
Role of microRNAs in Lung Carcinogenesis Induced by Asbestos

 , and
, and
Abstract
1. Introduction
2. Asbestos as a Carcinogenic Factor of the Environment
2.1. Asbestos and Lung Diseases
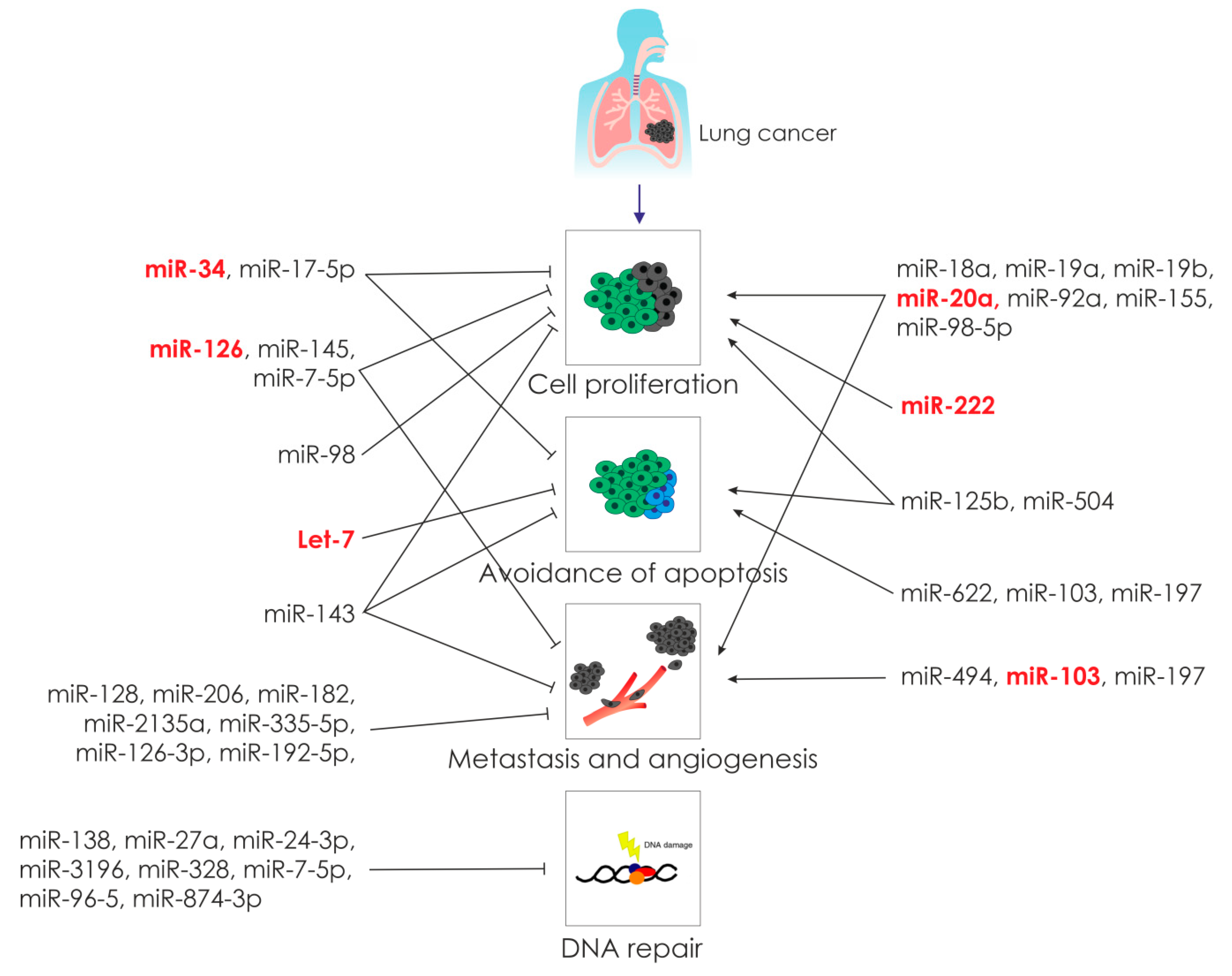
2.2. The Role of microRNAs in Lung Cancer Carcinogenesis
2.2.1. MicroRNA and Cell Proliferation in Lung Cancer
2.2.2. MicroRNA and Apoptosis in Lung Cancer
2.2.3. MicroRNA and Angiogenesis in Lung Cancer
2.2.4. MicroRNA and Metastasis in Lung Cancer
2.2.5. MicroRNAs Participating in Epithelial–Mesenchymal Transition (EMT)
2.2.6. MicroRNAs and DNA Repair in Lung Cancer

{kind=link}
{kind=link}
| MicroRNA | MicroRNA Targets and Mechanism | Role in Carcinogenesis | Ref. | |
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 |
| 1 | cluster of microRNA miR-17-92 (miR-18a, miR-19a, miR-19b, miR-20a and miR-92a) | miR-19a, miR-19b-1 hsa-miR-20a, and miR-92a inhibit translation of the messenger RNA encoding the tumor suppressor PTEN, enhancing cell proliferation and survival. miR-20a targets RNA encoding the E2F2/E2F3 transcription factors, which play a leading role in the regulation of the cell cycle. Repression of the TGF-β antiproliferative signaling pathway: miR-17 and miR-20a target TGF-β-receptor II (TGFBRII), miR-18a targets the participants of this signaling pathway Smad2 and Smad4; miR-18a and miR-19 directly inhibit the antiangiogenic factor thrombospondin-1 (TSP-1). | Proliferation, cell survival, angiogenesis | [35] |
| 2 | miR-155 | Inhibits translation of the messenger RNA encoding SHIP1 (negative regulator of proliferation) to promote cell growth; C/EBPβ (transcriptional activator for mir-143, which targets one of the main glycolysis enzymes—hk2); TP53INP1 (tumor suppressor regulating autophagy and apoptosis) leads to the inhibition of cell death; MSH2 and MSH6 (key misfit repair proteins) lead to decreased repair; FOXO3 (a transcription factor that regulates genes whose products are involved in apoptosis—for example, Bim and PUMA) leads to avoidance of apoptosis, SOCS1 (negative regulator of cytokine signal transduction) leads to increased proliferation; increases TNF-α levels by binding to the 3′ UTR region of mRNA and increasing transcript stability; with the participation of histone deacetylase HDAC2, represses BRCA1 transcription, which leads to a decrease in repair | Inhibition of apoptosis, proliferation metastases, Warburg effect | [42,43,44] |
| 3 | miR-125b | Inhibits translation of the messenger RNAs encoding p53 and proapoptotic proteins Bak1 and Puma; inhibits translation of the messenger RNA encoding the oncosuppressor p14ARF | Avoidance of apoptosis, proliferation | [50] |
| 4 | miR-504 | Inhibits translation of the messenger RNA encoding p53 | Avoidance of apoptosis, proliferation | [49] |
| 5 | miR-622 | Inhibits translation of the messenger RNA encoding Rb protein | Avoidance of apoptosis | [52] |
| 6 | miR -103 | Inhibits translation of the messenger RNA encoding proapoptotic protein Bax | Avoidance of apoptosis | [54] |
| 7 | miR-197 | Inhibits translation of the messenger RNA encoding NOXA and BMF | Avoidance of apoptosis | [55,56] |
| 8 | miR-494 | Inhibits translation of the messenger RNA encoding PTEN and subsequent activation of the Akt/eNOS pathway | Angiogenic effect | [68] |
| 9 | miR-222 | Inhibits translation of the messenger RNA encoding cyclin-dependent kinase inhibitor p27Kip1 | Proliferation | [46,47] |
| 10 | miR-10b | Inhibits translation of the messenger RNA encoding the homeobox D10, which increases expression prometastatic gene RHOC | Lung metastases | [83] |
| 11 | miR-652-3p | Inhibits translation of the messenger RNA encoding the Lgl1 protein, which promotes cell adhesion and inhibits cell migration by suppressing the expression of MMP2 and MMP14 and the re-expression of E-cadherin | Metastases | [102] |
| 12 | miR-98-5p | Inhibits translation of the messenger RNA encoding TGFBR1 | Proliferation, migration and invasion of A549 and H1299 cell lines | [64] |
| 13 | miR-27a | Inhibits translation of the messenger RNA encoding ATM | Cell survival and cell cycle progression | [113] |
| 14 | miR-346 | Inhibits translation of the messenger RNA encoding XPC | Promotion tumor growth | [110] |
| MicroRNA | MicroRNA Targets and Mechanism | Role in Carcinogenesis | Ref. | |
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 |
| 1 | Let-7 | Inhibits translation of the messenger RNAs encoding oncogenes, such as KRAS, NRAS, MYC, HMGA2, and MCT | Inhibits proliferation, inhibits PI3K-mTOR signaling pathway | [37] |
| 2 | miR-34 | Inhibits translation of the messenger RNA encoding the N-MUS oncogene; cyclin-dependent kinases CDK4 and CDK6; transmembrane receptor protein NOTCH1, involved in the signaling pathway of cancer stem cells; ubiquitin ligase MDMX, involved in p53 degradation; antiapoptotic protein BCL2; sirtuin 1 (SIRT1 gene) involved in p53 degradation; transcription factor E2F3; transcription factors involved in self-renewal of undifferentiated embryonic stem cells: NANOG and SOX2; an integral cellular glycoprotein that plays an important role in cell–cell interactions, cell adhesion, and CD44 migration | Inhibits proliferation, promotes cell cycle arrest and apoptosis | [48] |
| 3 | miR-126 | Inhibits translation of the messenger RNA encoding S1PR2, thereby inhibiting the PI3K/Akt signaling pathway; inhibits tumor angiogenesis by targeting VEGF-A | Inhibits proliferation and angiogenesis | [38,39] |
| 4 | miR-17-5p | Inhibits translation of the messenger RNA encoding TGFßR2, which is significantly increased in NSCLC tissues and cell lines | Inhibits proliferation, causes apoptosis of H460 NSCLC cells | [59] |
| 5 | miR-98 | Inhibits translation of the messenger RNA encoding PAK1, which promotes NSCLC cell proliferation, migration, and invasion | Inhibits proliferation | [66] |
| 6 | miR-128 | Inhibits translation of the messenger RNA encoding VEGF-A, vascular endothelial growth factor receptor 2, and VEGFR-3 | Inhibits angiogenesis | [69] |
| 7 | miR-206 | Inhibits the 14-3-3ζ/STAT3/HIF-1α/VEGF pathway | Inhibits angiogenesis | [70] |
| 8 | miR-135a | Decreased angiogenesis-related factors VEGF, bFGF, and IL-8 | Inhibits angiogenesis | [71] |
| 9 | miR-145 | Inhibits translation of the messenger RNA encoding mTOR/p70S6K1 | Inhibits proliferation | [41,42] |
| 10 | miR-646 | Inhibits translation of the messenger RNA encoding FGF2 and CCND2 | Inhibits proliferation, invasion, and suppress EMT of NSCLC cells in mice | [87] |
| 11 | miR-182 | Suppresses AKT phosphorylation and accumulation of the Snail transcription factor, which initiates EMT in lung cancer cells. Inhibits translation of the messenger RNA encoding the Met | Promotes the expression of E-cadherin, which leads to inhibition of EMT | [88] |
| 12 | miR-7-5p | Inhibits translation of the messenger RNA encoding NOVA2, which disrupts the angiogenesis | Inhibits proliferation, migration, and invasion of tumor | [89] |
| 13 | miR-206 | Inhibits translation of the messenger RNA encoding the actin-binding protein coronin 1C (CORO1C) | Reduces the ability of cells to grow and metastasize | [91] |
| 14 | miR-335-5p | Inhibits translation of the messenger RNA encoding ROCK1 | Leads to inhibition of TGF-β1-mediated EMT | [94] |
| 15 | miR-126-3p | Inhibits translation of the messenger RNA encoding chemokine receptor 1 (CCR1) | Inhibits NSCLC cell growth and metastasis | [96] |
| 16 | miR-192-5p | Inhibits translation of the messenger RNA encoding TRIM44 | Reduces migration and invasion | [97] |
| 17 | miR-143 | Inhibits translation of the messenger RNA encoding Limk1 | Suppresses NSCLC cell proliferation, induced apoptosis, and suppresses migration and invasion in vitro | [103] |
| 18 | miR-138 | Inhibits translation of the messenger RNA encoding H2AX | Inhibition on cell-cycle progression and cell grow | [111] |
| 19 | hsa-miR-526b | Inhibits translation of the messenger RNA encoding Ku80 | Suppresses NSCLC growth | [112] |
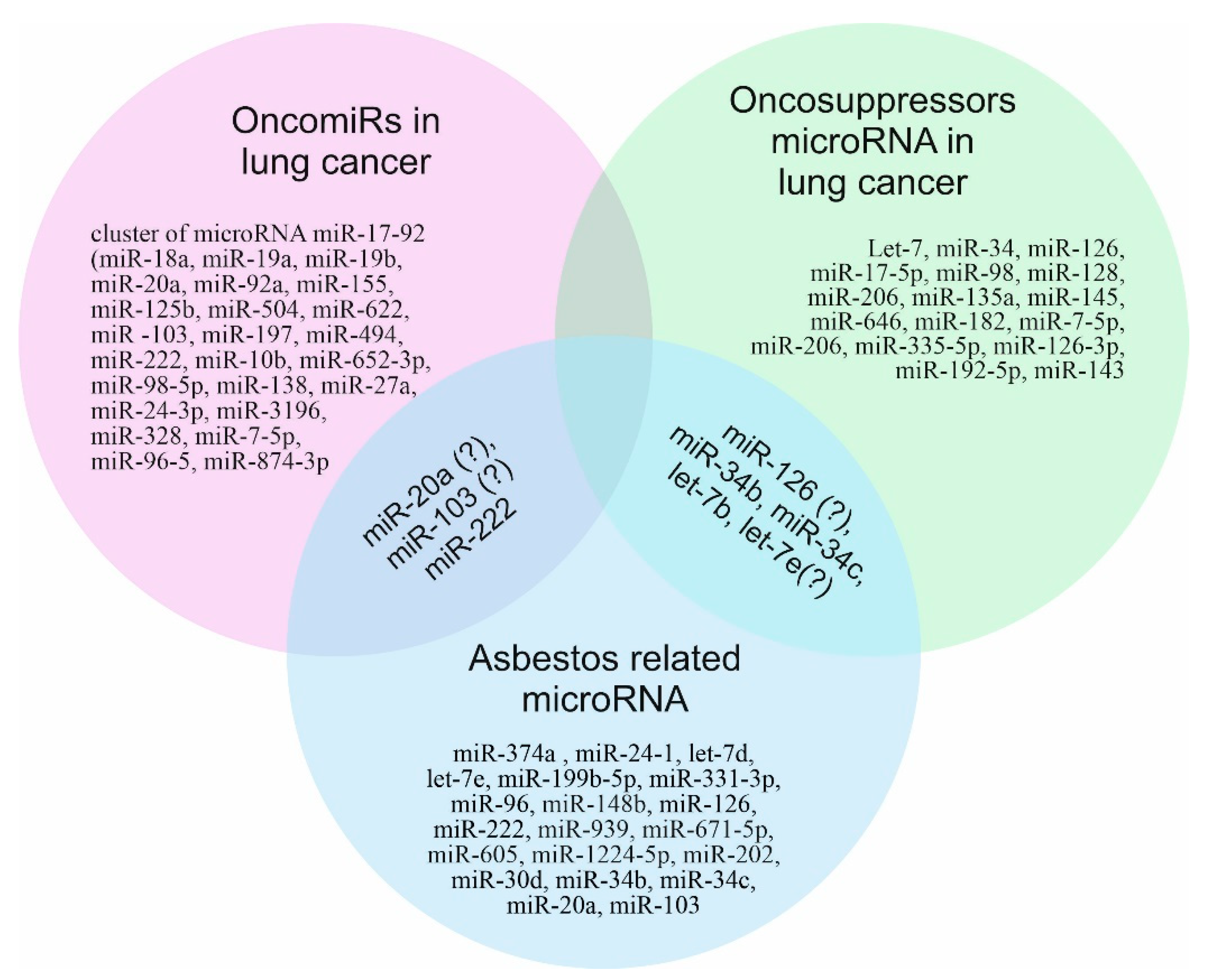
3. MicroRNAs as Biomarkers of Environmental Factors
4. Asbestos and MicroRNA
| MicroRNA | Expression Level | Type of Cancer | Sample | Ref. | |
|---|---|---|---|---|---|
| 1 | miR-374a | Overexpression | Lung cancer | Lung tissue | [130] |
| 2 | miR-24-1 | Overexpression | Lung cancer | Lung tissue | [130] |
| 3 | let-7d | Overexpression | Lung cancer | Lung tissue | [130] |
| 4 | let-7e | Overexpression | Lung cancer | Lung tissue | [130] |
| 5 | miR-199b-5p | Overexpression | Lung cancer | Lung tissue | [130] |
| 6 | miR-331-3p | Overexpression | Lung cancer | Lung tissue | [130] |
| 7 | miR-96 | Overexpression | Lung cancer | Lung tissue | [130] |
| 8 | miR-148b | Overexpression | Lung cancer | Lung tissue | [130] |
| 9 | miR-126 | Overexpression | Lung cancer | serum | [132] |
| 10 | miR-222 | Overexpression | Lung cancer | serum | [132] |
| 11 | miR-939 | down regulation | Lung cancer | Lung tissue | [130] |
| 12 | miR-671-5p | down regulation | Lung cancer | Lung tissue | [130] |
| 13 | miR-605 | down regulation | Lung cancer | Lung tissue | [130] |
| 14 | miR-1224-5p | down regulation | Lung cancer | Lung tissue | [130] |
| 15 | miR-202 | down regulation | Lung cancer | Lung tissue | [130] |
| 16 | miR-30d | down regulation | Lung cancer | Lung tissue | [140] |
| 17 | miR-34b | down regulation | mesothelioma | serum | [147] |
| 18 | miR-34c | down regulation | mesothelioma | serum | [147] |
| 19 | miR-20a | down regulation | mesothelioma | blood | [149] |
| 20 | miR-103 | down regulation | mesothelioma | blood | [149] |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Barta, J.A.; Powell, C.A.; Wisnivesky, J.P. Global epidemiology of lung cancer. Ann. Glob. Health 2019, 85, 1–16. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed]
- Didkowska, J.; Wojciechowska, U.; Manczuk, M.; Lobaszewski, J. Lung cancer epidemiology: Contemporary and future challenges worldwide. Ann. Transl. Med. 2016, 4, 150. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.D.; Nogueira, L.; Mariotto, A.B.; Rowland, J.H.; Yabroff, K.R.; Alfano, C.M.; Jemal, A.; Kramer, J.L.; Siegel, R.L. Cancer treatment and survivorship statistics, 2019. CA Cancer J. Clin. 2019, 69, 363–385. [Google Scholar] [CrossRef] [PubMed]
- Rahal, Z.E.; Nemr, S.; Sinjab, A.; Chami, H.; Tfayli, A.; Kadara, H. Smoking and lung cancer: AGeo-Regional Perspective. Front. Oncol 2017, 7, 194. [Google Scholar] [CrossRef]
- Dicker, D.; Nguyen, G.; Abate, D. GBD 2017 Mortality Collaborators. Global, regional, and national age-sex-specific mortality and life expectancy, 1950-2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1684–1735. [Google Scholar] [CrossRef]
- Pallis, A.G.; Syrigos, K.N. Lung cancer in never smokers: Disease characteristics and risk factors. Crit. Rev. Oncol. Hematol. 2013, 88, 494–503. [Google Scholar] [CrossRef]
- Martinez, V.D.; Sage, A.P.; Marshall, E.A.; Suzuki, M.; Goodarzi, A.A.; Dellaire, G.; Lam, W.L. Oncogenetics of Lung Cancer induced by environmental carcinogens. Oncog. Carcinog. 2019. [Google Scholar] [CrossRef]
- Testa, U.; Castelli, G.; Pelosi, E. Lung Cancers: Molecular Characterization, Clonal Heterogeneity and Evolution, and Cancer Stem Cells. Cancers 2018, 10, 248. [Google Scholar] [CrossRef]
- IARC. Asbestos (chrystolyte, amosite, crocidolite, trmolite, actioolite and antropolite). IARC Monogr. Eval. Carcinog. Risks Hum. 2012, 100c, 219–309. Available online: monographs.iarc.fr?ENG/Monographs/vol100c/index.php (accessed on 30 January 2021).
- Asbestos Safety and Eradication Agency. About Asbestos. 2019. Available online: https://www.asbestossafety.gov.au/about-asbestos/about-asbestos (accessed on 11 November 2019).
- Takahashi, K.; Landrigan, P.J. The Global health dimensions of asbestos and asbestos-related diseases. Ann. Glob. Health 2016, 82, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Norbet, C.; Joseph, A.; Rossi, S.S. Asbestos-related lung disease: A pictorial review. Curr. Probl. Diagn. Radiol. 2015, 44, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.X.; Jaurand, M.C.; Kamp, D.W.; Whysner, J.; Hei, T.K. Role of mutagenicity in asbestos fiber induced carcinogenicity and other diseases. J. Toxicol. Environ. Health Part B Crit. Rev. 2011, 14, 179–245. [Google Scholar] [CrossRef] [PubMed]
- The Mesothelioma Center. Asbestos-Related Lung Cancer. 2019. Available online: https://www.asbestos.com/cancer/lung-cancer/ (accessed on 11 November 2019).
- Kameda, T.; Takahashi, K.; Kim, R.; Jiang, Y.; Movahed, M.; Park, E.-K.; Rantanen, J. Asbestos: Use, bans and disease burden in Europe. Bull. World Health Organ. 2014, 92, 790–797. [Google Scholar] [CrossRef]
- Ospina, D.; Villegas, V.E.; Rodriguez-Leguizamon, G.; Rondon-Lagos, M. Analyzing biological and molecular charachteristics and genomic damage induced by exposure to asbestos. Cancer Manag. Res. 2019, 11, 4997–5012. [Google Scholar] [CrossRef]
- Wolff, H.; Vehmas, T.; Oksa, P.; Rantanen, J.; Vainio, H. Asbestos, asbestosis, and cancer, the Helsinki criteria for diagnosis and attribution 2014: Recommendations. Scand. J. Work Environ. Health 2015, 41, 5–15. [Google Scholar] [CrossRef]
- The International Ban Asbestos Secretariat. Current Asbestos Bans. 2018. Available online: http://www.ibasecretariat.org/alpha_ban_list.php (accessed on 1 April 2018).
- Liu, G.; Cheresh, P.; Kamp, D.W. Molecular basis of asbestos-induced lung disease. Annu. Rev. Pathol. 2013, 24, 161–187. [Google Scholar] [CrossRef]
- Mossman, B.T.; Lippmann, M.; Hesterberg, T.W.; Kelsey, K.T.; Barchowsky, A.; Bonner, J.C. Pulmonary endpoints (lung carcinomas and asbestosis) following inhalation exposure to asbestos. J. Toxicol. Environ. Health B Crit. Rev. 2011, 14, 76–121. [Google Scholar] [CrossRef]
- Markowitz, S. Asbestos-related lung cancer and malignant mesothelioma of the pleura: Selected current issues. Semin. Respir. Crit. Care Med. 2015, 36, 334–346. [Google Scholar] [CrossRef]
- Baldys, A.; Aust, A.E. Role of iron in inactivation of epidermal growth factor receptor after asbestos treatment of human lung and pleural target cells. Am. J. Respir. Cell Mol. Biol. 2005, 32, 436–442. [Google Scholar] [CrossRef]
- Aljandali, A.; Pollack, H.; Yeldandi, A. Asbestos causes apoptosis in alveolar epithelial cells: Role of iron-induced free radicals. J. Lab. Clin. Med. 2001, 137, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, D.; Kamp, D.W. Asbestos-induced pulmonary toxicity: Role of DNA damage and apoptosis. Exp. Biol. Med. 2003, 228, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Arsenic, M. Fibers and Dust International Agency for Research on Cancer (IARC) Monographs; International Agency for Research on Cancer: Lyon, France, 2012. [Google Scholar]
- Murthy, S.; Ryan, A.; He, C.; Mallampalli, R.K. Carter AB. Rac1-mediated mitochondrial H2O2 generation regulates MMP-9 gene expression in macrophages via inhibition of SP-1 and AP-1. J. Biol. Chem. 2010, 285, 25062–25073. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Murthy, S.; McCormick, M.L.; Spitz, D.R.; Ryan, A.J.; Carter, A.B. Mitochondrial Cu, Zn-superoxide dismutase mediates pulmonary fibrosis by augmenting H2O2 generation. J. Biol. Chem. 2011, 286, 15597–15607. [Google Scholar] [CrossRef]
- Bulgakova, O.; Kussainova, A.; Kausbekova, A.; Bersimbaev, R. The free-circulating mtDNA copies number in plasma of patients with NSCLC. Ann. Oncol. 2019, 30, 467. [Google Scholar] [CrossRef]
- Wu, K.L.; Tsai, Y.M.; Lien, C.T.; Kuo, P.L.; Hung, J.Y. The Roles of MicroRNA in Lung Cancer. Int. J. Mol. Sci. 2019, 20, 1611. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Chen, C.; Chu, X. New insights into the regulatory role of microRNA in tumor angiogenesis and clinical implications. Mol. Cancer 2018, 17, 22. [Google Scholar] [CrossRef]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Izzotti, A.; Carozzo, S.; Pulliero, A.; Zhabayeva, D.; Ravetti, J.L.; Bersimbaev, R. Extracellular MicroRNA in liquid biopsy: Applicability in cancer diagnosis and prevention. Am. J. Cancer Res. 2016, 6, 1461–1493. [Google Scholar]
- Bersimbaev, R.; Pulliero, A.; Bulgakova, O.; Kussainova, A.A.; Aripova, A.; Izzotti, A. Radon Biomonitoring and microRNA in Lung Cancer. Int. J. Mol. Sci. 2020, 21, 2154. [Google Scholar] [CrossRef]
- Hwang, H.W.; Mendell, J.T. MicroRNAs in cell proliferation, cell death, and tumorigenesis. Br. J. Cancer 2006, 94, 776–780. [Google Scholar] [CrossRef] [PubMed]
- García-Martínez, A.; López-Muñoz, B.; Fajardo, C.; Cámara, R.; Lamas, C.; Silva-Ortega, S.; Aranda, I.; Picó, A. Increased E2F1 mRNA and miR-17-5p Expression Is Correlated to Invasiveness and Proliferation of Pituitary Neuroendocrine Tumours. Diagnostics 2020, 10, 227. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, D.; Gu, C.; Liu, X.; Pei, W.; Li, J.; Cao, Y.; Jiao, Y.; Tong, J.; Nie, J. Down-regulation of let-7 microRNA increased K-ras expression in lung damage induced by radon. Environ. Toxicol. Pharmacol. 2015, 40, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Li, D.; Gu, Y.; Wen, Z.M.; Jie, J.; Zhao, D.; Peng, L.P. MicroRNA-126 Targeting PIK3R2 Inhibits NSCLC A549 Cell Proliferation, Migration, and Invasion by Regulation of PTEN/PI3K/AKT Pathway. Clin. Lung Cancer 2016, 17, e65–e75. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Chen, S.; Zhao, J.; Zhou, Y.; Xu, L. MicroRNA-126: A new and promising player in lung cancer (Review). Oncol. Lett. 2020, 21, 1. [Google Scholar] [CrossRef]
- Tsay, J.J.; Tchou-Wong, K.M.; Yie, T.; Leibert, E.; Segal, L.N.; Greenberg, A.; Pass, H.; Rom, W.N. Bronchial brushings’ microRNA and field cancerization in lung adenocarcinoma. Am. J. Respir. Crit. Care Med. 2013, 187, A4753. [Google Scholar]
- Ye, D.; Shen, Z.; Zhou, S. Function of microRNA-145 and mechanisms underlying its role in malignant tumor diagnosis and treatment. Cancer Manag. Res. 2019, 11, 969–979. [Google Scholar] [CrossRef]
- Xu, Q.; Liu, L.Z.; Qian, X.; Chen, Q.; Jiang, Y.; Li, D.; Lai, L.; Jiang, B.H. MiR-145 directly targets p70S6K1 in cancer cells to inhibit tumor growth and angiogenesis. Nucleic Acids Res. 2012, 40, 761–774. [Google Scholar] [CrossRef]
- Shao, C.; Yang, F.; Qin, Z.; Jing, X.; Shu, Y.; Shen, H. The value of miR-155 as a biomarker for the diagnosis and prognosis of lung cancer: A systematic review with meta-analysis. BMC Cancer 2019, 19, 1103. [Google Scholar] [CrossRef]
- Xiang, X.; Zhuang, X.; Ju, S.; Zhang, S.; Jiang, H.; Mu, J.; Zhang, L.; Miller, D.; Grizzle, W.; Zhang, H.G. miR-155 promotes macroscopic tumor formation yet inhibits tumor dissemination from mammary fat pads to the lung by preventing EMT. Oncogene 2011, 30, 3440–3453. [Google Scholar] [CrossRef]
- Kaipeng, X.H.; Cheng, L.; Cheng, W.; Na, Q.; Wei, S.; Yayun, G.; Caiwang, Y.; Kai, Z.; Ningbin, D.; Meng, Z.; et al. A functional variant in miR-155 regulation region contributes to lung cancer risk and survival. Oncotarget 2015, 6, 42781–42792. [Google Scholar]
- Zaporozhchenko, I.A.; Morozkin, E.S.; Ponomaryova, A.A.; Rykova, E.Y.; Cherdyntseva, N.V.; Zheravin, A.A.; Pashkovskaya, O.A.; Pokushalov, E.A.; Vlassov, V.V.; Laktionov, P.P. Profiling of 179 microRNA Expression in Blood Plasma of Lung Cancer Patients and Cancer-Free Individuals. Sci. Rep. 2018, 8, 6348. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Ding, S.; Xu, Y.; Huang, H. MicroRNA-222 promotes human non-small cell lung cancer H460 growth by targeting p27. Int. J. Clin. Exp. Med. 2015, 8, 5534–5540. [Google Scholar] [PubMed]
- Navarro, F.; Lieberman, J. miR-34 and p53: New Insights into a Complex Functional Relationship. PLoS ONE 2015, 10, e0132767. [Google Scholar] [CrossRef] [PubMed]
- Bublik, D.R.; Bursać, S.; Sheffer, M.; Oršolić, I.; Shalit, T.; Tarcic, O.; Kotler, E.; Mouhadeb, O.; Hoffman, Y.; Fuchs, G.; et al. Regulatory module involving FGF13, miR-504, and p53 regulates ribosomal biogenesis and supports cancer cell survival. Proc. Natl. Acad. Sci. USA 2017, 114, E496–E505. [Google Scholar] [CrossRef] [PubMed]
- Banzhaf-Strathmann, J.; Edbauer, D. Good guy or bad guy: The opposing roles of microRNA 125b in cancer. CCS 2014, 12, 30. [Google Scholar] [CrossRef]
- Wang, Y.; Scheiber, M.N.; Neumann, C.; Calin, G.A.; Zhou, D. MicroRNA regulation of ionizing radiation-induced premature senescence. Int. J. Radiat. Oncol. Biol. Phys. 2011, 81, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Yu, J.; Qi, X.; Liang, L.; Zhang, Y.; Ding, Y.; Lin, X.; Li, G.; Ding, Y. Radiation-induced microRNA-622 causes radioresistance in colorectal cancer cells by down-regulating Rb. Oncotarget 2015, 6, 15984–15994. [Google Scholar] [CrossRef]
- Zevine, A.J.; Hu, W.; Feng, Z. The P53 pathway: What questions remain to be explored? Cell Death Differ. 2006, 13, 1027–1036. [Google Scholar]
- Zhou, J.; Zhang, H.; Chen, L.; Song, Q. Effect of miR 103 on proliferation and apoptosis of non small cell lung cancer by inhibition of FBW7 expression. Oncol. Lett. 2020. [Google Scholar] [CrossRef]
- Fiori, M.; Barbini, C.; Haas, T.L.; Marroncelli, N.; Patrizii, M.; Biffoni, M.; De Maria, R. Antitumor effect of miR-197 targeting in p53 wild-type lung cancer. Cell Death Differ. 2014, 21, 774–782. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, C. miR-197-3p induced downregulation of lysine 63 deubiquitinase promotes cell proliferation and inhibits cell apoptosis in lung adenocarcinoma cell lines. Mol. Med. Rep. 2018, 17, 3921–3927. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhou, H.; Luo, J.; Huang, J. MicroRNA-17-5p inhibits proliferation and triggers apoptosis in non-small cell lung cancer by targeting transforming growth factor β receptor 2. Exp. Med. 2017, 13, 2715–2722. [Google Scholar] [CrossRef]
- Chi, E.Y.; Viriyapak, B.; Kwack, H.S.; Lee, Y.K.; Kim, S.I.; Lee, K.H.; Park, T.C. Regulation of paclitaxel-induced programmed cell death by autophagic induction: A model for cervical cancer. Obstet. Gynecol. Sci. 2013, 56, 84. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chatterjee, A.; Chattopadhyay, D.; Chakrabarti, G. miR-17-5p Downregulation Contributes to Paclitaxel Resistance of Lung Cancer Cells through Altering Beclin1 Expression. PLoS ONE 2014, 9, e95716. [Google Scholar] [CrossRef]
- Li, J.; Zhou, Q.; Liang, Y.; Pan, W.; Bei, Y.; Zhang, Y.; Wang, J.; Jiao, Z. miR-486 inhibits PM2.5-induced apoptosis and oxidative stress in human lung alveolar epithelial A549 cells. Ann. Transl. Med. 2018, 6, 209. [Google Scholar] [CrossRef]
- Ahn, M.J.; Kang, K.A.; Ryu, Y.S.; Hyun, Y.J.; Shilnikova, K.; Zhen, A.X.; Jeong, J.W.; Choi, Y.H.; Kang, H.K.; Koh, Y.S.; et al. Particulate matter 2.5 damages skin cells by inducing oxidative stress, subcellular organelle dysfunction, and apoptosis. Arch. Toxicol. 2018, 92, 2077–2091. [Google Scholar]
- Yang, N.; Liang, Y.; Yang, P.; Yang, T.; Jiang, L. Propofol inhibits lung cancer cell viability and induces cell apoptosis by upregulating microRNA-486 expression. Braz. J. Med. Biol. Res. 2017, 50. [Google Scholar] [CrossRef]
- Yu, S.; Geng, S.; Hu, Y. miR-486-5p inhibits cell proliferation and invasion through repressing GAB2 in non-small cell lung cancer. Oncol. Lett. 2018, 16, 3525–3530. [Google Scholar] [CrossRef]
- Zhou, H.; Huang, Z.; Chen, X.; Chen, S. miR-98 inhibits expression of TWIST to prevent progression of non-small cell lung cancers. Biomed. Pharmacother. 2017, 89, 1453–1461. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Liu, X.; Chen, Q.; Liu, T.; Lu, C.; Yu, J.; Miao, Y.; Wei, J. Downregulated miR-98-5p promotes PDAC proliferation and metastasis by reversely regulating MAP4K4. J. Exp. Clin. Cancer Res. 2018, 37, 130. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Zhang, X.; Shi, J. MiR-98 inhibits cell proliferation and invasion of non-small cell carcinoma lung cancer by targeting PAK1. Int. J. Clin. Exp. Med. 2015, 8, 20135–20145. [Google Scholar] [PubMed]
- Lou, W.; Liu, J.; Gao, Y.; Zhong, G.; Chen, D.; Shen, J.; Bao, C.; Xu, L.; Pan, J.; Cheng, J.; et al. MicroRNAs in cancer metastasis and angiogenesis. Oncotarget 2017, 8, 115787–115802. [Google Scholar] [CrossRef] [PubMed]
- Mao, G.; Liu, Y.; Fang, X.; Liu, Y.; Fang, L.; Lin, L.; Liu, X.; Wang, N. Tumor-derived microRNA-494 promotes angiogenesis in non-small cell lung cancer. Angiogenesis 2015, 18, 373–382. [Google Scholar] [CrossRef]
- Hu, J.; Cheng, Y.; Li, Y.; Jin, Z.; Pan, Y.; Liu, G.; Fu, S.; Zhang, Y.; Feng, K.; Feng, Y. microRNA-128 plays a critical role in human non-small cell lung cancer tumourigenesis, angiogenesis and lymphangiogenesis by directly targeting vascular endothelial growth factor-C. Eur. J. Cancer 2014, 50, 2336–2350. [Google Scholar] [CrossRef]
- Xue, D.; Yang, Y.; Liu, Y.; Wang, P.; Dai, Y.; Liu, Q.; Chen, L.; Shen, J.; Ju, H.; Li, Y.; et al. MicroRNA-206 attenuates the growth and angiogenesis in non-small cell lung cancer cells by blocking the 14-3-3ζ/STAT3/HIF-1α/VEGF signaling. Oncotarget 2016, 7, 79805–79813. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, S.; Li, J.; Wang, D.; Li, Q. Effect of microRNA-135a on Cell Proliferation, Migration, Invasion, Apoptosis and Tumor Angiogenesis Through the IGF-1/PI3K/Akt Signaling Pathway in Non-Small Cell Lung Cancer. Cell Physiol. Biochem. 2017, 42, 1431–1446. [Google Scholar] [CrossRef]
- Chaffer, C.L.; Weinberg, R.A. A perspective on cancer cell metastasis. Science 2011, 331, 1559–1564. [Google Scholar] [CrossRef]
- Wu, S.G.; Chang, T.H.; Liu, Y.N.; Shih, J.Y. MicroRNA in Lung Cancer Metastasis. Cancers 2019, 11, 265. [Google Scholar] [CrossRef]
- Shin, D.Y.; Na, I.I.; Kim, C.H.; Park, S.; Baek, H.; Yang, S.H. EGFR mutation and brain metastasis in pulmonary adenocarcinomas. J. Thorac. Oncol. 2014, 9, 195–199. [Google Scholar] [CrossRef]
- Cho, Y.J.; Cho, Y.M.; Kim, S.H.; Shin, K.H.; Jung, S.T.; Kim, H.S. Clinical analysis of patients with skeletal metastasis of lung cancer. BMC Cancer 2019, 19, 303. [Google Scholar] [CrossRef] [PubMed]
- Tamura, T.; Kurishima, K.; Nakazawa, K.; Kagohashi, K.; Ishikawa, H.; Satoh, H.; Hizawa, N. Specific organ metastases and survival in metastatic non-small-cell lung cancer. Mol. Clin. Oncol. 2015, 3, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Edme, N.; Downward, J.; Thiery, J.P.; Boyer, B. Ras induces NBT-II epithelial cell scattering through the coordinate activities of Ras and MAPK pathways. J. Cell Sci. 2002, 115, 2591–2601. [Google Scholar] [PubMed]
- Jenndahl, L.E.; Isakson, P.; Baeckstrom, D. c-erbB2-induced epithelial-mesenchymal transition in mammary epithelial cells is suppressed by cell-cell contact and initiated prior to E-cadherin downregulation. Int. J. Oncol. 2005, 27, 439–448. [Google Scholar] [CrossRef]
- Wise, R.; Zolkiewska, A. Metalloprotease-dependent activation of EGFR modulates CD44(+)/CD24(-) populations in triple negative breast cancer cells through the MEK/ERK pathway. Breast Cancer Res. Treat. 2017, 166, 421–433. [Google Scholar] [CrossRef]
- Claperon, A.; Mergey, M.; Ho-Bouldoires, T.H.N.; Vignjevic, D.; Wendum, D.; Chretien, Y.; Merabtene, F.; Frazao, A.; Paradis, V.; Housset, C.; et al. EGF/EGFR axis contributes to the progression of cholangiocarcinoma through the induction of an epithelial-mesenchymal transition. J. Hepatol. 2014, 61, 325–332. [Google Scholar] [CrossRef]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.; Nieto, M.A. Epithelial-mesenchymal transitions in development and disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef]
- Ma, L.; Teruya-Feldstein, J.; Weinberg, R.A. Tumour invasion and metastasis initiated by microRNA-10b in breast cancer. Nature 2007, 449, 682–688. [Google Scholar] [CrossRef]
- Li, Y.; Li, Y.; Liu, J.; Fan, Y.; Li, X.; Dong, M.; Liu, H.; Chen, J. Expression levels of microRNA-145 and microRNA-10b are associated with metastasis in non-small cell lung cancer. Cancer Biol. 2016, 17, 272–279. [Google Scholar] [CrossRef]
- Li, W.; Liu, M.; Feng, Y.; Xu, Y.-F.; Huang, Y.-F.; Che, J.-P.; Wang, G.-C.; Yao, X.-D.; Zheng, J.-H. Downregulated miR-646 in clear cell renal carcinoma correlated with tumour metastasis by targeting the nin one binding protein (NOB1). Br. J. Cancer 2014, 111, 1188–1200. [Google Scholar] [CrossRef]
- Zhang, P.; Tang, W.M.; Zhang, H.; Li, Y.Q.; Peng, Y.; Wang, J.D.; Liu, G.N.; Huang, X.T.; Zhao, J.J.; Li, G.X.; et al. MiR-646 inhibited cell proliferation and EMT-induced metastasis by targeting FOXK1 in gastric cancer. Br. J. Cancer 2017, 117, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Jin, Y.; Deng, S.-C.; Deng, S.-J.; Zhu, S.; Liu, Y.; Li, X.; He, C.; Liu, M.-L.; Zeng, Z.; et al. MicroRNA-646-mediated reciprocal repression between HIF-1alpha and MIIP contributes to tumorigenesis of pancreatic cancer. Oncogene 2018, 37, 1743–1758. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wu, J.; Li, Y.; Jiang, Y.; Wang, L.; Chen, Y.; Lv, Y.; Zou, Y.; Ding, X. Circ_0000527 promotes the progression of retinoblastoma by regulating miR-646/LRP6 axis. Cancer Cell Int. 2020, 20, 1. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, H.; Li, Y.; Zhao, C.; Fan, Y.; Liu, J.; Li, X.; Liu, H.; Chen, J. MiR-182 inhibits the epithelial to mesenchymal transition and metastasis of lung cancer cells by targeting the Met gene. Mol. Carcinog. 2018, 57, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H. MiR-7-5p suppresses tumor metastasis of non-small cell lung cancer by targeting NOVA2. Cell. Mol. Biol. Lett. 2019, 24, 60. [Google Scholar] [CrossRef] [PubMed]
- Giampietro, C.; Deflorian, G.; Gallo, S.; Di Matteo, A.; Pradella, D.; Bonomi, S.; Belloni, E.; Nyqvist, D.; Quaranta, V.; Confalonieri, S.; et al. The alternative splicing factor Nova2 regulates vascular development and lumen formation. Nat. Commun. 2015, 6, 8479. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Peng, L. MiR-206 may suppress non-small lung cancer metastasis by targeting CORO1C. Cell. Mol. Biol. Lett. 2020, 25, 22. [Google Scholar] [CrossRef]
- Cheng, X.; Wang, X.; Wu, Z.; Tan, S.; Zhu, T.; Ding, K. CORO1C expression is associated with poor survival rates in gastric cancer and promotes metastasis in vitro. FEBS Open Bio 2019, 9, 1097–1108. [Google Scholar] [CrossRef]
- Lim, J.P.; Shyamasundar, S.; Gunaratne, J.; Scully, O.J.; Matsumoto, K.; Bay, B.H. YBX1 gene silencing inhibits migratory and invasive potential via CORO1C in breast cancer in vitro. BMC Cancer 2017, 17, 201. [Google Scholar] [CrossRef]
- Du, W.; Tang, H.; Lei, Z.; Zhu, J.; Zeng, Y.; Liu, Z.; Huang, J.-A. miR-335-5p inhibits TGF-β1-induced epithelial–mesenchymal transition in non-small cell lung cancer via ROCK1. Respir. Res. 2019, 20, 225. [Google Scholar] [CrossRef]
- Hu, C.; Zhou, H.; Liu, Y.; Huang, J.; Liu, W.; Zhang, Q.; Tang, Q.; Sheng, F.; Li, G.; Zhang, R. ROCK1 promotes migration and invasion of non-small-cell lung cancer cells through the PTEN/PI3K/FAK pathway. Int. J. Oncol. 2019, 55, 833–844. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Zhang, Y.-S.; Zhang, S.; Cheng, Z.-M.; Yu, J.-L.; Zhou, S.; Song, J. MiR-126-3p suppresses the growth, migration and invasion of NSCLC via targeting CCR1. Eur. Rev. Med. Pharm. Sci. 2019, 23, 679–689. [Google Scholar]
- Zou, P.; Zhu, M.; Lian, C.; Wang, J.; Chen, Z.; Zhang, X.; Yang, Y.; Chen, X.; Cui, X.; Liu, J.; et al. miR-192-5p suppresses the progression of lung cancer bone metastasis by targeting TRIM44. Sci. Rep. 2019, 9, 19619. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Meng, Q.; Chen, X.; Zhao, Y.; Liu, W.; Hu, J.; Xue, F.; Wang, X.; Cai, L. TRIM44 promotes proliferation and metastasis in non-small cell lung cancer via mTOR signaling pathway. Oncotarget 2016, 7, 30479–30491. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.Y.; Wang, L.; Zhu, M.X.; Deng, X.Y.; Wang, D.H.; Zhang, S.M.; Ying, J.H.; Yuan, X.; Wang, Q.; Xuan, T.F.; et al. TRIM44 activates the AKT/mTOR signal pathway to induce melanoma progression by stabilizing TLR4. J. Exp. Clin. Cancer Res. 2019, 38, 137. [Google Scholar] [CrossRef]
- Xiong, D.; Jin, C.; Ye, X.; Qiu, B.; Jianjun, X.; Zhu, S.; Xiang, L.; Wu, H.; Yongbing, W. TRIM44 promotes human esophageal cancer progression via the AKT/mTOR pathway. Cancer Sci. 2018, 109, 3080–3092. [Google Scholar] [CrossRef]
- Zhao, Y.; Zheng, R.; Ning, D.; Xie, F. MiR-625 Inhibits Tumor Cell Invasion, Migration and EMT by Negatively Regulating the Expression of Resistin in Non-Small Cell Lung. Cancer Manag. Res. 2020, 12, 4171–4180. [Google Scholar] [CrossRef]
- Yang, W.; Zhou, C.; Luo, M.; Shi, X.; Li, Y.; Sun, Z.; Zhou, F.; Chen, Z.; He, J. MiR-652-3p is upregulated in non-small cell lung cancer and promotes proliferation and metastasis by directly targeting Lgl1. Oncotarget 2016, 7, 16703–16715. [Google Scholar] [CrossRef]
- Xia, H.; Sun, S.; Wang, B.; Wang, T.; Liang, C.; Li, G.; Huang, C.; Qi, D.; Chu, X. miR-143 Inhibits NSCLC Cell Growth and Metastasis by Targeting Limk1. Int. J. Mol. Sci 2014, 15, 11973–11983. [Google Scholar] [CrossRef]
- Tapia, T.; Ottman, R.; Chakrabarti, R. LIM kinase1 modulates function of membrane type matrix metalloproteinase 1: Implication in invasion of prostate cancer cells. Mol. Cancer 2011, 10, 6. [Google Scholar] [CrossRef]
- Bagheri-Yarmand, R.; Mazumdar, A.; Sahin, A.A.; Kumar, R. LIM kinase 1 increases tumor metastasis of human breast cancer cells via regulation of the urokinase-type plasminogen activator system. Int. J. Cancer 2006, 118, 2703–2710. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Yu, Q.; Chu, Y.; Zhu, X.; Lu, W.; Liu, Q.; Wang, Q. MicroRNA-98-5p inhibits proliferation and metastasis in non-small cell lung cancer by targeting TGFBR1. Int. J. Oncol. 2019, 54, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Zha, Y.; Li, T.; Bai, J.; Tang, L.; Deng, J.; He, R.; Dong, F.; Zhang, Q. Oxidative effects of lungs in Wistar rats caused by long-term exposure to four kinds of China representative chrysotile. Environ. Sci. Pollut. Res. Int. 2019, 26, 18708–18718. [Google Scholar] [CrossRef] [PubMed]
- Mairinger, F.D.; Werner, R.; Flom, E.; Schmeller, J.; Borchert, S.; Wessolly, M.; Wohlschlaeger, J.; Hager, T.; Mairinger, T.; Kollmeier, J.; et al. miRNA regulation is important for DNA damage repair and recognition in malignant pleural mesothelioma. Virchows Arch. 2017, 470, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Nouws, J.; Wan, F.; Finnemore, E.; Roque, W.; Kim, S.J.; Bazan, I.S.; Li, C.X.; Sköld, C.M.; Dai, Q.; Yan, X.; et al. MicroRNA miR-24-3p reduces DNA damage responses, apoptosis, and susceptibility to chronic obstructive pulmonary disease. JCI Insight 2020, 8, 134218. [Google Scholar] [CrossRef]
- Szatkowska, M.; Krupa, R. Regulation of DNA Damage Response and Homologous Recombination Repair by microRNA in Human Cells Exposed to Ionizing Radiation. Cancers 2020, 12, 1838. [Google Scholar] [CrossRef]
- Yang, H.; Luo, J.; Liu, Z.; Zhou, R.; Luo, H. MicroRNA-138 Regulates DNA Damage Response in Small Cell Lung Cancer Cells by Directly Targeting H2AX. Cancer Investig. 2015, 33, 126–136. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Fu, S.L.; Xu, S.Q.; Zhou, X.; Liu, X.S.; Xu, Y.J.; Zhao, J.P.; Wei, S. By downregulating Ku80, HSA-miR-526b suppresses non-small cell lung cancer. Oncotarget 2015, 6, 1462–1477. [Google Scholar] [CrossRef]
- Di Francesco, A.; De Pittà, C.; Moret, F.; Barbieri, V.; Celotti, L.; Mognato, M. The DNA-damage response to γ-radiation is affected by miR-27a in A549 cells. Int. J. Mol. Sci. 2013, 14, 17881–17896. [Google Scholar] [CrossRef]
- Cortez, M.A.; Valdecanas, D.; Niknam, S.; Peltier, H.J.; Diao, L.; Giri, U.; Komaki, R.; Calin, G.A.; Gomez, D.R.; Chang, J.Y.; et al. In Vivo Delivery of miR-34a Sensitizes Lung Tumors to Radiation Through RAD51 Regulation. Mol. Ther. Nucleic Acids 2015, 4, e270. [Google Scholar] [CrossRef]
- Piotto, C.; Biscontin, A.; Millino, C.; Mognato, M. Functional validation of miRNAs targeting genes of DNA double-strand break repair to radiosensitize non-small lung cancer cells. Biochim. Biophys. Acta Gene Regul. Mech. 2018, 1861, 1102–1118. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.; Yang, H.; Zhu, Y.; Ruan, M.; Huang, Y.; Zhang, Q. MiR-7-5p-mediated downregulation of PARP1 impacts DNA homologous recombination repair and resistance to doxorubicin in small cell lung cancer. BMC Cancer 2019, 19, 602. [Google Scholar] [CrossRef]
- Yang, H.; Rivera, Z.; Jube, S.; Nasu, M.; Bertino, P.; Goparaju, C.; Franzoso, G.; Lotze, M.; Krausz, T.; Pass, H.I.; et al. Programmed necrosis induced by asbestos in human mesothelial cells causes high-mobility group box 1 protein release and resultant inflammation. Proc. Natl. Acad. Sci. USA 2010, 107, 12611–12616. [Google Scholar] [CrossRef] [PubMed]
- Valencia-Quintana, R.; Sánchez-Alarcón, J.; Tenorio-Arvide, M.G.; Deng, Y.; Montiel-González, J.M.; Gómez-Arroyo, S.; Villalobos-Pietrini, R.; Cortés-Eslava, J.; Flores-Márquez, A.R.; Arenas-Huertero, F. The microRNAs as potential biomarkers for predicting the onset of aflatoxin exposure in human beings: A review. Front. Microbiol. 2014, 5, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Izzotti, A.; Pulliero, A. The effects of environmental chemical carcinogens on the microRNA machinery. Int. J. Hyg. Environ. Health 2014, 217, 601–627. [Google Scholar] [CrossRef]
- Jayanthy, A.; Setaluri, V. Light-regulated MicroRNAs. Photochem. Photobiol. 2015, 91, 163–172. [Google Scholar] [CrossRef]
- Metheetrairut, C.; Slack, F.J. MicroRNAs in the ionizing radiation response and in radiotherapy. Curr. Opin. Genet. Dev. 2013, 23, 12–19. [Google Scholar] [CrossRef]
- Czochor, J.R.; Glazer, P.M. microRNAs in cancer cell response to ionizing radiation. Antioxid. Redox Signal. 2014, 21, 293–312. [Google Scholar] [CrossRef]
- Halimi, M.; Asghari, S.M.; Sariri, R.; Moslemi, D.; Parsian, H. Cellular response to ionizing radiation: A microRNA story. Int. J. Mol. Cell. Med. 2012, 1, 178–184. [Google Scholar]
- Simone, N.L.; Soule, B.P.; Ly, D.; Saleh, A.D.; Savage, J.E.; Degraff, W.; Cook, J.; Harris, C.C.; Gius, D.; Mitchell, J.B. Ionizing radiation-induced oxidative stress alters microRNA expression. PLoS ONE 2009, 4, e6377. [Google Scholar] [CrossRef]
- Shin, S.; Cha, H.J.; Lee, E.M.; Lee, S.J.; Seo, S.K.; Jin, H.O.; Park, I.C.; Jin, Y.W.; An, S. Alteration of microRNA profiles by ionizing radiation in A549 human non-small cell lung cancer cells. Int. J. Oncol. 2009, 35, 81–86. [Google Scholar]
- Templin, T.; Young, E.F.; Smilenov, L.B. Whole mouse blood microRNA as biomarkers for exposure to -rays and (56) Fe ion. Int. J. Radiat. Biol. 2011, 87, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Templin, T.; Young, E.F.; Smilenov, L.B. Proton radiation-induced microRNA signatures in mouse blood: Characterization and comparison with 56Fe-ion and gamma radiation. Int. J. Radiat. Biol. 2012, 88, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.M.; Li, J.X.; Chen, Q.; Du, H.B.; Zhang, S.Y.; Nie, J.H.; Cao, J.P.; Zhou, P.K.; Hei, T.K.; Tong, J. Radon-induced alterations in micro-RNA expression profiles in transformed BEAS2B cells. J. Toxicol. Environ. Health 2013, 76, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Bulgakova, O.; Zhabayeva, D.; Kussainova, A.; Pulliero, A.; Izzotti, A.; Bersimbaev, R. miR‑19 in blood plasma reflects lung cancer occurrence but is not specifically associated with radon exposure. Oncol. Lett. 2018, 15, 8816–8824. [Google Scholar] [CrossRef] [PubMed]
- Nymark, P.; Guled, M.; Borze, I.; Faisal, A.; Lahti, L.; Salmenkivi, K.; Kettunen, E.; Anttila, S.; Knuutila, S. Integrative analysis of microRNA, mRNA and aCGH data reveals asbestos- and histology-related changes in lung cancer. Genes Chromosom. Cancer 2011, 50, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Kettunen, E.; Aavikko, M.; Nymark, P.; Ruosaari, S.; Wikman, H.; Vanhala, E.; Salmenkivi, K.; Pirinen, R.; Karjalainen, A.; Kuosma, E.; et al. DNA copy number loss and allelic imbalance at 2p16 in lung cancer associated with asbestos exposure. Br. J. Cancer 2009, 100, 1336–1342. [Google Scholar] [CrossRef]
- Santarelli, L.; Gaetani, S.; Monaco, F.; Bracci, M.; Valentino, M.; Amati, M.; Rubini, C.; Sabbatini, A.; Pasquini, E.; Zanotta, N.; et al. Four-miRNA Signature to identify asbestos related lung malignancies. Cancer Epidemiol. Biomark. Prev. 2019, 28. [Google Scholar] [CrossRef]
- Kettunen, E.; Hernandez-Vargas, H.; Cros, M.-P.; Durand, G.; Le Calvez-Kelm, F.; Stuopelyte, K.; Jarmalaite, S.; Salmenkivi, K.; Anttila, S.; Wolff, H.; et al. Asbestos-associated genome-wide DNA methylation changes in lung cancer. Int. J. Cancer 2017, 141, 2014–2029. [Google Scholar] [CrossRef]
- Cheng, Y.Y.; Rath, E.M.; Linton, A.; Yuen, M.L.; Takahashi, K.; Lee, K. The current understanding of asbestos-induced epigenetic changes associated with lung cancer. Lung Cancer 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Oren, M. Decision making by p53: Life, death and cancer. Cell Death Differ. 2003, 10, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Andujar, P.; Pairon, J.C.; Renier, A.; Descatha, A.; Hysi, I. Differential mutation profiles and similar intronic TP53 polymorphisms in asbestos-related lung cancer and pleural mesothelioma. Mutagenesis 2013, 28, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Inamura, K.; Ninomiya, H.; Nomura, K.; Tsuchiya, E.; Satoh, Y.; Okumura, S.; Nakagawa, K.; Takata, A.; Kohyama, N.; Ishikawa, Y. Combined effects of asbestos and cigarette smoke on the development of lung adenocarcinoma: Different carcinogens may cause different genomic changes. Oncol. Rep. 2014, 32, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Panduri, V.; Surapureddi, S.; Soberanes, S.; Weitzman, S.A.; Chandel, N.; Kamp, D.W. P53 Mediates Amosite Asbestos–Induced Alveolar Epithelial Cell Mitochondria-Regulated Apoptosis. Am. J. Respir. Cell Mol. Biol. 2006, 34, 443–452. [Google Scholar] [CrossRef]
- Ju, L.; Wu, W.; Yin, X.; Xiao, Y.; Jia, Z.; Lou, J.; Zhu, L. miR-30d is related to asbestos exposure and inhibits migration and invasion in NCI-H2452 cells. FEBS Open Bio 2017, 7, 1469–1479. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, S.M.; Soltani, B.M.; Tavallaei, M.; Mowla, S.J.; Tafsiri, E.; Bagheri, A.; Khorshid, H.R.K. Clinically Significant Dysregulation of hsa-miR-30d-5p and hsa-let-7b Expression in Patients with Surgically Resected Non-Small Cell Lung Cancer. Avicenna J. Med. Biotechnol. 2018, 10, 98–104. [Google Scholar]
- Gao, L.; He, R.; Wu, H.-Y.; Zhang, T.-T.; Liang, H.; Ye, Z.-H.; Li, Z.-Y.; Xie, T.-T.; Shi, Q.; Ma, J.; et al. Expression Signature and Role of miR-30d-5p in Non-Small Cell Lung Cancer: A Comprehensive Study Based on in Silico Analysis of Public Databases and In Vitro Experiments. Cell Physiol. Biochem. 2018, 50, 1964–1987. [Google Scholar] [CrossRef]
- Luan, N.; Wang, Y.; Liu, X. Absent expression of miR-30a promotes the growth of lung cancer cells by targeting MEF2D. Oncol. Lett. 2018, 16, 1173–1179. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.-X.; Shi, L.; Zhang, Y.; Zhu, Y.; Bai, C.; Wang, X.; Zhou, J.-B. Myocyte enhancer factor 2D provides a cross-talk between chronic inflammation and lung cancer. J. Transl. Med. 2015, 15, 65. [Google Scholar] [CrossRef]
- Hylebos, M.; Guy, V.C. The Genetic Landscape of Malignant Pleural Mesothelioma: Results from Massively Parallel Sequencing. J. Thorac. Oncol. 2016, 11, 1615–1626. [Google Scholar] [CrossRef]
- Menges, C.W.; Kadariya, Y.; Altomare, D.; Kadariya, Y.; Altomare, D. Tumor suppressor alterations cooperate to drive aggressive mesotheliomas with enriched cancer stem cells via a p53-miR-34a-c-Met axis. Cancer Res. 2014, 74, 1261–1271. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.H.; Kim, H.S.; Li, X.Y.; Lee, I.; Choi, H.S.; Kang, S.E.; Cha, S.Y.; Ryu, J.K.; Yoon, D.; Fearon, E.R.; et al. A p53/microRNA-34 axis regulates Snail1-dependent cancer cell epithelial-mesenchymal transition. J. Cell Biol. 2011, 195, 417–433. [Google Scholar] [CrossRef] [PubMed]
- Muraoka, T.; Soh, J.; Toyooka, S.; Aoe, K.; Fujimoto, N.; Hashida, S.; Maki, Y.; Tanaka, N.; Shien, K.; Furukawa, M.; et al. The degree of microRNA-34b/c methylation in serum-circulating DNA is associated with malignant pleural mesothelioma. Lung Cancer 2013, 82, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.Y.; Huang, J.S.; Shiah, S.; Chung, S.Y.; Lay, J.D.; Yang, Y.Y.; Lai, G.M.; Cheng, A.L.; Chen, L.T.; Chuang, S.E. Negative feedback regulation of AXL by miR-34a modulates apoptosis in lung cancer cells. RNA 2016, 22, 303–315. [Google Scholar] [CrossRef]
- Weber, D.G.; Johnen, G.; Bryk, O.; Jöckel, K.H.; Brüning, T. Identification of miRNA-103 in the cellular fraction of human peripheral blood as a potential biomarker for malignant mesothelioma—A pilot study. PLoS ONE 2012, 7, e30221. [Google Scholar] [CrossRef]
- Okayasu, R.; Takahashi, S.; Yamada, S.; Hei, T.K.; Ullrich, R.L. Asbestos and DNA double strand breaks. Cancer Res. 1999, 59, 298–300. [Google Scholar]
- Castranova, V. Signaling pathways controlling the production of inflammatory mediators in response to crystalline silica exposure: Role of reactive oxygen/nitrogen species. Free Radic Biol. Med. 2004, 37, 916–925. [Google Scholar] [CrossRef]
- Msiska, Z.; Pacurari, M.; Mishra, A.; Leonard, S.S.; Castranova, V.; Vallyathan, V. DNA double-strand breaks by asbestos, silica, and titanium dioxide: Possible biomarker of carcinogenic potential? Am. J. Respir. Cell Mol. Biol. 2010, 43, 210–219. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, L.; Duan, L.; Lu, C. MicroRNA-3196 is inhibited by H2AX phosphorylation and attenuates lung cancer cell apoptosis by downregulating PUMA. Oncotarget 2016, 7, 77764–77776. [Google Scholar] [CrossRef]
- Micolucci, L.; Akhtar, M.M.; Olivieri, F.; Rippo, M.R.; Procopio, A.D. Diagnostic value of microRNAs in asbestos exposure and malignant mesothelioma: Systematic review and qualitative meta-analysis. Oncotarget 2016, 7, 58606–58637. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bersimbaev, R.; Bulgakova, O.; Aripova, A.; Kussainova, A.; Ilderbayev, O. Role of microRNAs in Lung Carcinogenesis Induced by Asbestos. J. Pers. Med. 2021, 11, 97. https://doi.org/10.3390/jpm11020097
Bersimbaev R, Bulgakova O, Aripova A, Kussainova A, Ilderbayev O. Role of microRNAs in Lung Carcinogenesis Induced by Asbestos. Journal of Personalized Medicine. 2021; 11(2):97. https://doi.org/10.3390/jpm11020097
Chicago/Turabian StyleBersimbaev, Rakhmetkazhy, Olga Bulgakova, Akmaral Aripova, Assiya Kussainova, and Oralbek Ilderbayev. 2021. "Role of microRNAs in Lung Carcinogenesis Induced by Asbestos" Journal of Personalized Medicine 11, no. 2: 97. https://doi.org/10.3390/jpm11020097
APA StyleBersimbaev, R., Bulgakova, O., Aripova, A., Kussainova, A., & Ilderbayev, O. (2021). Role of microRNAs in Lung Carcinogenesis Induced by Asbestos. Journal of Personalized Medicine, 11(2), 97. https://doi.org/10.3390/jpm11020097