.8) |
| Chlorpyrifos | Malathion | |||||||
| Dirab | 2048 | 9 | 6–12 a | 2.6 (0.5) | - | - | - | - |
| Al-Masanie | - | - | - | - | 2048 | 8 | 5–11 bc | 2.6 (0.5) |
| Al-Uraija | 2048 | 11 | 7–15 a | 2.6 (0.5) | 2048 | 12 | 7–15 ab | 3.0 (0.7) |
| Al-Washlah | 2048 | 2 | 1–2 b | 2.2 (0.4) | 2048 | 18 | 12–23 a | 2.4 (0.5) |
| Al-Muzahmiya | 2048 | 3 | 0–5 b | 0.9 (0.2) | 2048 | 3 | 1–5 c | 1.1 (0.2) |
| Population | Conc. ppm | 1 LT50 (h) | 2 FL (95%) | Slope (SE) | Conc. ppm | 1 LT50 (h) | 2 FL (95%) | Slope (SE) |
|---|---|---|---|---|---|---|---|---|
| Alpha-cypermethrin | Deltamethrin | |||||||
| Dirab | 256 | 10 | 5–13 bc | 3.6 (1.0) | 2048 | 4 | 2–8 c | 1.1 (0.2) |
| Al-Masanie | 256 | 13 | 5–42 abc | 0.6 (0.2) | 2048 | 4 | 1–7 c | 1.0 (0.2) |
| Al-Uraija | 256 | 30 | 18–71 a | 1.1 (0.3) | 2048 | 17 | 11–26 ab | 1.4 (0.3) |
| Al-Washlah | 256 | 19 | 13–25 ab | 2.2 (0.5) | 2048 | 24 | 18–35 a | 1.5 (0.2) |
| Al-Muzahmiya | 256 | 3 | 0–7 c | 0.6 (0.2) | 2048 | 6 | 2–15 bc | 0.7 (0.2) |
| Bifenthrin | Cypermethrin | |||||||
| Dirab | 2048 | 5 | 3–9 b | 1.3 (0.2) | 2048 | 1 | 0–4 a | 0.5 (0.2) |
| Al-Masanie | 2048 | 49 | 24–262 a | 0.8 (0.2) | 2048 | 4 | 1–7 a | 0.9 (0.2) |
| Al-Uraija | 2048 | - | - | - | 2048 | 3 | 1–8 a | 0.8 (0.4) |
| Al-Washlah | 2048 | 14 | 8–27 ab | 1.0 (0.2) | 2048 | 4 | 1–11 a | 0.26 (0.2) |
| Al-Muzahmiya | 2048 | 17 | 10–31 a | 1.1 (0.2) | 2048 | 1 | 0.1–3 a | 0.7 (0.2) |
| Cyfluthrin | Fenitrothion | |||||||
| Dirab | 2048 | 4 | 2–7 a | 1.4 (0.2) | 2048 | 36 | 20–113 a | 0.9 (0.2) |
| Al-Masanie | 2048 | 2 | 0–5 a | 0.7 (0.2) | 2048 | 22 | 18–27 a | 3.5 (0.7) |
| Al-Uraija | 2048 | 2 | 0–4 a | 0.9 (0.2) | 2048 | 14 | 8–23 a | 1.2 (0.2) |
| Al-Washlah | 2048 | 7 | 0–31 a | 0.4 (0.2) | 2048 | 20 | 15–25 a | 3.1 (0.6) |
| Al-Muzahmiya | 2048 | 6 | 1–13 a | 0.7 (0.2) | 2048 | 24 | 17–35 a | 1.9 (0.4) |
| Chlorpyrifos | Malathion | |||||||
| Dirab | 2048 | 10 | 7–14 b | 2.0 (0.4) | 2048 | 8 | 5–11 c | 2.3 (0.4) |
| Al-Masanie | 2048 | 8 | 5–11 b | 2.8 (0.6) | - | - | - | - |
| Al-Uraija | 2048 | 21 | 16–27 a | 2.6 (0.6) | 2048 | 15 | 10–21 bc | 1.8 (0.3) |
| Al-Washlah | 2048 | 3 | 2–4 c | 1.9 (0.3) | 2048 | 22 | 16–31 ab | 2.2 (0.5) |
| Al-Muzahmiya | 2048 | 14 | 9–21 ab | 1.6 (0.3) | 2048 | 34 | 27–48 a | 2.8 (0.6) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hafez, A.M. First Evaluation of Field Evolved Resistance to Commonly Used Insecticides in House Fly Populations from Saudi Arabian Dairy Farms. Insects 2021, 12, 1120. https://doi.org/10.3390/insects12121120
Hafez AM. First Evaluation of Field Evolved Resistance to Commonly Used Insecticides in House Fly Populations from Saudi Arabian Dairy Farms. Insects. 2021; 12(12):1120. https://doi.org/10.3390/insects12121120
Chicago/Turabian StyleHafez, Abdulwahab M. 2021. "First Evaluation of Field Evolved Resistance to Commonly Used Insecticides in House Fly Populations from Saudi Arabian Dairy Farms" Insects 12, no. 12: 1120. https://doi.org/10.3390/insects12121120
APA StyleHafez, A. M. (2021). First Evaluation of Field Evolved Resistance to Commonly Used Insecticides in House Fly Populations from Saudi Arabian Dairy Farms. Insects, 12(12), 1120. https://doi.org/10.3390/insects12121120