
Performance of T-Track® SARS-CoV-2, an Innovative Dual Marker RT-qPCR-Based Whole-Blood Assay for the Detection of SARS-CoV-2-Reactive T Cells


Abstract
:1. Introduction
2. Materials and Methods
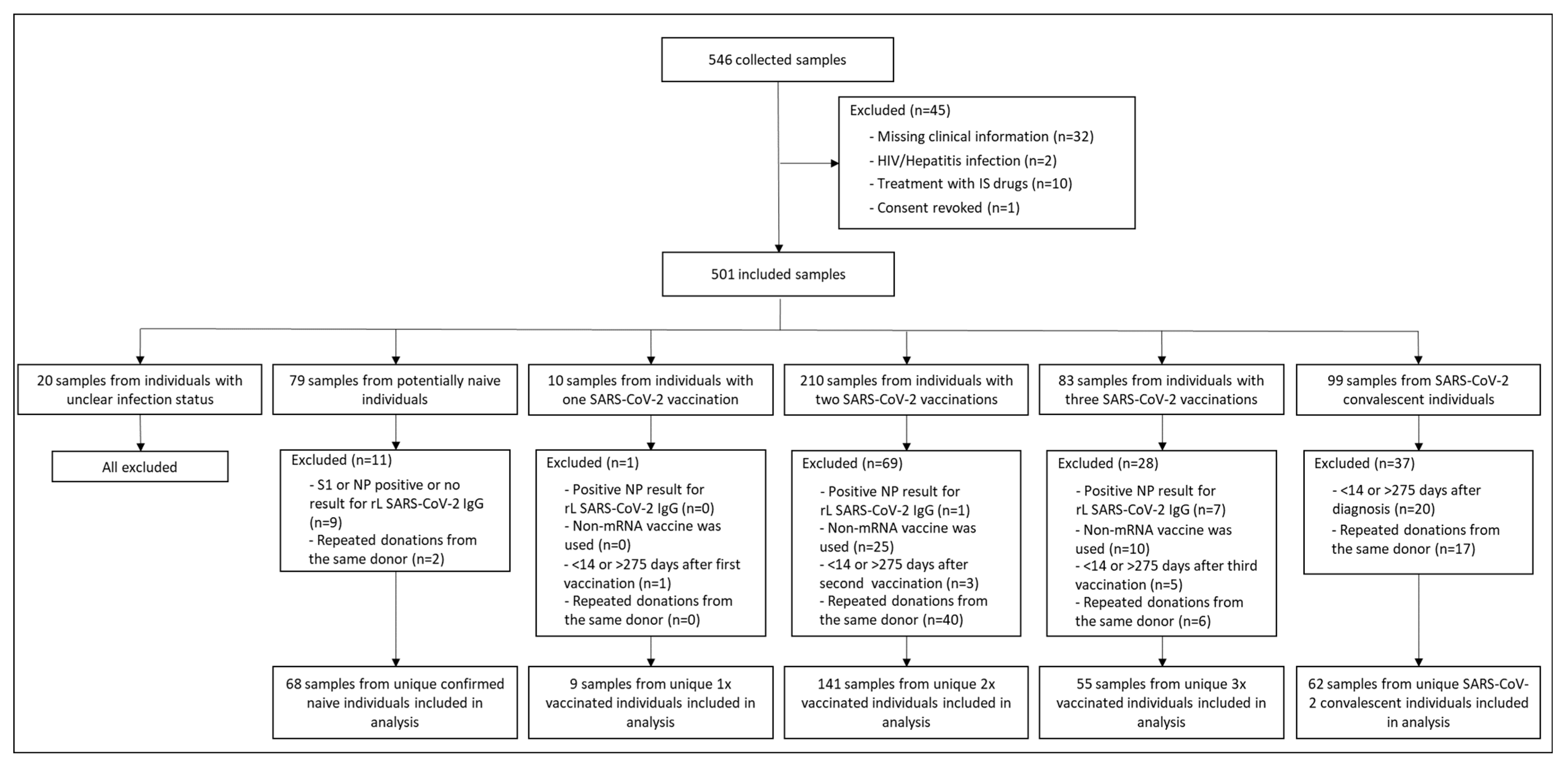
2.1. Study Design and Participants
2.2. Sample Collection
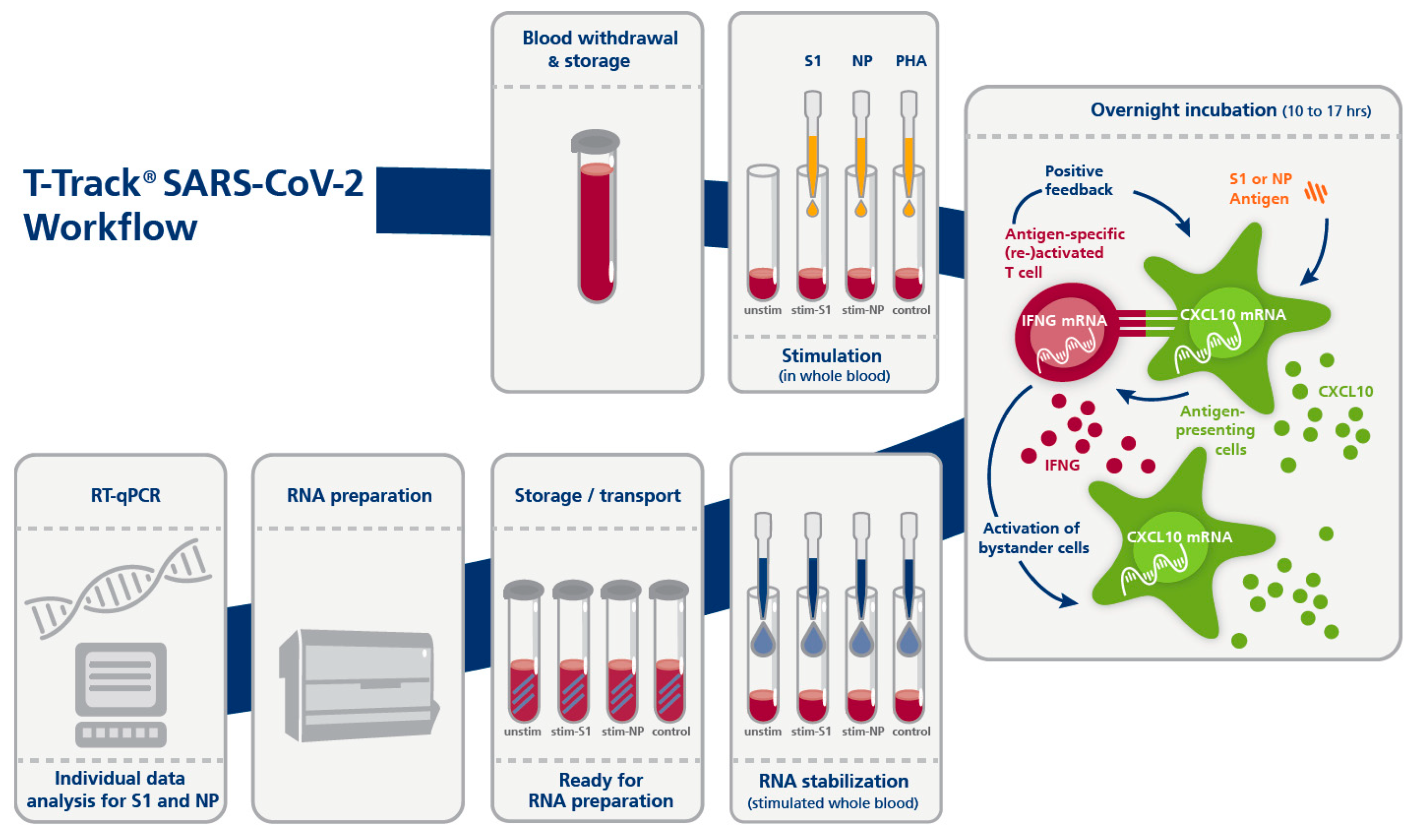
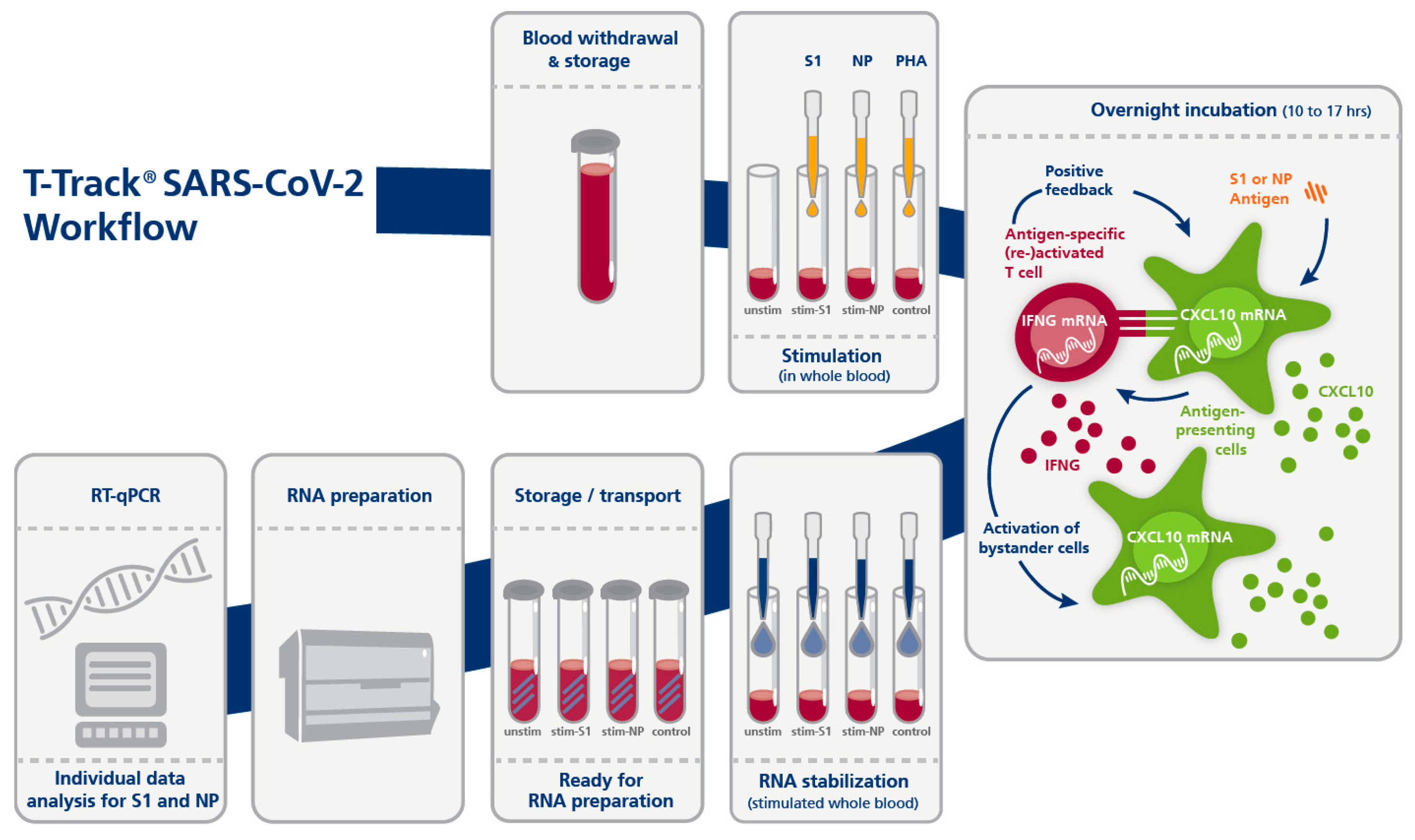
2.3. T-Track® SARS-CoV-2 Assay
2.4. recomLine SARS-CoV-2 IgG
2.5. Statistical Analysis
3. Results
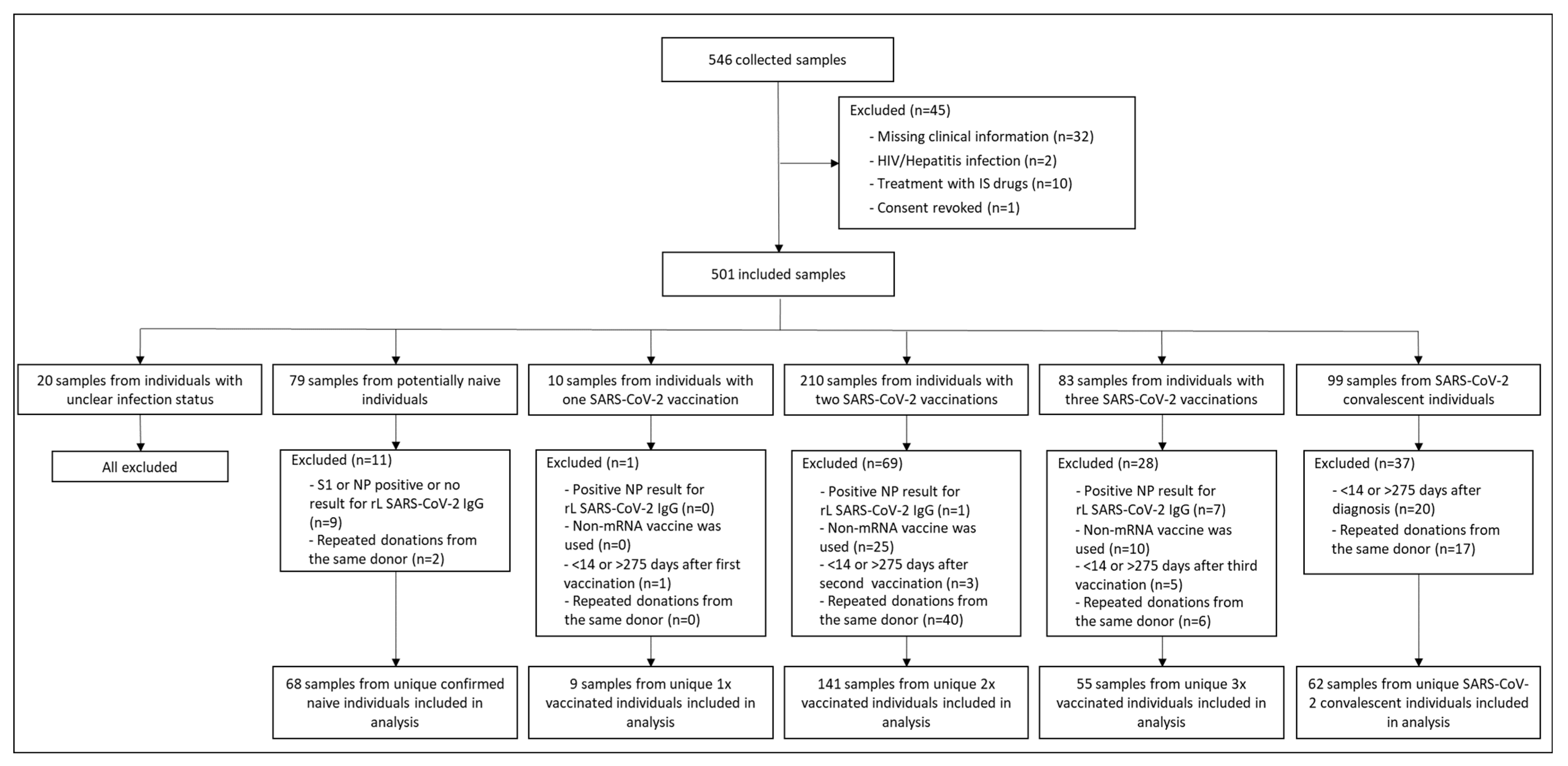
3.1. Study Participants’ Characteristics
3.2. Diagnostic Performance of T-Track® SARS-CoV-2
3.3. Age-Related Performance of T-Track® SARS-CoV-2
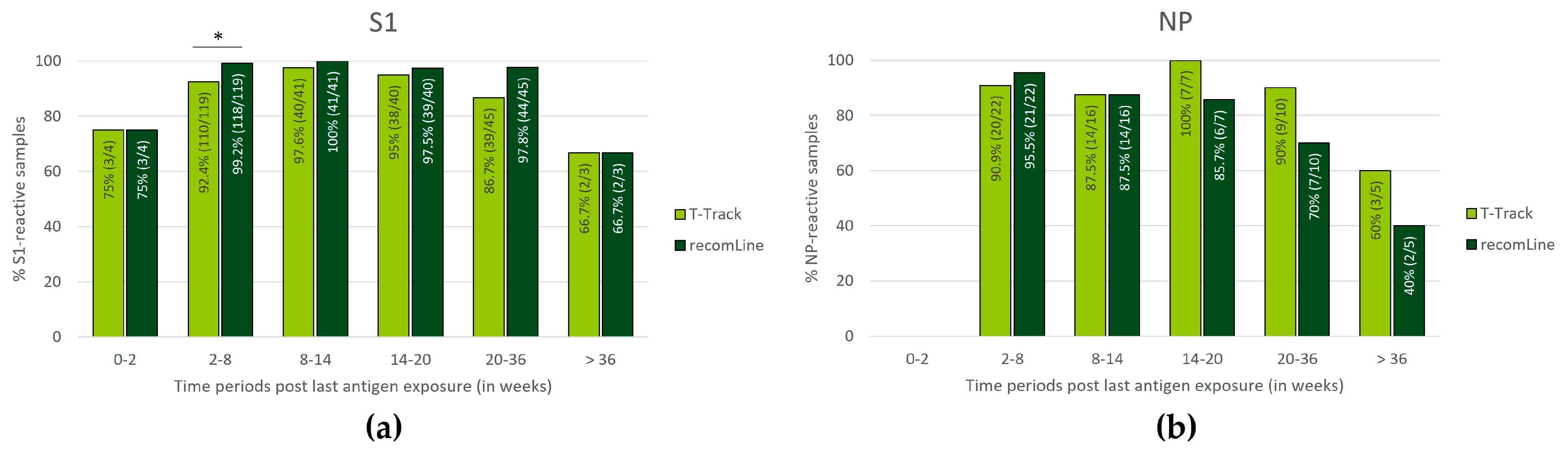
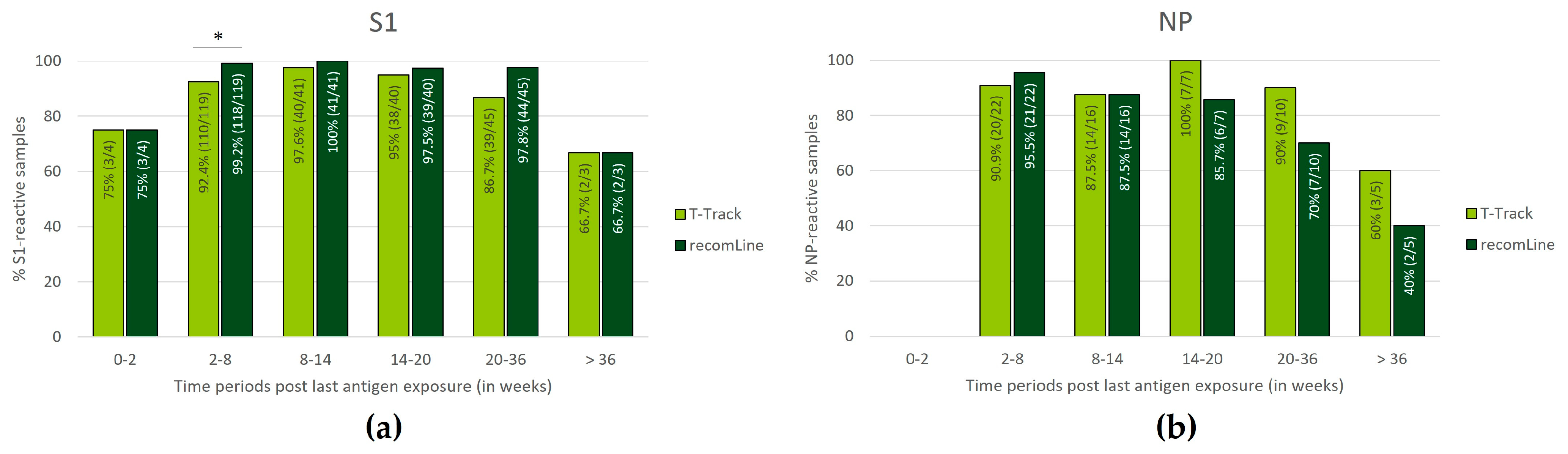
3.4. Persistence of SARS-CoV-2-Specific Humoral and Cellular Immune Responses
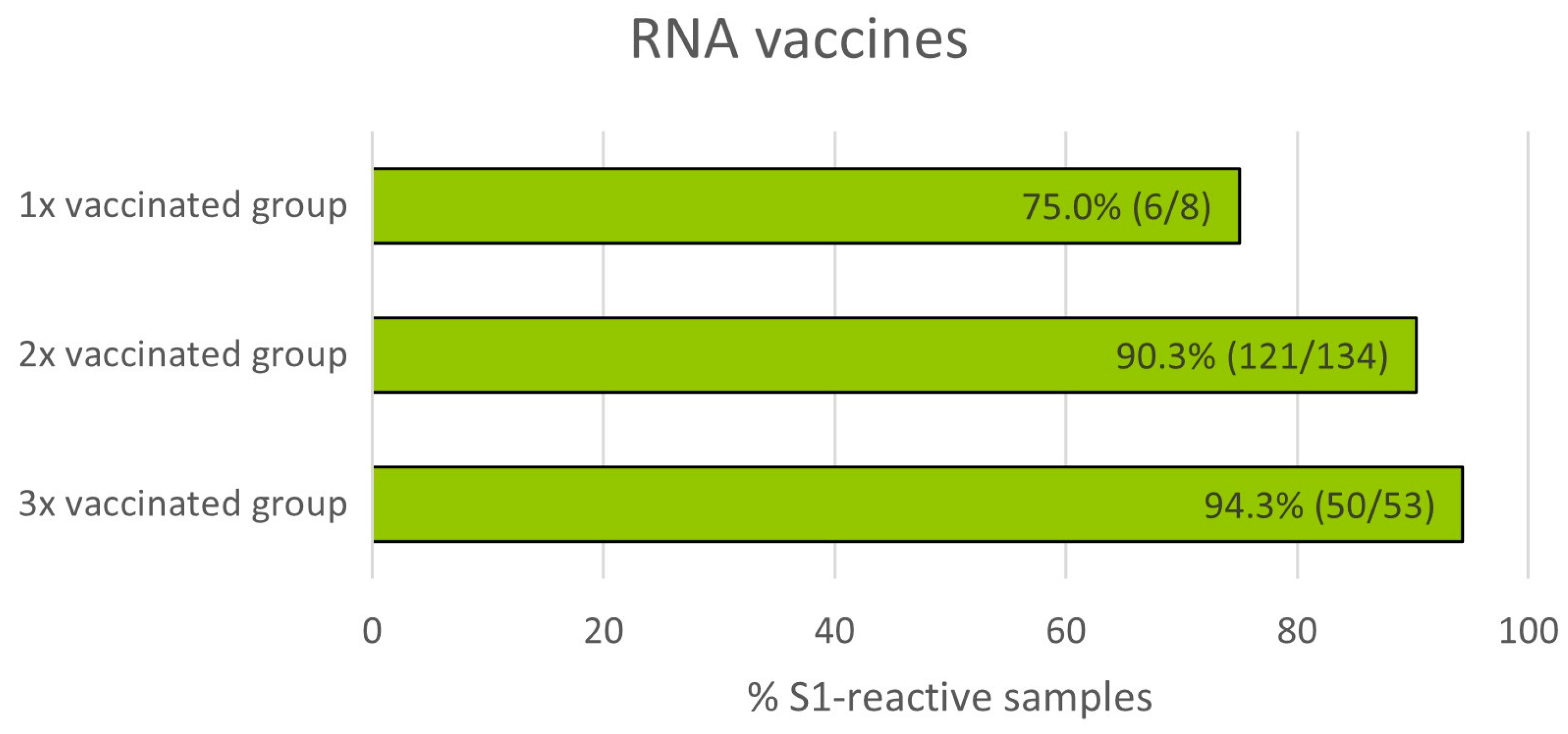
3.5. Impact of Number of mRNA Vaccinations on T Cell Reactivity
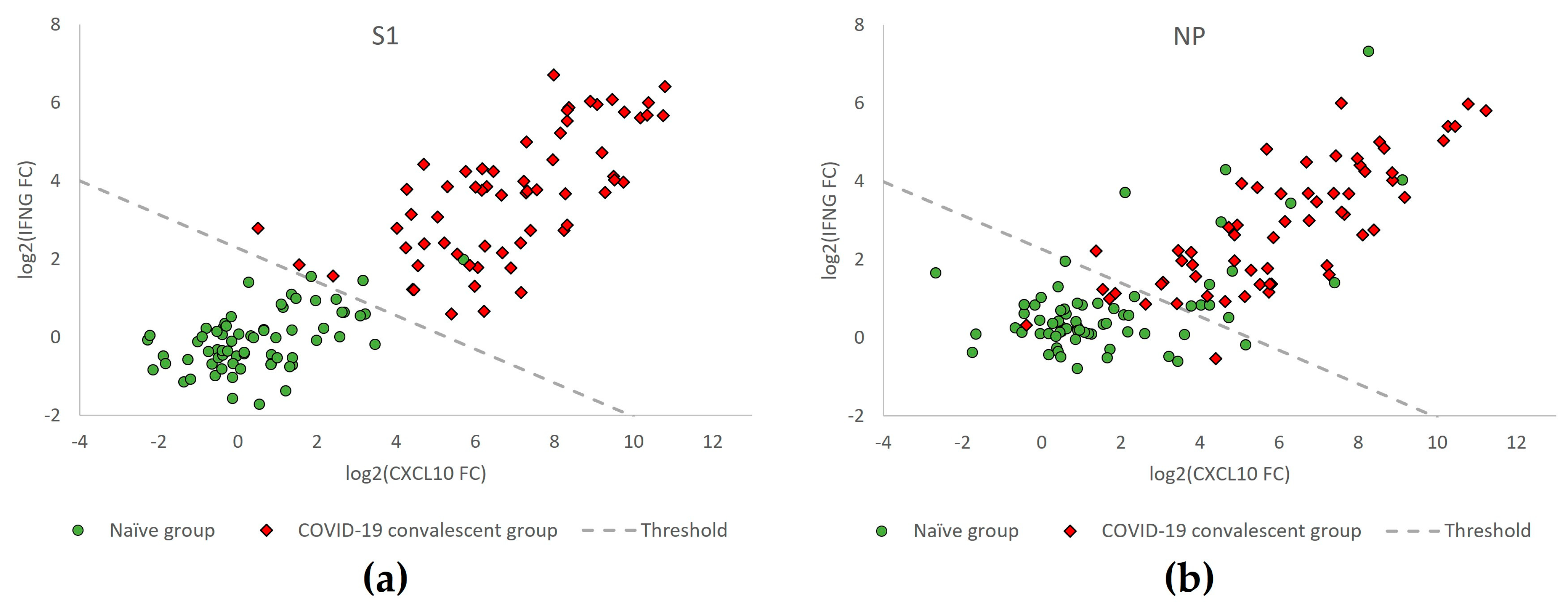
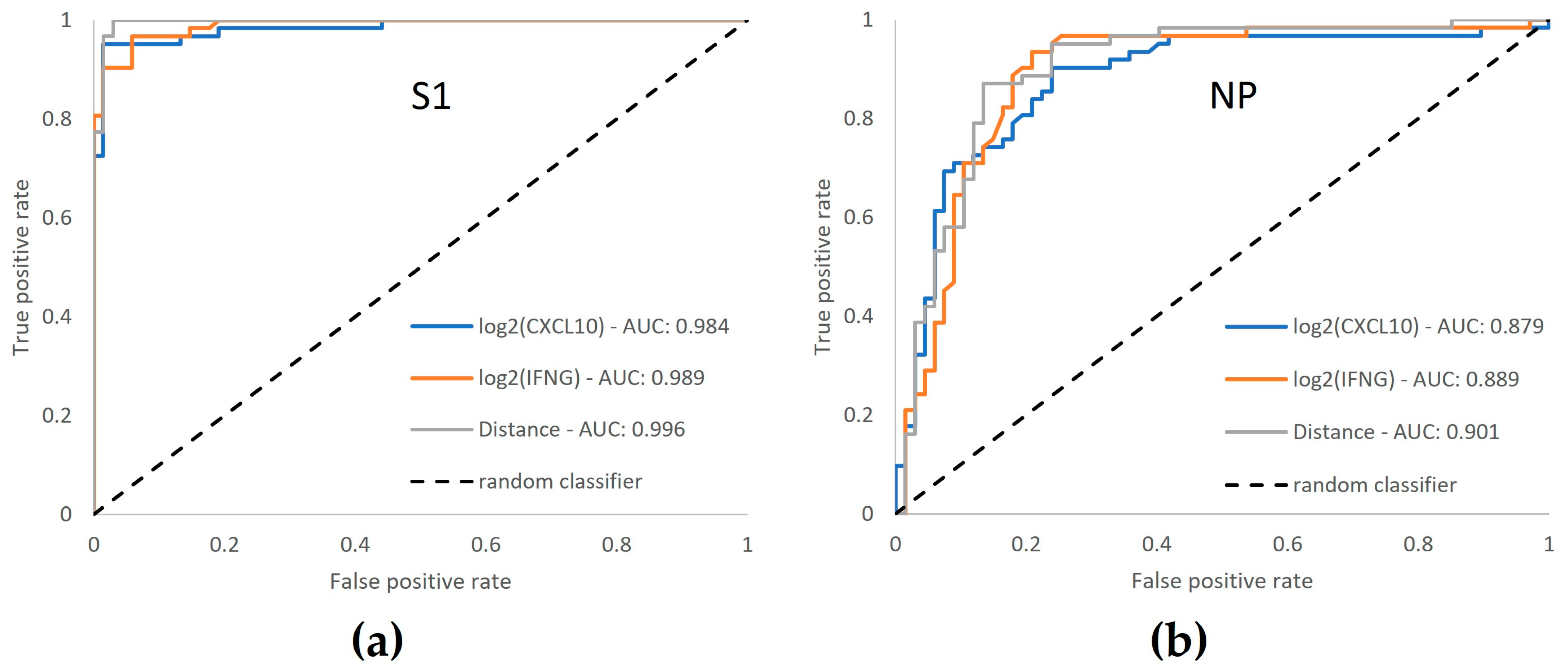
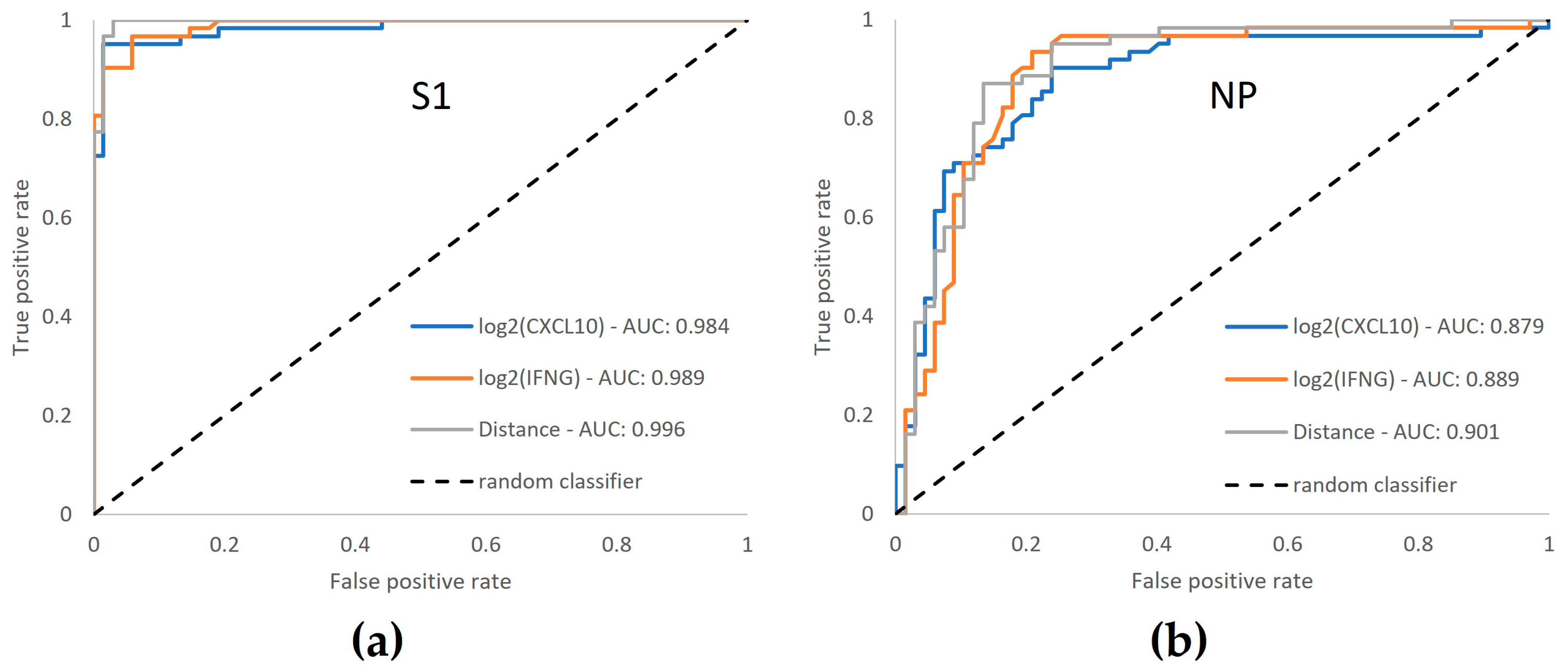
3.6. ROC Curve Analysis Confirms Enhanced Performance by Dual-Marker Analysis
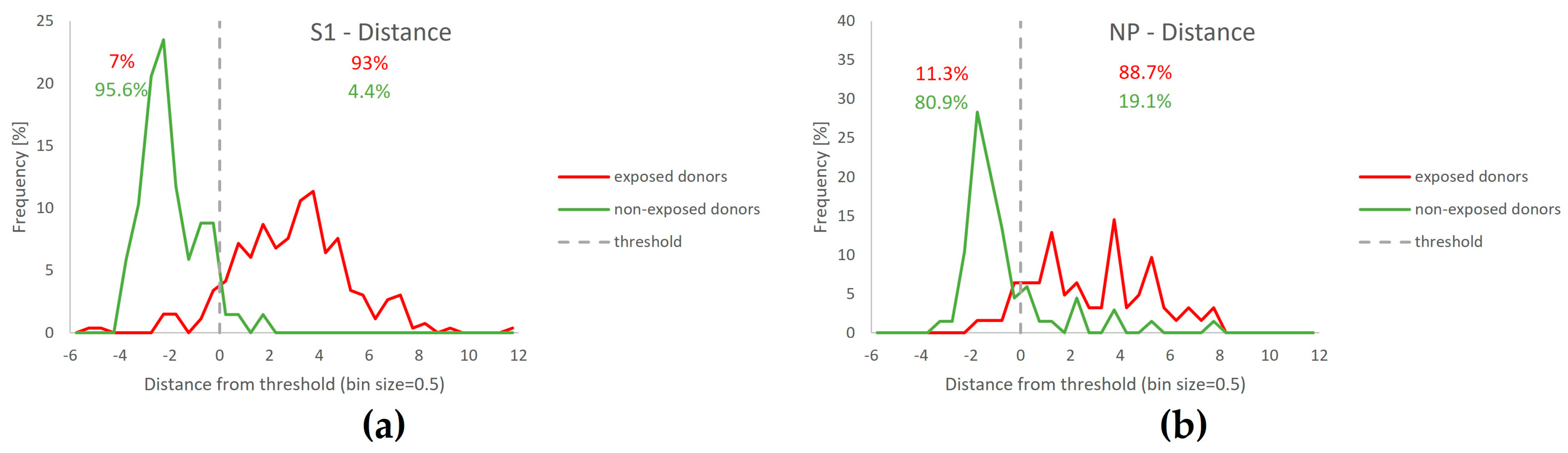
3.7. Separation of T-Track® SARS-CoV-2 Test Results Relative to the Classification Threshold, According to Donors’ SARS-CoV-2 Exposure Status
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sagar, M.; Reifler, K.; Rossi, M.; Miller, N.S.; Sinha, P.; White, L.F.; Mizgerd, J.P. Recent Endemic Coronavirus Infection Is Associated with Less-Severe COVID-19. J. Clin. Investig. 2021, 131, e143380. [Google Scholar] [CrossRef] [PubMed]
- Swadling, L.; Diniz, M.O.; Schmidt, N.M.; Amin, O.E.; Chandran, A.; Shaw, E.; Pade, C.; Gibbons, J.M.; Le Bert, N.; Tan, A.T.; et al. Pre-Existing Polymerase-Specific T Cells Expand in Abortive Seronegative SARS-CoV-2. Nature 2022, 601, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Loyal, L.; Braun, J.; Henze, L.; Kruse, B.; Dingeldey, M.; Reimer, U.; Kern, F.; Schwarz, T.; Mangold, M.; Unger, C.; et al. Cross-Reactive CD4 + T Cells Enhance SARS-CoV-2 Immune Responses upon Infection and Vaccination. Science 2021, 374, eabh1823. [Google Scholar] [CrossRef] [PubMed]
- Kundu, R.; Narean, J.S.; Wang, L.; Fenn, J.; Pillay, T.; Fernandez, N.D.; Conibear, E.; Koycheva, A.; Davies, M.; Tolosa-Wright, M.; et al. Cross-Reactive Memory T Cells Associate with Protection against SARS-CoV-2 Infection in COVID-19 Contacts. Nat. Commun. 2022, 13, 80. [Google Scholar] [CrossRef] [PubMed]
- Swadling, L.; Maini, M.K. T Cells in COVID-19—United in Diversity. Nat. Immunol. 2020, 21, 1307–1308. [Google Scholar] [CrossRef] [PubMed]
- Sette, A.; Crotty, S. Adaptive Immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880. [Google Scholar] [CrossRef] [PubMed]
- Moss, P. The T Cell Immune Response against SARS-CoV-2. Nat. Immunol. 2022, 23, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, M.; Rabaan, A.A.; Fawarah, M.M.A.; Almuthree, S.A.; Alsubki, R.A.; Alfaraj, A.H.; Mashraqi, M.M.; Alshamrani, S.A.; Abduljabbar, W.A.; Alwashmi, A.S.S.; et al. Updated Insights into the T Cell-Mediated Immune Response against SARS-CoV-2: A Step towards Efficient and Reliable Vaccines. Vaccines 2023, 11, 101. [Google Scholar] [CrossRef]
- Dan, J.M.; Mateus, J.; Kato, Y.; Hastie, K.M.; Yu, E.D.; Faliti, C.E.; Grifoni, A.; Ramirez, S.I.; Haupt, S.; Frazier, A.; et al. Immunological Memory to SARS-CoV-2 Assessed for up to 8 Months after Infection. Science 2021, 371, eabf4063. [Google Scholar] [CrossRef]
- Wajnberg, A.; Amanat, F.; Firpo, A.; Altman, D.R.; Bailey, M.J.; Mansour, M.; McMahon, M.; Meade, P.; Mendu, D.R.; Muellers, K.; et al. Robust Neutralizing Antibodies to SARS-CoV-2 Infection Persist for Months. Science 2020, 370, 1227–1230. [Google Scholar] [CrossRef]
- Tang, F.; Quan, Y.; Xin, Z.-T.; Wrammert, J.; Ma, M.-J.; Lv, H.; Wang, T.-B.; Yang, H.; Richardus, J.H.; Liu, W.; et al. Lack of Peripheral Memory B Cell Responses in Recovered Patients with Severe Acute Respiratory Syndrome: A Six-Year Follow-up Study. J. Immunol. 2011, 186, 7264–7268. [Google Scholar] [CrossRef] [PubMed]
- Sekine, T.; Perez-Potti, A.; Rivera-Ballesteros, O.; Strålin, K.; Gorin, J.-B.; Olsson, A.; Llewellyn-Lacey, S.; Kamal, H.; Bogdanovic, G.; Muschiol, S.; et al. Robust T Cell Immunity in Convalescent Individuals with Asymptomatic or Mild COVID-19. Cell 2020, 183, 158–168.e14. [Google Scholar] [CrossRef] [PubMed]
- Nelde, A.; Bilich, T.; Heitmann, J.S.; Maringer, Y.; Salih, H.R.; Roerden, M.; Lübke, M.; Bauer, J.; Rieth, J.; Wacker, M.; et al. SARS-CoV-2-Derived Peptides Define Heterologous and COVID-19-Induced T Cell Recognition. Nat. Immunol. 2021, 22, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Schwarzkopf, S.; Krawczyk, A.; Knop, D.; Klump, H.; Heinold, A.; Heinemann, F.M.; Thümmler, L.; Temme, C.; Breyer, M.; Witzke, O.; et al. Cellular Immunity in COVID-19 Convalescents with PCR-Confirmed Infection but with Undetectable SARS-CoV-2–Specific IgG. Emerg. Infect. Dis. 2021, 27, 122. [Google Scholar] [CrossRef]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef] [PubMed]
- Sahin, U.; Muik, A.; Derhovanessian, E.; Vogler, I.; Kranz, L.M.; Vormehr, M.; Baum, A.; Pascal, K.; Quandt, J.; Maurus, D.; et al. COVID-19 Vaccine BNT162b1 Elicits Human Antibody and TH1 T Cell Responses. Nature 2020, 586, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Oberhardt, V.; Luxenburger, H.; Kemming, J.; Schulien, I.; Ciminski, K.; Giese, S.; Csernalabics, B.; Lang-Meli, J.; Janowska, I.; Staniek, J.; et al. Rapid and Stable Mobilization of CD8+ T Cells by SARS-CoV-2 MRNA Vaccine. Nature 2021, 597, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Braun, J.; Loyal, L.; Frentsch, M.; Wendisch, D.; Georg, P.; Kurth, F.; Hippenstiel, S.; Dingeldey, M.; Kruse, B.; Fauchere, F.; et al. SARS-CoV-2-Reactive T Cells in Healthy Donors and Patients with COVID-19. Nature 2020, 587, 270–274. [Google Scholar] [CrossRef]
- Le Bert, N.; Tan, A.T.; Kunasegaran, K.; Tham, C.Y.L.; Hafezi, M.; Chia, A.; Chng, M.H.Y.; Lin, M.; Tan, N.; Linster, M.; et al. SARS-CoV-2-Specific T Cell Immunity in Cases of COVID-19 and SARS, and Uninfected Controls. Nature 2020, 584, 457–462. [Google Scholar] [CrossRef]
- Rydyznski Moderbacher, C.; Ramirez, S.I.; Dan, J.M.; Grifoni, A.; Hastie, K.M.; Weiskopf, D.; Belanger, S.; Abbott, R.K.; Kim, C.; Choi, J.; et al. Antigen-Specific Adaptive Immunity to SARS-CoV-2 in Acute COVID-19 and Associations with Age and Disease Severity. Cell 2020, 183, 996–1012.e19. [Google Scholar] [CrossRef]
- Schulien, I.; Kemming, J.; Oberhardt, V.; Wild, K.; Seidel, L.M.; Killmer, S.; Sagar, N.; Daul, F.; Salvat Lago, M.; Decker, A.; et al. Characterization of Pre-Existing and Induced SARS-CoV-2-Specific CD8+ T Cells. Nat. Med. 2021, 27, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Soresina, A.; Moratto, D.; Chiarini, M.; Paolillo, C.; Baresi, G.; Focà, E.; Bezzi, M.; Baronio, B.; Giacomelli, M.; Badolato, R. Two X-Linked Agammaglobulinemia Patients Develop Pneumonia as COVID-19 Manifestation but Recover. Pediatr. Allergy Immunol. 2020, 31, 565–569. [Google Scholar] [CrossRef]
- Geers, D.; Shamier, M.C.; Bogers, S.; den Hartog, G.; Gommers, L.; Nieuwkoop, N.N.; Schmitz, K.S.; Rijsbergen, L.C.; van Osch, J.A.T.; Dijkhuizen, E.; et al. SARS-CoV-2 Variants of Concern Partially Escape Humoral but Not T-Cell Responses in COVID-19 Convalescent Donors and Vaccinees. Sci. Immunol. 2021, 6, eabj1750. [Google Scholar] [CrossRef] [PubMed]
- Kalimuddin, S.; Tham, C.Y.L.; Qui, M.; de Alwis, R.; Sim, J.X.Y.; Lim, J.M.E.; Tan, H.-C.; Syenina, A.; Zhang, S.L.; Le Bert, N.; et al. Early T Cell and Binding Antibody Responses Are Associated with COVID-19 RNA Vaccine Efficacy Onset. Med 2021, 2, 682–688.e4. [Google Scholar] [CrossRef] [PubMed]
- Kruse, M.; Dark, C.; Aspden, M.; Cochrane, D.; Competiello, R.; Peltz, M.; Torres, L.; Wrighton-Smith, P.; Dudek, M. Performance of the T-SPOT®. COVID Test for Detecting SARS-CoV-2-Responsive T Cells. Int. J. Infect. Dis. 2021, 113, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Krüttgen, A.; Klingel, H.; Haase, G.; Haefner, H.; Imöhl, M.; Kleines, M. Evaluation of the QuantiFERON SARS-CoV-2 Interferon-ɣ Release Assay in MRNA-1273 Vaccinated Health Care Workers. J. Virol. Methods 2021, 298, 114295. [Google Scholar] [CrossRef] [PubMed]
- Lochmanová, A.; Martinek, J.; Tomášková, H.; Zelená, H.; Dieckmann, K.; Grage-Griebenow, E.; Ježo, E.; Janošek, J. Comparison of Two Commercially Available Interferon-γ Release Assays for T-Cell-Mediated Immunity and Evaluation of Humoral Immunity against SARS-CoV-2 in Healthcare Workers. Diagnostics 2023, 13, 637. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.T.; Linster, M.; Tan, C.W.; Le Bert, N.; Chia, W.N.; Kunasegaran, K.; Zhuang, Y.; Tham, C.Y.L.; Chia, A.; Smith, G.J.D.; et al. Early Induction of Functional SARS-CoV-2-Specific T Cells Associates with Rapid Viral Clearance and Mild Disease in COVID-19 Patients. Cell Rep. 2021, 34, 108728. [Google Scholar] [CrossRef]
- Zhang, N.; Zhao, Y.-D. CXCL10 an Important Chemokine Associated with Cytokine Storm in COVID-19 Infected Patients. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 7497–7505. [Google Scholar] [CrossRef]
- Lu, Z.; Laing, E.D.; Pena DaMata, J.; Pohida, K.; Tso, M.S.; Samuels, E.C.; Epsi, N.J.; Dorjbal, B.; Lake, C.; Richard, S.A.; et al. Durability of SARS-CoV-2–Specific T-Cell Responses at 12 Months Postinfection. J. Infect. Dis. 2021, 224, 2010–2019. [Google Scholar] [CrossRef]
- Hou, H.; Zhang, Y.; Tang, G.; Luo, Y.; Liu, W.; Cheng, C.; Jiang, Y.; Xiong, Z.; Wu, S.; Sun, Z.; et al. Immunologic Memory to SARS-CoV-2 in Convalescent COVID-19 Patients at 1 Year Postinfection. J. Allergy Clin. Immunol. 2021, 148, 1481–1492.e2. [Google Scholar] [CrossRef] [PubMed]
- Meier, J.P.; Möbus, S.; Heigl, F.; Asbach-Nitzsche, A.; Niller, H.H.; Plentz, A.; Avsar, K.; Heiß-Neumann, M.; Schaaf, B.; Cassens, U.; et al. Performance of T-Track® TB, a Novel Dual Marker RT-QPCR-Based Whole-Blood Test for Improved Detection of Active Tuberculosis. Diagnostics 2023, 13, 758. [Google Scholar] [CrossRef] [PubMed]
- Agresti, A.; Coull, B.A. Approximate Is Better than “Exact” for Interval Estimation of Binomial Proportions. Am. Stat. 1998, 52, 119–126. [Google Scholar] [CrossRef]
- Hanley, J.A.; McNeil, B.J. A Method of Comparing the Areas under Receiver Operating Characteristic Curves Derived from the Same Cases. Radiology 1983, 148, 839–843. [Google Scholar] [CrossRef] [PubMed]
- Bieberich, F.; Vazquez-Lombardi, R.; Yermanos, A.; Ehling, R.A.; Mason, D.M.; Wagner, B.; Kapetanovic, E.; Di Roberto, R.B.; Weber, C.R.; Savic, M.; et al. A Single-Cell Atlas of Lymphocyte Adaptive Immune Repertoires and Transcriptomes Reveals Age-Related Differences in Convalescent COVID-19 Patients. Front. Immunol. 2021, 12, 701085. [Google Scholar] [CrossRef] [PubMed]
- Dietz, L.L.; Juhl, A.K.; Søgaard, O.S.; Reekie, J.; Nielsen, H.; Johansen, I.S.; Benfield, T.; Wiese, L.; Stærke, N.B.; Jensen, T.Ø.; et al. Impact of Age and Comorbidities on SARS-CoV-2 Vaccine-Induced T Cell Immunity. Commun. Med. 2023, 3, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, M.; Torre, D.; Lozano-Ojalvo, D.; Tan, A.T.; Tabaglio, T.; Mzoughi, S.; Sanchez-Tarjuelo, R.; Le Bert, N.; Lim, J.M.E.; Hatem, S.; et al. Rapid, Scalable Assessment of SARS-CoV-2 Cellular Immunity by Whole-Blood PCR. Nat. Biotechnol. 2022, 40, 1680–1689. [Google Scholar] [CrossRef] [PubMed]
- Cohen, K.W.; Linderman, S.L.; Moodie, Z.; Czartoski, J.; Lai, L.; Mantus, G.; Norwood, C.; Nyhoff, L.E.; Edara, V.V.; Floyd, K.; et al. Longitudinal Analysis Shows Durable and Broad Immune Memory after SARS-CoV-2 Infection with Persisting Antibody Responses and Memory B and T Cells. Cell Rep. Med. 2021, 2, 100354. [Google Scholar] [CrossRef]
- Aubry, A.; Demey, B.; François, C.; Duverlie, G.; Castelain, S.; Helle, F.; Brochot, E. Longitudinal Analysis and Comparison of Six Serological Assays up to Eight Months Post-COVID-19 Diagnosis. JCM 2021, 10, 1815. [Google Scholar] [CrossRef]
- Jo, N.; Hidaka, Y.; Kikuchi, O.; Fukahori, M.; Sawada, T.; Aoki, M.; Yamamoto, M.; Nagao, M.; Morita, S.; Nakajima, T.E.; et al. Impaired CD4+ T Cell Response in Older Adults Is Associated with Reduced Immunogenicity and Reactogenicity of MRNA COVID-19 Vaccination. Nat. Aging 2023, 3, 82–92. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control (ECDC) High-Risk Groups for COVID-19. Available online: https://www.ecdc.europa.eu/en/covid-19/high-risk-groups (accessed on 16 May 2023).
- Lebrasseur, A.; Fortin-Bédard, N.; Lettre, J.; Raymond, E.; Bussières, E.-L.; Lapierre, N.; Faieta, J.; Vincent, C.; Duchesne, L.; Ouellet, M.-C.; et al. Impact of the COVID-19 Pandemic on Older Adults: Rapid Review. JMIR Aging 2021, 4, e26474. [Google Scholar] [CrossRef] [PubMed]
- The Novel Coronavirus Pneumonia Emergency Response Epidemiology Team. The Epidemiological Characteristics of an Outbreak of 2019 Novel Coronavirus Diseases (COVID-19)—China, 2020. CCDCW 2020, 2, 113–122. [Google Scholar] [CrossRef]
- Stuart, A.; Katz, D.; Stevenson, C.; Gooch, D.; Harkin, L.; Bennasar, M.; Sanderson, L.; Liddle, J.; Bennaceur, A.; Levine, M.; et al. Loneliness in Older People and COVID-19: Applying the Social Identity Approach to Digital Intervention Design. Comput. Hum. Behav. Rep. 2022, 6, 100179. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.T.; Garcia-Carreras, B.; Hitchings, M.D.T.; Yang, B.; Katzelnick, L.C.; Rattigan, S.M.; Borgert, B.A.; Moreno, C.A.; Solomon, B.D.; Trimmer-Smith, L.; et al. A Systematic Review of Antibody Mediated Immunity to Coronaviruses: Kinetics, Correlates of Protection, and Association with Severity. Nat. Commun. 2020, 11, 4704. [Google Scholar] [CrossRef] [PubMed]
- Irrgang, P.; Gerling, J.; Kocher, K.; Lapuente, D.; Steininger, P.; Habenicht, K.; Wytopil, M.; Beileke, S.; Schäfer, S.; Zhong, J.; et al. Class Switch toward Noninflammatory, Spike-Specific IgG4 Antibodies after Repeated SARS-CoV-2 MRNA Vaccination. Sci. Immunol. 2023, 8, eade2798. [Google Scholar] [CrossRef]
- Saggau, C.; Martini, G.R.; Rosati, E.; Meise, S.; Messner, B.; Kamps, A.-K.; Bekel, N.; Gigla, J.; Rose, R.; Voß, M.; et al. The Pre-Exposure SARS-CoV-2-Specific T Cell Repertoire Determines the Quality of the Immune Response to Vaccination. Immunity 2022, 55, 1924–1939.e5. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Study Group | ||||
|---|---|---|---|---|---|
| Naїve | 1× Vaccinated | 2× Vaccinated | 3× Vaccinated | COVID-19 Convalescent | |
| Study population, N (%) | 68 (100%) | 9 (100%) | 141 (100%) | 55 (100%) | 62 (100%) |
| Age in years, median (range) | 49.5 (18–87) | 43 (30–61) | 40 (20–86) | 43 (21–67) | 38.5 (20–68) |
| Sex, N (%) | |||||
| Female | 40 (58.8%) | 7 (77.8%) | 99 (70.2%) | 38 (69.1%) | 41 (66.1%) |
| Male | 28 (41.2%) | 2 (22.2%) | 42 (29.8%) | 17 (30.9%) | 21 (33.9%) |
| Time since diagnosis in days, median (range) | n.a. | n.a. | n.a. | n.a. | 62 (17–240) |
| Time since last immunisation in days, median (range) | n.a. | 20 (16–35) | 97 (17–197) | 32 (15–190) | n.a. |
| Antigen | Samples with Reactive T Cells, n/N (%) (95% CI) | Invalid Rate n/N (%) | Inconclusive Rate n/N (%) | |
|---|---|---|---|---|
| Naїve Group | COVID-19 Convalescent Group | |||
| S1 | 3/68 (4.4%) (1.0–12.7) | 62/62 (100.0%) (93.0–100.0) | 0/130 (0.0%) | 0/130 (0.0%) |
| NP | 13/68 (19.1%) (11.4–30.1) | 55/62 (88.7%) (78.2–94.7) | 0/130 (0.0%) | 0/130 (0.0%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanis, F.M.; Meier, J.P.; Guldan, H.; Niller, H.-H.; Dahm, M.; Dansard, A.; Zander, T.; Struck, F.; Soutschek, E.; Deml, L.; et al. Performance of T-Track® SARS-CoV-2, an Innovative Dual Marker RT-qPCR-Based Whole-Blood Assay for the Detection of SARS-CoV-2-Reactive T Cells. Diagnostics 2023, 13, 2722. https://doi.org/10.3390/diagnostics13172722
Kanis FM, Meier JP, Guldan H, Niller H-H, Dahm M, Dansard A, Zander T, Struck F, Soutschek E, Deml L, et al. Performance of T-Track® SARS-CoV-2, an Innovative Dual Marker RT-qPCR-Based Whole-Blood Assay for the Detection of SARS-CoV-2-Reactive T Cells. Diagnostics. 2023; 13(17):2722. https://doi.org/10.3390/diagnostics13172722
Chicago/Turabian StyleKanis, Franziska M., Johannes P. Meier, Harald Guldan, Hans-Helmut Niller, Michael Dahm, Alexander Dansard, Thomas Zander, Friedhelm Struck, Erwin Soutschek, Ludwig Deml, and et al. 2023. "Performance of T-Track® SARS-CoV-2, an Innovative Dual Marker RT-qPCR-Based Whole-Blood Assay for the Detection of SARS-CoV-2-Reactive T Cells" Diagnostics 13, no. 17: 2722. https://doi.org/10.3390/diagnostics13172722
APA StyleKanis, F. M., Meier, J. P., Guldan, H., Niller, H.-H., Dahm, M., Dansard, A., Zander, T., Struck, F., Soutschek, E., Deml, L., Möbus, S., & Barabas, S. (2023). Performance of T-Track® SARS-CoV-2, an Innovative Dual Marker RT-qPCR-Based Whole-Blood Assay for the Detection of SARS-CoV-2-Reactive T Cells. Diagnostics, 13(17), 2722. https://doi.org/10.3390/diagnostics13172722