Hymenoptera Complex Associated with Myzus persicae and Hyalopterus spp. in Peach Orchards in Northeastern Spain and Prospects for Biological Control of Aphids

 , ,
, ,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Species Identification of M. persicae Parasitoids and Hyperparasitoids
2.2. Species Identification of Hyalopterus spp. Parasitoids and Hyperparasitoids
3. Results
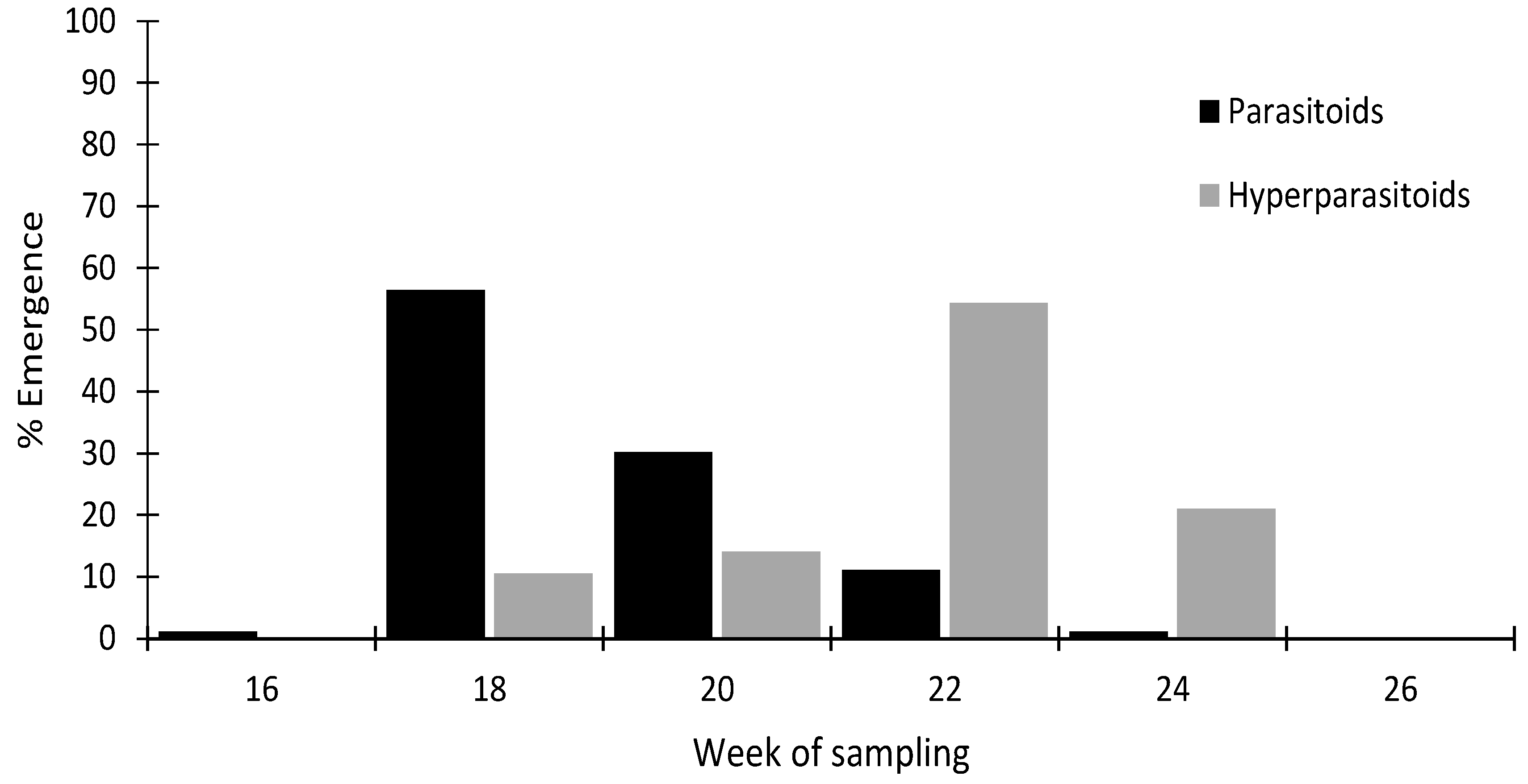
3.1. Species Identification of M. persicae Parasitoids and Hyperparasitoids
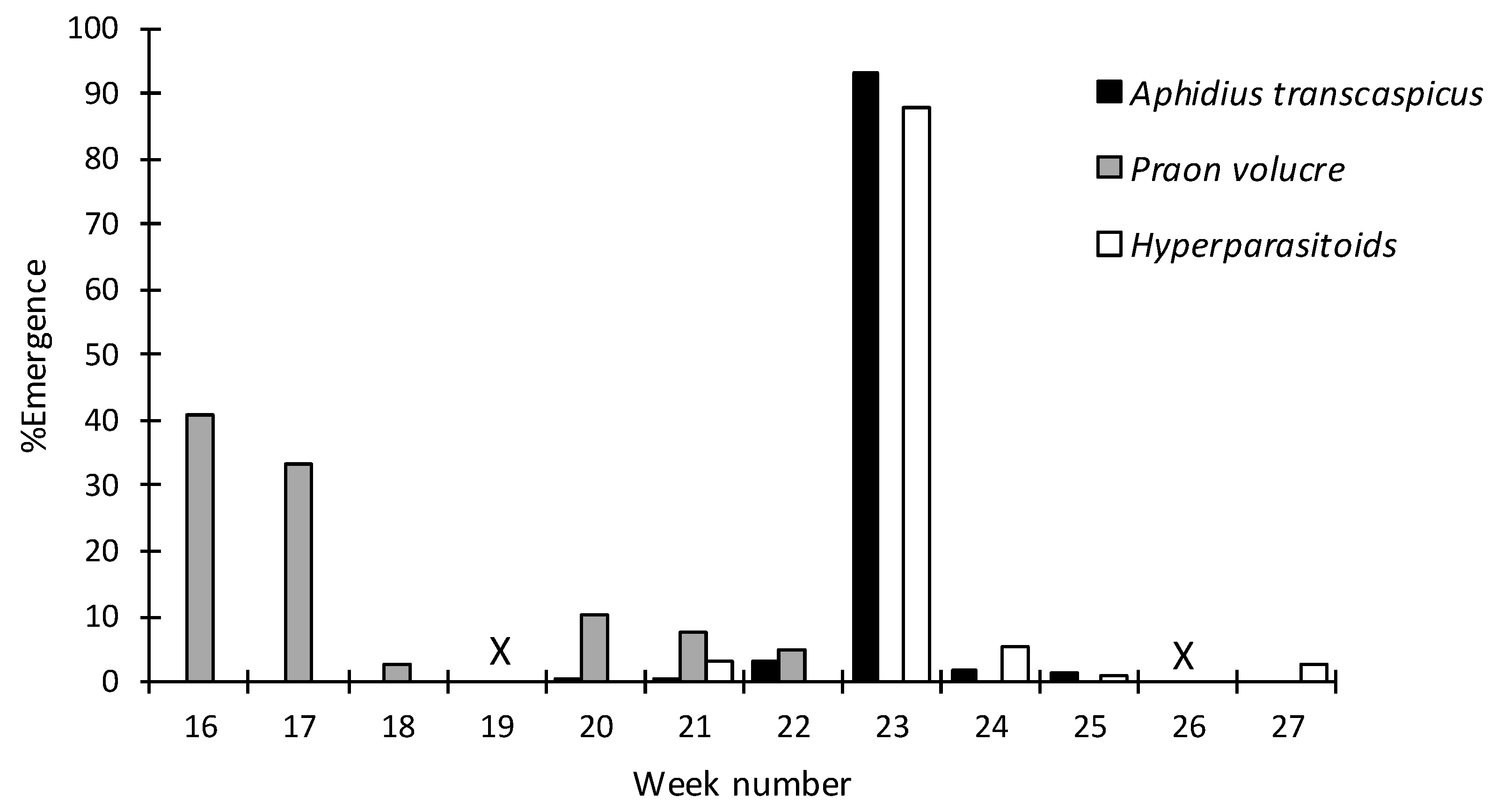
3.2. Species Identification of Hyalopterus spp. Parasitoids and Hyperparasitoids
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization Statistics (FAOSTAT): Crops. 2017. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 12 January 2019).
- Ministerio de Agricultura, Pesca y Alimentación (MAPA). 2017. Available online: https://www.mapa.gob.es/es/estadistica/temas/estadisticas-agrarias/agricultura/superficies-producciones-anuales-cultivos/ (accessed on 2 March 2019).
- Alhmedi, A.; Raymaekers, S.; Tomanović, Ž.; Bylemans, D.; Belien, T. Food web structure of aphids and their parasitoids in Belgian fruit agroecosystems. Entomol. Sci. 2018, 21, 279–291. [Google Scholar] [CrossRef]
- Kavallieratos, N.G.; Tomanović, Ž.; Starý, P.; Bogdanović-Mitrovski, A. Parasitoids (Hymenoptera: Braconidae: Aphidiinae) attacking aphids feeding on Prunoideae and Maloideae crops in Southeast Europe: Aphidiine-aphid-plant associations and key. Zootaxa 2008, 1793, 47–64. [Google Scholar] [CrossRef]
- Basky, Z. Predators and parasites of Hyalopterus pruni and Hyalopterus amygdali populations living on peach, plum and reed. Acta Phytopathol. Acad. Sci. Hung. 1982, 17, 311–316. [Google Scholar]
- Blackman, R.L.; Eastop, V.F. Aphids on the World’s Crops, 2nd ed.; John Wiley & Sons: Chichester, UK, 2000; ISBN 9780471851912. [Google Scholar]
- Dedryver, C.A.; Le Ralec, A.; Fabre, F. The conflicting relationships between aphids and men: A review of aphid damage and control strategies. C. R. Biol. 2010, 333, 539–553. [Google Scholar] [CrossRef] [PubMed]
- Barbagallo, S.; Cocuzza, G.E.M.; Cravedi, P.; Komazaki, S. IPM case studies: Deciduous fruit trees aphids. In Aphids as Crop Pests, 2nd ed.; Emden, H.F., van Harrington, R., Eds.; CABI: Wallingford, UK, 2017; pp. 632–642. ISBN 9781780647098. [Google Scholar]
- Isac, M.; Preda, S.; Marcu, M. Aphid species—Vectors of plum pox virus. Acta Virol. 1998, 42, 233–234. [Google Scholar] [PubMed]
- Penvern, S.; Bellon, S.; Fauriel, J.; Sauphanor, B. Peach orchard protection strategies and aphid communities: Towards an integrated agroecosystem approach. Crop Prot. 2010, 29, 1148–1156. [Google Scholar] [CrossRef]
- Foster, S.P.; Denholm, I.; Harling, Z.K.; Moores, G.D. Intensification of insecticide resistance in UK field populations of the peach-potato aphid, Myzus persicae (Hemiptera: Aphididae) in 1996. Bull. Entomol. Res. 1998, 88, 127–130. [Google Scholar] [CrossRef]
- Slater, R.; Paul, V.L.; Andrews, M.; Garbay, M.; Camblin, P. Identifying the presence of neonicotinoid-resistant peach-potato aphid (Myzus persicae) in the peach-growing regions of southern France and northern Spain. Pest Manag. Sci. 2012, 68, 634–638. [Google Scholar] [CrossRef]
- Angalet, G.W.; Fuester, R. The Aphidius parasites of the pea aphid Acyrthosiphon pisum in the eastern half of the United States. Ann. Entomol. Soc. Am. 1977, 70, 87–96. [Google Scholar] [CrossRef]
- Starý, P.; Lyon, J.P.; Lecant, F. Biocontrol of aphids by the introduced Lysiphlebus testaceipes (Ceresson) (Hym: Aphidiidae) in Mediterranean France. J. Appl. Entomol. 1988, 105, 74–87. [Google Scholar] [CrossRef]
- Thies, C.; Roschewitz, I.; Tscharntke, T. The landscape context of cereal aphid–parasitoid interactions. Proc. R. Soc. 2005, 272, 203–210. [Google Scholar] [CrossRef]
- Brodeur, J.; McNeil, J.N. Life history of the aphid hyperparasitoid Asaphes vulgaris Walker (Pteromalidae): Possible consequences on the efficacy of the primary parasitoid Aphidius nigripes Ashmead (Aphidiidae). Can. Entomol. 1994, 126, 1493–1497. [Google Scholar] [CrossRef]
- Rosenheim, J.A. Higher-order predators and the regulation of insect herbivore populations. Ann. Rev. Entomol. 1998, 43, 421–447. [Google Scholar] [CrossRef]
- Sampaio, M.V.; Korndörfer, A.P.; Pujade-Villar, J.; Hubaide, J.E.A.; Ferreira, S.E.; Arantes, S.O.; Bortoletto, D.M.; Guimarães, C.M.; Sánchez-Espigares, J.A.; Caballero-López, B. Brassica aphid (Hemiptera: Aphididae) populations are conditioned by climatic variables and parasitism level: A study case of Triângulo Mineiro, Brazil. Bull. Entomol. Res. 2017, 107, 410–418. [Google Scholar] [CrossRef]
- Schooler, S.S.; De Barro, P.; Ives, A.R. The potential for hyperparasitism to compromise biological control: Why don’t hyperparasitoids drive their primary parasitoid hosts extinct? Biol. Control 2011, 32, 172–179. [Google Scholar] [CrossRef]
- Sullivan, D.J. Insect hyperparasitism. Ann. Rev. Entomol. 1987, 32, 49–70. [Google Scholar] [CrossRef]
- Beddington, J.R.; Hammond, P.S. On the dynamics of host-parasite–hyperparasite interactions. J. Anim. Ecol. 1977, 46, 811–821. [Google Scholar] [CrossRef]
- de Vere Graham, M.W.R. The Pteromalidae of north-western Europe (Hymenoptera: Chalcidoidea). Bull. Br. Mus. Nat. Hist. Zool. 1969, 16, 908. [Google Scholar]
- Kamijo, K.; Takada, H. Studies on aphid hyperparasites of Japan. II. Aphid hyperparasites of the Pteromalidae occurring in Japan (Hymenoptera). Insecta Matsumurana 1973, 2, 39–76. [Google Scholar]
- Rakshani, E.; Kazemzadeh, S.; Starý, P.; Barahoei, H.; Kavallieratos, N.G.; Ćetković, A.; Popović, A.; Bodlah, I.; Tomanović, Ž. Parasitoids (Hymenoptera: Braconidae: Aphidiinae) of northeastern Iran: Aphidiine-aphid-plant associations, key and description of a new species. J. Insect Sci. 2012, 12, 143. [Google Scholar] [CrossRef] [PubMed]
- Rakshani, E.; Starý, P.; Tomanović, Ž.; Mifsud, D. Aphidiinae (Hymenoptera, Braconidae) aphid parasitoids of Malta: Review and key to species. Bull. Entomol. Soc. Malta 2015, 7, 121–137. [Google Scholar] [CrossRef]
- Kavallieratos, N.G.; Tomanović, Ž.; Petrović, A.; Janković, M.; Starý, P.; Yovkova, M.; Athanassiou, C.G. Review and Key for the Identification of Parasitoids (Hymenoptera: Braconidae: Aphidiinae) of Aphids Infesting Herbaceous and Shrubby Ornamental Plants in Southeastern Europe. Ann. Entomol. Soc. Am. 2013, 106, 294–309. [Google Scholar] [CrossRef]
- Ghaliow, M.E.; Petrović, A.; Kocić, K.; ČkrkiĆ, J.; Bogdanović, A.M.; Starý, P.; Kavallieratos, N.G.; Tomanović, Ž. Key for identification of the parasitoids (Hymenoptera: Braconidae: Aphidiinae) of aphids infesting alfalfa in Europe. Zootaxa 2018, 4378, 98–110. [Google Scholar] [CrossRef]
- Lumbierres, B.; Starý, P.; Pons, X. Seasonal parasitism of cereal aphids in a Mediterranean arable crop system. J. Pest Sci. 2007, 80, 125–130. [Google Scholar] [CrossRef]
- Mackauer, M. Insect parasites of the green peach aphid, Myzus persicae Sulz., and their control potential. Entomophaga 1968, 13, 91–106. [Google Scholar] [CrossRef]
- Remaudière, G.; Leclant, F. Le complexe des ennemis naturels des aphides du pêcher dans la moyenne vallée du Rhône. Entomophaga 1971, 16, 255–267. [Google Scholar] [CrossRef]
- Starý, P.; Remaudière, G.; Leclant, F. Les Aphidiidae (Hymenoptera) de France et leurs hôtes (Homoptera: Aphididae). Entomophaga Mémoire Hors Série 1971, 5, 1–74. [Google Scholar]
- Michelena, J.M.; Sanchís, A.; González, P. Afidíinos sobre pulgones de frutales en la Comunidad Valenciana. Bol. San. Veg. Plagas 1994, 20, 465–470. [Google Scholar]
- Michelena, J.M.; González, P.; Soler, E. Parasitoides afidiinos (Hymenoptera, Braconidae, Aphidiinae) de pulgones de cultivos agrícolas en la Comunidad Valenciana. Bol. San. Veg. Plagas 2004, 30, 317–326. [Google Scholar]
- Laamari, M.; Khenissa, N.; Merouani, H.; Ghodbane, S.; Starý, P. Importance des Hyménoptères prasitoïdes des pucerons en Algérie. In Proceedings of the Colloque International sur la Gestion des Risques Phytosanitaires, Marrakech, Morocco, 9–11 November 2009; pp. 581–587. [Google Scholar]
- Pennacchio, F. The Italian species of the genus Aphidius Nees (Hymenoptera: Braconidae: Aphidiinae). Boll. Lab. Entomol. Agraria ‘Filippo Silvestri’ 1989, 46, 75–106. [Google Scholar]
- Boivin, G.; Hance, T.; Brodeur, J. Aphid parasitoids in biological control. Can. J. Plant Sci. 2012, 92, 1–12. [Google Scholar] [CrossRef]
- Pons, X.; Comas, J.; Albajes, R. Overwintering of cereal aphids (Homoptera: Aphididae) on durum wheat in a Mediterranean climate. Environ. Entomol. 1993, 22, 381–387. [Google Scholar] [CrossRef]
- Pons, X.; Starý, P. Spring aphid-parasitoid (Hom., Aphididae, Hym., Braconidae) associations and interactions in a Mediterranean arable crop ecosystem, including Bt maize. J. Pest Sci. 2003, 76, 133–138. [Google Scholar] [CrossRef]
- Starý, P. A review of Aphidius species (Hymenoptera: Aphidiidae) of Europe. Annot. Zool. Bot. Bratislava 1973, 85, 1–85. [Google Scholar]
- van Lenteren, J.C. The state of commercial augmentative biological control: Plenty of natural enemies, but a frustrating lack of uptake. BioControl 2012, 57, 1–20. [Google Scholar] [CrossRef]
- Kocić, K.; Tomanović, Z. Faculty of Biology, University of Belgrade, Belgrade, Serbia. Unpublished Data. 2018. [Google Scholar]
- Kos, K.; Trdan, S.; Petrović, A.; Starý, P.; Kavallieratos, N.G.; Petrović-Obradović, O.; Tomanović, Ž. Aphidiinae (Hymenoptera, Braconidae, Aphidiinae) from Slovenia, with description of a new Aphidius species. Zootaxa 2012, 3456, 36–50. [Google Scholar] [CrossRef]
- Starý, P. A review of the parasites of aphids associated with Prunus trees in Czechoslovakia (Hymenoptera: Aphidiidae, Homoptera: Aphidoidea). Acta Entomol. Bohemoslov. 1966, 63, 67–75. [Google Scholar]
- Starý, P. Aphid Parasitoids of the Czech Republic (Hymenoptera: Braconidae: Aphidiinae); Academia: Prague, Czech Republic, 2006. [Google Scholar]
- Starý, P.; Lukáš, J. Aphid parasitoids and their tritrophic associations in Slovakia (Hymenoptera: Braconidae: Aphidiinae). Folia Hymenopterol. Slovaca 2009, 1, 63. [Google Scholar]
- Starý, P. Parasites (Hymenoptera, Aphidiidae) of leaf-curling apple aphids in Czechoslovakia. Acta Entomol. Bohemoslov. 1975, 72, 99–114. [Google Scholar]
- Adisu, B.; Starý, P.; Freier, B.; Büttner, C. Aphidius colemani Vier. (Hymenoptera, Braconidae, Aphidiinae) detected in cereal fields in Germany. J. Pest. Sci. 2002, 75, 89–94. [Google Scholar] [CrossRef]
- Starý, P. Field establishment of Aphidius colemani Vier. (Hym., Braconidae, Aphidiinae) in the Czech Republic. J. Appl. Entomol. 2002, 126, 405–408. [Google Scholar] [CrossRef]
- El-Mali, M.U.; Starý, P.; Shahbaz, F.; Özsemercl, F. A review of Aphid parasitoids (Hym. Braconidae, Aphidiinae) of Turkey. Egypt. J. Biol. Pest Control 2004, 14, 355–370. [Google Scholar] [CrossRef]
- Gärdenfors, U. Taxonomic and biological revision of Palearctic Ephedrus Haliday (Hymenoptera: Braconidae: Aphidiinae). Entomol. Scand. 1986, 27, 1–95. [Google Scholar]
- Rothschild, G.H.L. Records of hymenopterous parasites and hyperparasites of certain fruit-tree aphids. Entomol. Mon. Mag. 1963, 99, 126–127. [Google Scholar]
- Starý, P. A study on the relationship of the Anuraphidina and their aphidiid parasites in Europe (Homoptera, Aphidoidea; Hymenoptera, Aphidiidae). Beitr. Entomol. 1969, 19, 665–671. [Google Scholar]
- Kavallieratos, N.G.; Lykouressis, D.P.; Sarlis, G.P.; Stathas, G.J.; Segovia, A.S.; Athanassiou, C.G. The aphidiinae (Hym: Ichneumonidea: Braconidae) of Greece. Phytoparasitica 2001, 29, 306–340. [Google Scholar] [CrossRef]
- Wang, X.; Messing, R. Potential host range of newly introduced aphid parasitoid Aphidius transcaspicus (Hymenoptera: Braconidae) in Hawaii. Proc. Hawai. Entomol. Soc. 2006, 38, 81–86. [Google Scholar]
- Lozier, J.D.; Roderick, G.K.; Mills, N.J. Evolutionarily significant units in natural enemies: Identifying regional populations of Aphidius transcaspicus (Hymenoptera: Braconidae) for use in biological control of mealy plum aphid. Biol. Control 2008, 46, 532–541. [Google Scholar] [CrossRef]
- Starý, P. Integrated control of the mealy plum aphid Hyalopterus pruni (Geoffr.) in Czechoslovakia (Hom. Aphidiidae). Ann. Soc. Entomol. Fr. 1965, 1, 177–180. [Google Scholar]
- González, F.P.; Michelena, J.M. Relaciones parasitoide-pulgón (Hymenoptera: Aphidiinae; Homoptera:Aphididae) en la provincia de Alicante. Bol. Asoc. Esp. Entomol. 1987, 11, 245–258. [Google Scholar]
- Starý, P. Parasite spectrum and relative abundance of parasites of cereal aphids in Czechoslovakia (Hymenoptera, Aphidiidae; Homoptera, Aphidoidea). Acta Entomol. Bohemoslov. 1976, 73, 216–223. [Google Scholar]
- Tomanović, Ž.; Brajkovic, M. Aphid parasitoids (Hymenoptera: Aphidiidae) of agroecosystems of the south part of the Pannonian area. Arch. Biol. Sci. 2001, 53, 57–64. [Google Scholar]
- Ferrer-Suay, M.; Selfa, J.; Ribes, A.; Pujade-Villar, J. Contribucions al coneixement dels Charipins de Catalunya (Insecta, Hymenoptera). Orsis 2012, 26, 113–134. [Google Scholar]
- Ferrer-Suay, M.; Selfa, J.; Ribes, A.; Pujade-Villar, J. A key of the Charipinae (Hymenoptera, Cynipoidea, Figitidae) from Spain, including new records and species. Bol. Asoc. Esp. Entomol. 2013, 37, 315–341. [Google Scholar]
- Gómez-Marco, F.; Urbaneja, A.; Jacas, J.A.; Rugman-Jones, P.; Stouthamer, R.; Tena, A. Untangling the aphid-parasitoid food web in citrus: Can hyperparasitoids disrupt biological control? Biol. Control 2015, 81, 111–121. [Google Scholar] [CrossRef]
- Suay-Cano, V.A.; Martínez, F.L.; Michelena, J.M. Parasitoides no afidíinos de pulgones (Chalcidoidea: Aphelinidae) e hiperparasitoides de las superfamilias Chalcidoidea, Ceraphronoidea y Cynipoidea (Hymenoptera: Apócrifa: Parastitica) en la provincia de Valencia. Bol. Asoc. Esp. Entomol. 1998, 22, 99–113. [Google Scholar]
- Acheampong, S.; Gillespie, D.R.; Quiring, D. Survey of parasitoids and hyperparasitoids (Hymenoptera) of the green peach aphid, Myzus persicae and the foxglove aphid, Aulacorthum solani (Hemiptera: Aphididae) in British Columbia. J. Entomol. Soc. Br. Columbia 2012, 109, 12–22. [Google Scholar]
- Sullivan, D.J. Comparative behaviour and competition between two aphid hyperparasites: Alloxysta victrix and Asaphes californicus (Hymenoptera: Cynipidae; Pteromalidae). Environ. Entomol. 1972, 1, 234–244. [Google Scholar] [CrossRef]
- Kanuck, M.J.; Sullivan, D.J. Ovipositional Behavior and Larval Development of Aphidencyrtus aphidivorus (Hymenoptera: Encyrtidae), an Aphid Hyperparasitoid. J. N. Y. Entomol. Soc. 1992, 100, 527–532. [Google Scholar]
- van Lenteren, J.C. Quality Control and Production of Biological Control Agents: Theory and Testing Procedures; CABI: Wallingford, Oxon, UK, 2003; ISBN 0851996884. [Google Scholar]
- Alignier, A.; Raymond, L.; Deconchat, M.; Menozzi, P.; Monteil, C.; Sarthou, J.P.; Vialatte, A.; Ouin, A. The effect of semi-natural habitats on aphids and their natural enemies across spatial and temporal scales. Biol. Control 2014, 77, 76–82. [Google Scholar] [CrossRef]
- Blaauw, B.R.; Isaacs, R. Wildflower plantings enhance the abundance of natural enemies and their services in adjacent blueberry fields. Biol. Control 2015, 91, 94–103. [Google Scholar] [CrossRef]
- Gagić, V.; Hänke, S.; Thies, C.; Scherber, C.; Tomanović, Ž.; Tscharntke, T. Agricultural intensification and cereal aphid–parasitoid–hyperparasitoid food webs: Network complexity, temporal variability and parasitism rates. Oecologia 2012, 170, 1099–1109. [Google Scholar] [CrossRef] [PubMed]
- Tomanović, Ž.; Kavallieratos, N.G.; Starý, P.; Stanisavljević, L.Z.; Cetković, A.; Stamenković, S.; Jovanović, S.; Athanassiou, C.G. Regional tritrophic relationship patterns of five aphid parasitoid species (Hymenoptera: Braconidae: Aphidiinae) in agroecosystem-dominated landscapes of southeastern Europe. J. Econ. Entomol. 2009, 102, 836–854. [Google Scholar] [CrossRef]
- Vollhardt, I.M.G.; Bianchi, F.J.J.A.; Wäckers, F.L.; Thies, C.; Tscharntke, T. Spatial distribution of flower vs. honeydew resources in cereal fields may affect aphid parasitism. Biol. Control 2010, 53, 204–213. [Google Scholar] [CrossRef]
- Aparicio, Y.; Gabarra, R.; Arnó, J. Attraction of Aphidius ervi (Hymenoptera: Braconidae) and Aphidoletes aphidimyza (Diptera: Cecidomyiidae) to sweet alyssum and assessment of plant resources effects on their fitness. J. Econ. Entomol. 2018, 111, 533–541. [Google Scholar] [CrossRef]
- Jado, R.H.; Araj, S.E.; Irmaileh, B.A.; Shields, M.W. Floral resources to enhance the potential of the parasitoid Aphidius colemani for biological control of the aphid Myzus persicae. J. Appl. Entomol. 2019, 143, 34–42. [Google Scholar] [CrossRef]
- Aparicio, Y.; Gabarra, R.; Riudavets, J.; Arnó, J. Sustainable Plant Protection Programme, Institut de Recerca i Tecnologia Agroalimentàries (IRTA), Cabrils, Spain. Personal Observation. 2015–2016. [Google Scholar]
- Hassell, M.P.; Waage, J.K. Host-parasitoid population interactions. Ann. Rev. Entomol. 1984, 29, 89–114. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Aphid Species | Area | Sampling Method | Year | ||
|---|---|---|---|---|---|
| 2015 | 2016 | 2017 | |||
| Number of Samples Collected (Time Period) | |||||
| Myzus persicae | Lleida | Sentinel plants | 4 (18–24) | 6 (16–26) | — |
| Random samples | 5 (18–27) | 6 (17–23) | 2 (19–20) | ||
| Hyalopterus spp. | Lleida | Random samples | 5 (22–31) | 2 (23–28) | 1 (21) |
| Barcelona | Random samples | — | 3 (23–26) | 10 (16–27) | |
| Family and Subfamily | Species | Sentinel Plants | Random Sampling | |||
|---|---|---|---|---|---|---|
| 2015 | 2016 | 2015 | 2016 | 2017 | ||
| Parasitoids | ||||||
| Braconidae Aphidiinae | ||||||
| Aphidius matricariae (Haliday) | 344 | 213 | 27 | 26 | 180 | |
| Aphidius ervi Haliday | 12 | 8 | 0 | 0 | 9 | |
| Aphidius colemani Viereck | 1 | 16 | 0 | 0 | 0 | |
| Aphidius transcaspicus Telenga | 2 | 0 | 0 | 0 | 0 | |
| Lipolexis sp. | 4 | 11 | 0 | 0 | 1 | |
| Ephedrus persicae Froggat | 0 | 5 | 0 | 3 | 0 | |
| Ephedrus plagiator (Nees) | 1 | 0 | 0 | 0 | 0 | |
| Praon volucre (Haliday) | 1 | 5 | 0 | 0 | 0 | |
| Praon abjectum (Haliday) | 0 | 1 | 0 | 0 | 0 | |
| Diaeretiella rapae (McIntosh) | 1 | 0 | 0 | 0 | 0 | |
| Lysiphlebus testaceipes (Cresson) | 0 | 1 | 0 | 0 | 0 | |
| Hyperparasitoids | ||||||
| Figitidae Charipinae | ||||||
| Phaenoglyphis villosa (Hartig) | 6 | 18 | 1 | 23 | 0 | |
| Alloxysta pusilla (Kieffer) | 3 | 4 | 2 | 14 | 0 | |
| Alloxysta victrix (Westwood) | 0 | 5 | 0 | 3 | 0 | |
| Alloxysta arcuata (Kieffer) | 4 | 0 | 0 | 0 | 4 | |
| Alloxysta castanea (Hartig) | 0 | 0 | 0 | 1 | 0 | |
| Alloxysta fuscicornis (Hartig) | 0 | 0 | 0 | 0 | 1 | |
| Pteromalidae Asaphinae | ||||||
| Asaphes suspensus (Nees) | 0 | 0 | 0 | 38 | 1 | |
| Asaphes vulgaris (Walker) | 0 | 0 | 0 | 4 | 0 | |
| Pteromalinae | ||||||
| Pachyneuron aphidis (Bouché) | 1 | 0 | 0 | 13 | 10 | |
| Encyrtidae Encyrtinae | ||||||
| Syrphophagus nr. africanus | 16 | 0 | 0 | 9 | 0 | |
| Families and Subfamilies | Species | Lleida | Barcelona |
|---|---|---|---|
| Pteromalidae | |||
| Pteromalinae | Pachyneuron aphidis (Bouche) | 53 | 260 |
| Asaphinae | Asaphes suspensus (Nees) | 2 | 8 |
| Encyrtidae | |||
| Encyrtinae | Syrphophagus nr. africanus | 19 | 3 |
| Figitidae | |||
| Charipinae | Phaenoglyphis villosa (Hartig) | 1 | 2 |
| Alloxysta fuscicornis (Hartig) | 1 | 2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aparicio, Y.; Gabarra, R.; Riudavets, J.; Starý, P.; Tomanović, Ž.; Kocić, K.; Pujade Villar, J.; Ferrer Suay, M.; Cuesta Porta, V.; Arnó, J. Hymenoptera Complex Associated with Myzus persicae and Hyalopterus spp. in Peach Orchards in Northeastern Spain and Prospects for Biological Control of Aphids. Insects 2019, 10, 109. https://doi.org/10.3390/insects10040109
Aparicio Y, Gabarra R, Riudavets J, Starý P, Tomanović Ž, Kocić K, Pujade Villar J, Ferrer Suay M, Cuesta Porta V, Arnó J. Hymenoptera Complex Associated with Myzus persicae and Hyalopterus spp. in Peach Orchards in Northeastern Spain and Prospects for Biological Control of Aphids. Insects. 2019; 10(4):109. https://doi.org/10.3390/insects10040109
Chicago/Turabian StyleAparicio, Yahana, Rosa Gabarra, Jordi Riudavets, Petr Starý, Željko Tomanović, Korana Kocić, Juli Pujade Villar, Mar Ferrer Suay, Victor Cuesta Porta, and Judit Arnó. 2019. "Hymenoptera Complex Associated with Myzus persicae and Hyalopterus spp. in Peach Orchards in Northeastern Spain and Prospects for Biological Control of Aphids" Insects 10, no. 4: 109. https://doi.org/10.3390/insects10040109
APA StyleAparicio, Y., Gabarra, R., Riudavets, J., Starý, P., Tomanović, Ž., Kocić, K., Pujade Villar, J., Ferrer Suay, M., Cuesta Porta, V., & Arnó, J. (2019). Hymenoptera Complex Associated with Myzus persicae and Hyalopterus spp. in Peach Orchards in Northeastern Spain and Prospects for Biological Control of Aphids. Insects, 10(4), 109. https://doi.org/10.3390/insects10040109