Emerging Approaches for Fluorescence-Based Newborn Screening of Mucopolysaccharidoses

Abstract
1. Introduction
2. Overview of Newborn Screening for MPS Disorders
3. Historical Overview of Laboratory Methods for Enzyme Activity Measurement
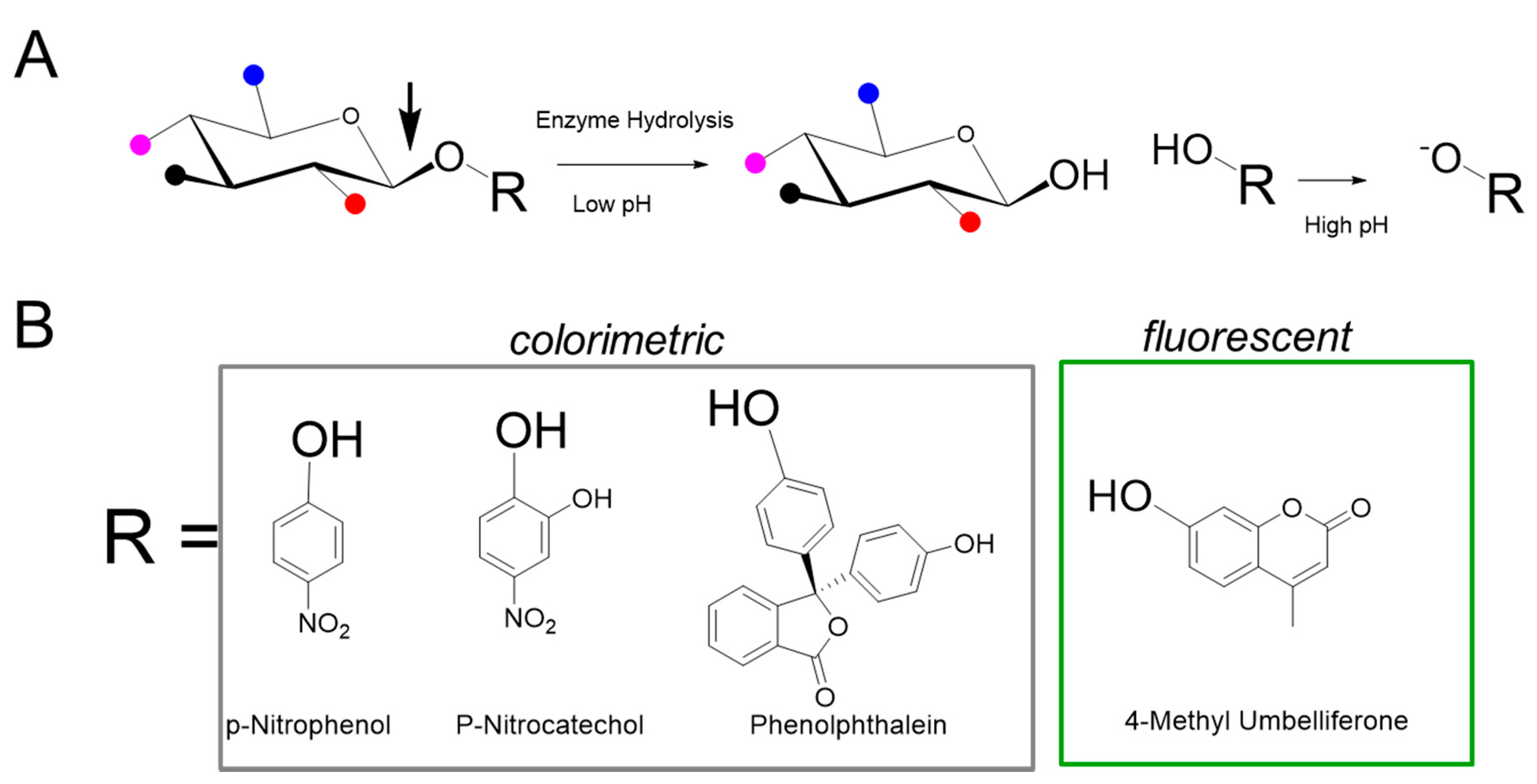
4. Fluorescent Enzyme Activity Measurement
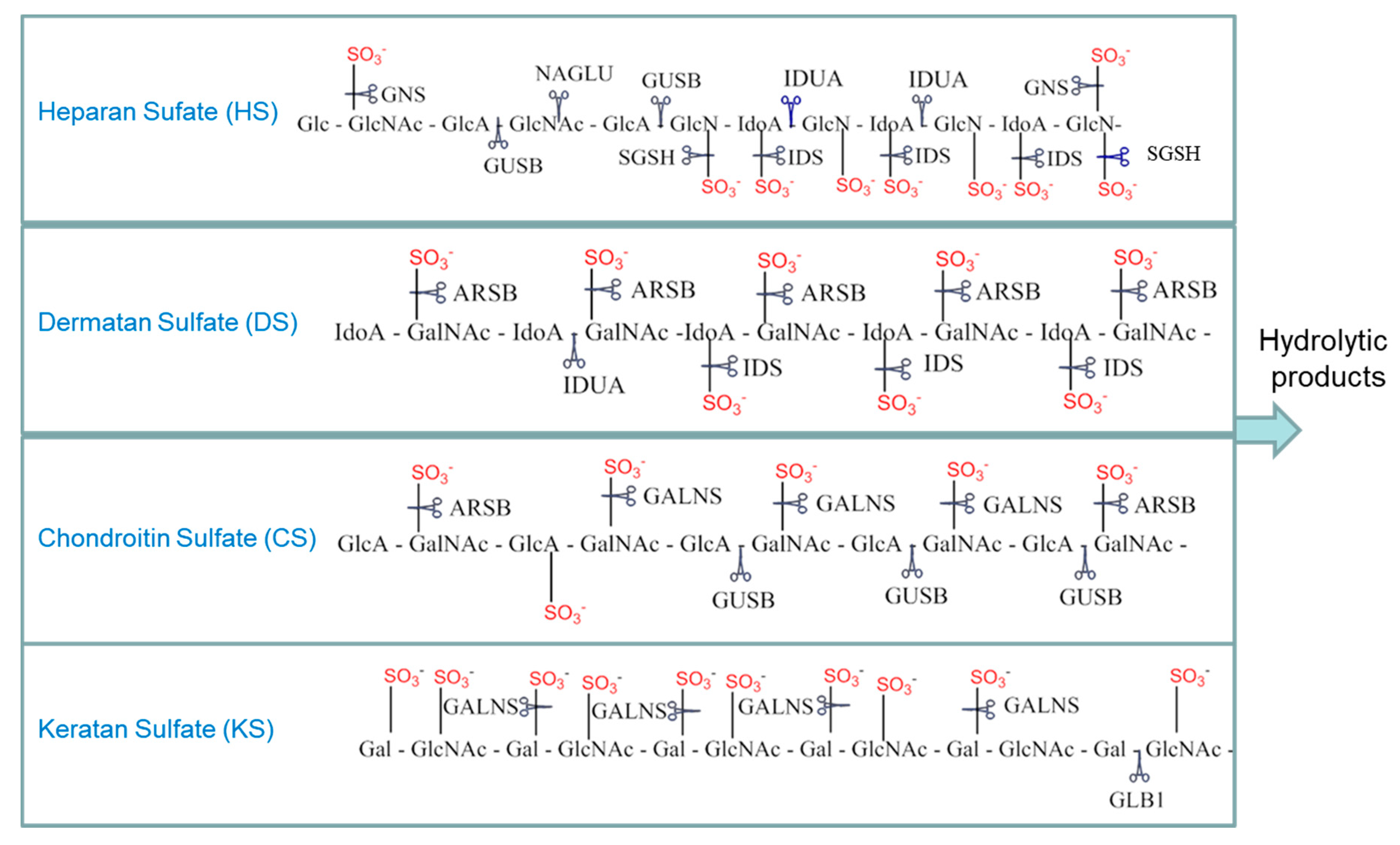
5. Fluorescent Assays for MPSs
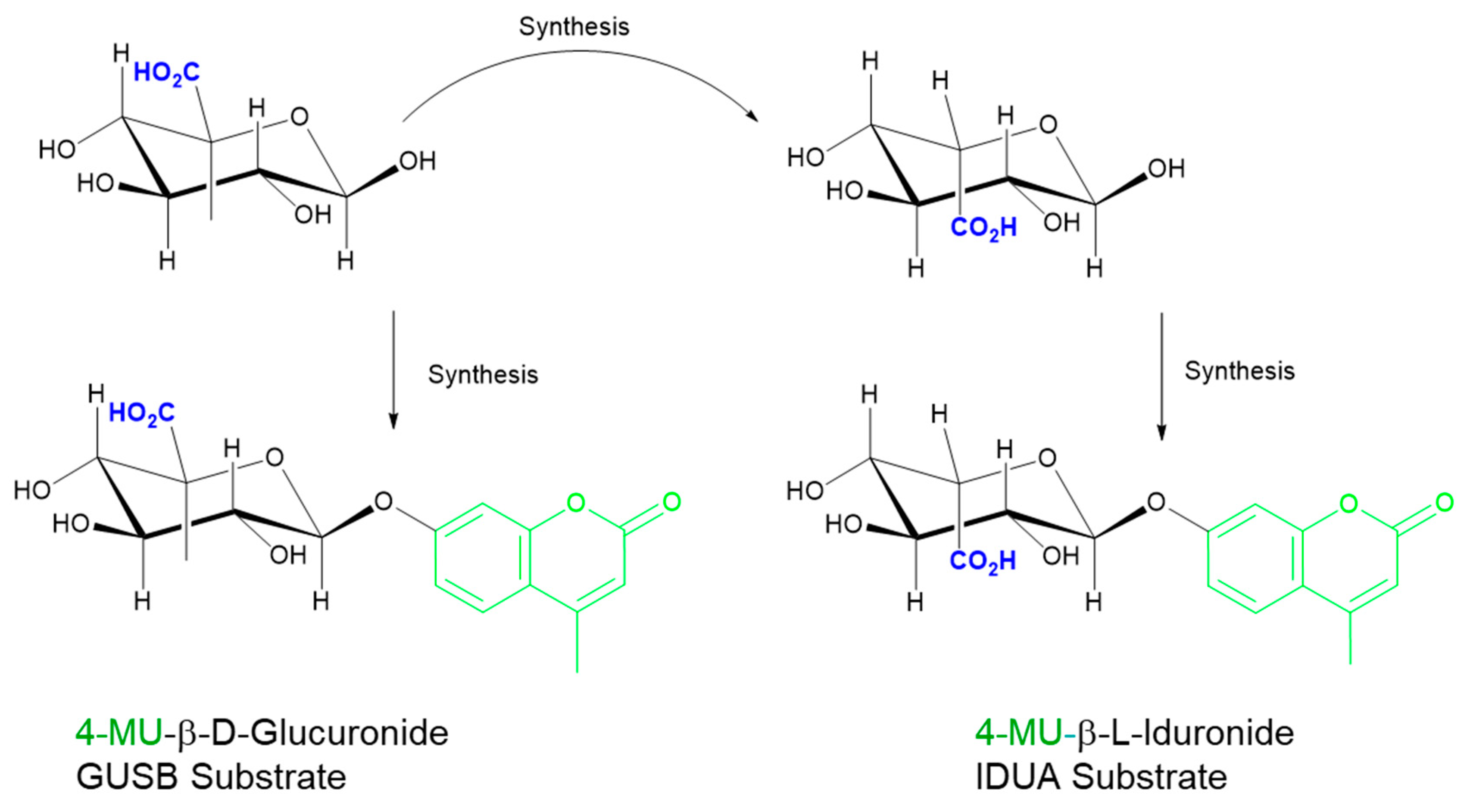
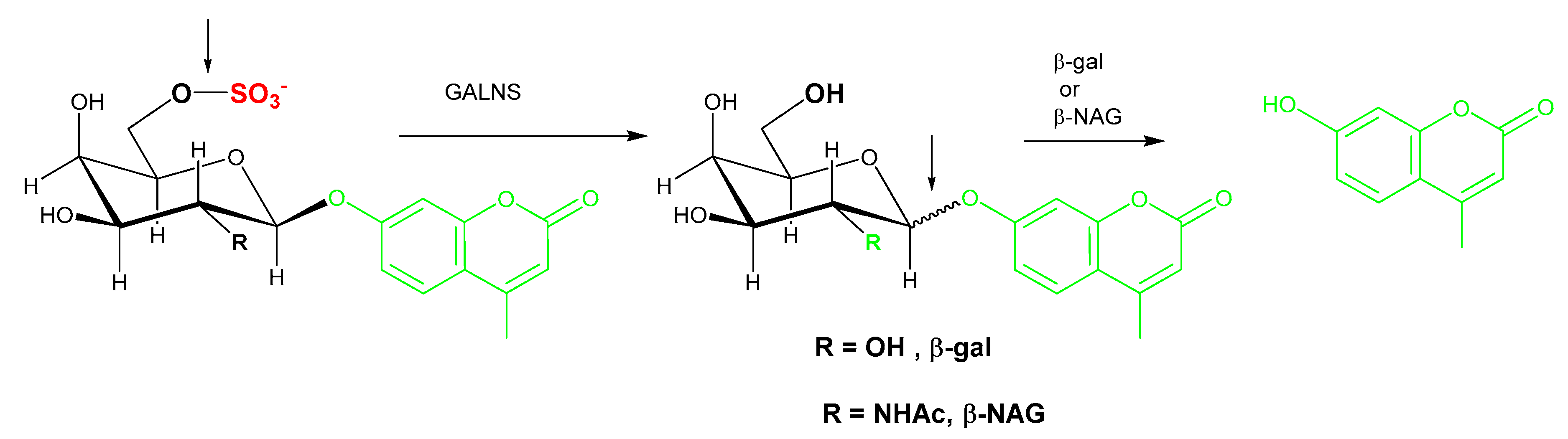
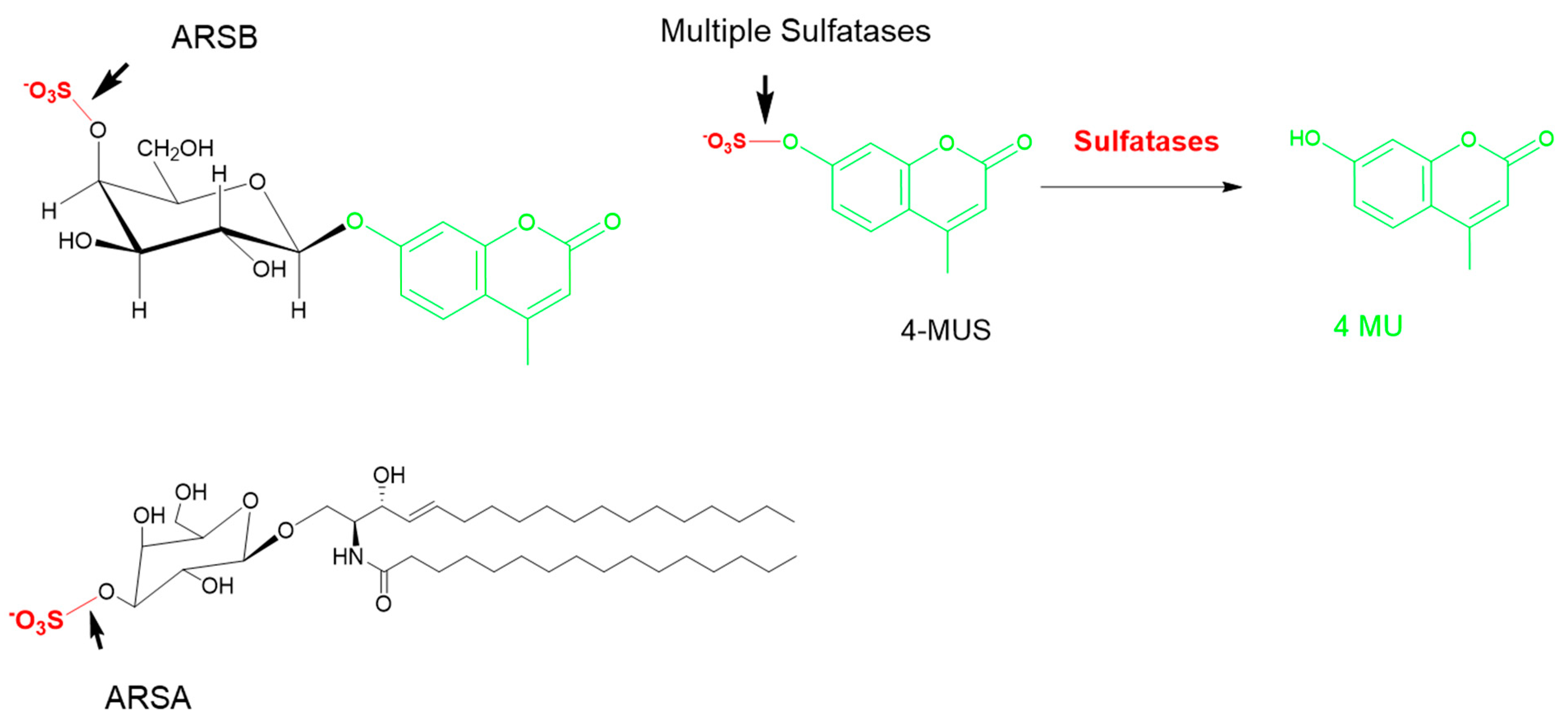
5.1. One-Step Hydrolysis Assays
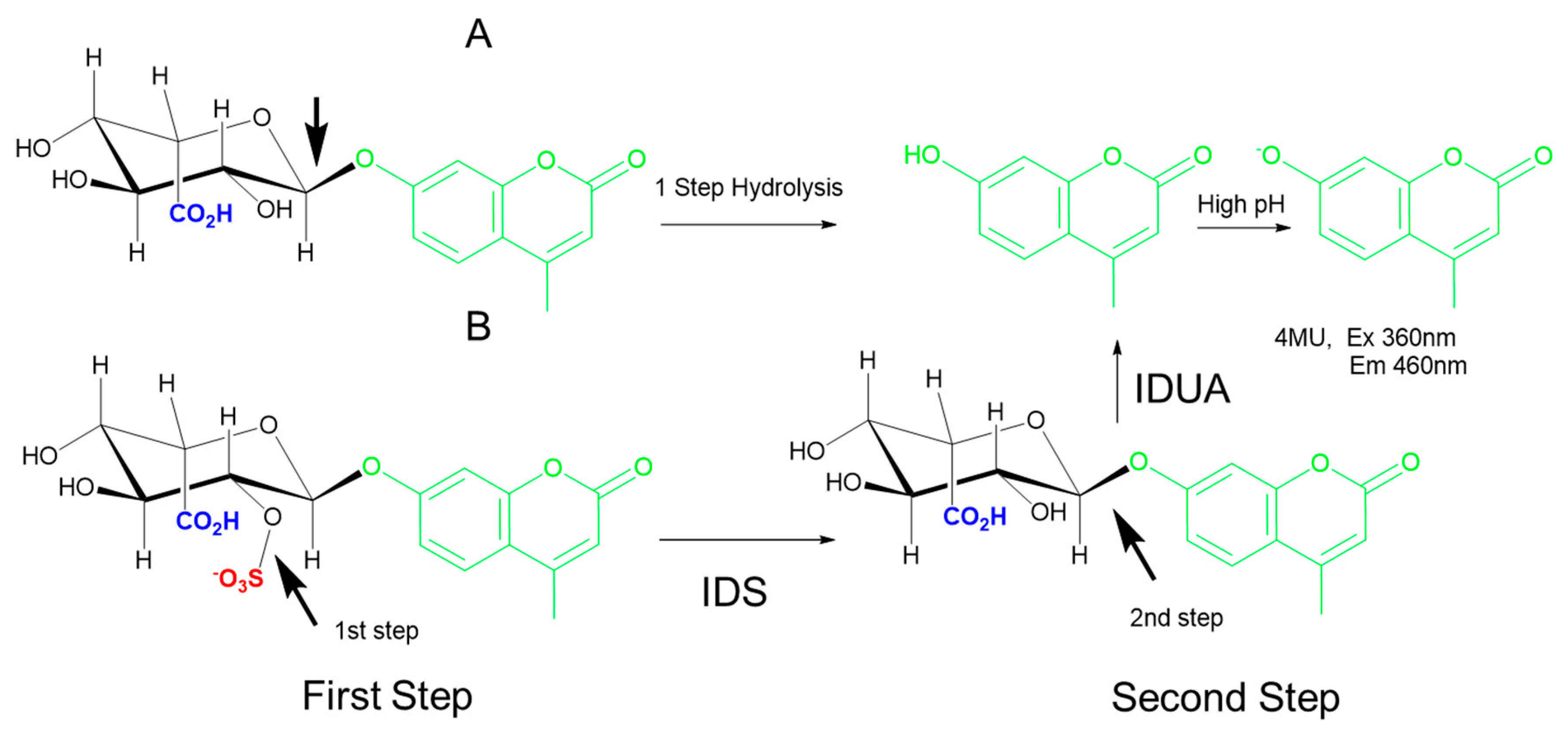
5.2. Two-Step/Sequential Reactions
5.3. Current Challenges in MPS NBS Assays
5.3.1. Matrix Challenges and Interference
5.3.2. Substrate Specificity
5.3.3. Multiplexing Two or More Enzymes
5.3.4. Slow Enzymes
5.4. MPS Newborn Screening Using GAGs
6. Other Considerations for NBS of MPS
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 4-MU | 4-methylumbelliferone |
| DMF | digital microfluidics |
| DMF-F | digital microfluidics fluorescence |
| NBS | newborn screening |
| LSD | lysosomal storage disorder |
| GAG | glycosaminoglycan |
| DBS | dried blood spot |
| MPS | mucopolysaccharidosis |
| IDUA | α-iduronidase |
| IDS | α-iduronide sulfatase |
| SGSH | N-sulfoglucosamine sulfohydrolase |
| NAGLU | acetyl α-glucosaminidase |
| HGSNAT | α-glucosaminide N-acetyl transferase |
| GNS | N-acetyl glucosamine -6-sulfatase |
| GALNS | N-acetyl galactosamine -6-sulfatase |
| GUSB | β-glucuronidase |
| β-gal | β-galactosidase |
| GLB1 | β-glucuronidase |
| ARSB | N-acetyl galactosamine 4-sulfatase |
| ARSA | arylsulfatase A |
| Ido | iduronate |
| CS | chondroitin sulfate |
| DS | dermatan sulfate |
| HS | heparan sulfate |
| KS | keratin sulfate |
| MS/MS | tandem mass spectrometry |
| LC–MS/MS | liquid chromatography–tandem mass spectrometry |
References
- Chuang, C.K.; Lin, H.Y.; Wang, T.J.; Huang, Y.H.; Chan, M.J.; Liao, H.C.; Lo, Y.T.; Wang, L.Y.; Tu, R.Y.; Fang, Y.Y.; et al. Status of newborn screening and follow up investigations for mucopolysaccharidoses I and II in Taiwan. Orphanet J. Rare Dis. 2018, 13, 1–14. [Google Scholar] [CrossRef]
- Tomatsu, S.; Fujii, T.; Fukushi, M.; Oguma, T.; Shimada, T.; Maeda, M.; Kida, K.; Shibata, Y.; Futatsumori, H.; Montaño, A.M.; et al. Newborn screening and diagnosis of mucopolysaccharidoses. Mol. Genet. Metab. 2013, 110, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Muenzer, J. Overview of the mucopolysaccharidoses. Rheumatology 2011, 50, v4–v12. [Google Scholar] [CrossRef] [PubMed]
- Wraith, J.E. Mucopolysaccharidoses and mucolipidoses. In Handbook of Clinical Neurology; Elsevier B.V.: Amsterdam, The Netherlands, 2013; Volume 113, pp. 1723–1729. [Google Scholar]
- Pennock, C.A.; Barnes, I.C. The mucopolysaccharidoses. J. Med. Genet. 1976, 13, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Natowicz, M.R.; Short, M.P.; Wang, Y.; Dickersin, G.R.; Gebhardt, M.C.; Rosenthal, D.I.; Sims, K.B.; Rosenberg, A.E. Brief report: Clinical and biochemical manifestations of hyaluronidase deficiency. N. Engl. J. Med. 1996, 335, 1029–1033. [Google Scholar] [CrossRef]
- Almannai, M.; Marom, R.; Sutton, V.R. Newborn screening: A review of history, recent advancements, and future perspectives in the era of next generation sequencing. Curr. Opin. Pediatr. 2016, 28, 694–699. [Google Scholar] [CrossRef]
- Guthrie, R.; Susi, A. A simple phenylalanine method for detecting phenylketonuria in large populations of newborn infants. Pediatrics 1963, 32, 338–343. [Google Scholar]
- Adam, B.W.; Hall, E.M.; Sternberg, M.; Lim, T.H.; Flores, S.R.; O’Brien, S.; Simms, D.; Li, L.X.; De Jesus, V.R.; Hannon, W.H. The stability of markers in dried-blood spots for recommended newborn screening disorders in the United States. Clin. Biochem. 2011, 44, 1445–1450. [Google Scholar] [CrossRef]
- Millington, D.S. The role of technology in newborn screening. N. C. Med. J. 2019, 80, 49–53. [Google Scholar] [CrossRef]
- Millington, D.S.; Kodo, N.; Norwood, D.L.; Roe, C.R. Tandem mass spectrometry: A new method for acylcarnitine profiling with potential for neonatal screening for inborn errors of metabolism. J. Inherit. Metab. Dis. 1990, 13, 321–324. [Google Scholar] [CrossRef]
- Discretionary Advisory Committee on Heritable Disorders in Newborns and Children Nomination for Newborn Screening of Mucopolysaccharidosis I. Available online: https://www.hrsa.gov/sites/default/files/hrsa/advisory-committees/heritable-disorders/reports-recommendations/letter-to-sec-mps1.pdf (accessed on 29 October 2019).
- Newborn Screening Status for All Disorders | NewSTEPs. Available online: https://www.newsteps.org/resources/newborn-screening-status-all-disorders (accessed on 29 October 2019).
- Klug, T.; Bilyeu, H. Validation and Implementation of MPS II Newborn Screening in Missouri Using a Fluorimetric Assay. Available online: https://www.aphl.org/conferences/NBS-Symposiums/Documents/Poster-abstracts-web-2-26.pdf (accessed on 29 October 2019).
- Beck, M. Treatment strategies for lysosomal storage disorders. Dev. Med. Child Neurol. 2018, 60, 13–18. [Google Scholar] [CrossRef]
- Li, M. Enzyme replacement therapy: A review and its role in treating lysosomal storage siseases. Pediatr. Ann. 2018, 47, e191–e197. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yi, F.; Kumar, A.B.; Kumar Chennamaneni, N.; Hong, X.; Scott, C.R.; Gelb, M.H.; Turecek, F. Multiplex Tandem Mass Spectrometry Enzymatic Activity Assay for Newborn Screening of the Mucopolysaccharidoses and Type 2 Neuronal Ceroid Lipofuscinosis. Clin. Chem. 2017, 63, 1118–1126. [Google Scholar] [CrossRef] [PubMed]
- Chennamaneni, N.K.; Kumar, A.B.; Barcenas, M.; Spáčil, Z.; Scott, C.R.; Tureček, F.; Gelb, M.H. Improved reagents for newborn screening of mucopolysaccharidosis types I, II, and VI by tandem mass spectrometry. Anal. Chem. 2014, 86, 4508–4514. [Google Scholar] [CrossRef] [PubMed]
- Van Hoof, F.; Hers, H.G. The abnormalities of lysosomal enzymes in mucopolysacc- haridoses. Eur. J. Biochem. 1968, 7, 34–44. [Google Scholar] [CrossRef]
- Baum, H.; Dodgson, K.S.; Spencer, B. The assay of arylsulphatases A and B in human urine. Clin. Chim. Acta 1959, 4, 453–455. [Google Scholar] [CrossRef]
- Guilbault, G.G.; Kramer, D.N. Fluorometric determination of lipase, acylase, alpha-, and gamma-chymotrypsin and inhibitors of these enzymes. Anal. Chem. 1964, 36, 409–412. [Google Scholar] [CrossRef]
- Sherman, W.R.; Robins, E. Fluorescence of substituted 7-hydroxycoumarins. Anal. Chem. 1968, 40, 803–805. [Google Scholar] [CrossRef]
- Drake, C.R.; Miller, D.C.; Jones, E.F. Activatable optical probes for the detection of enzymes. Curr. Org. Synth. 2011, 8, 498–520. [Google Scholar] [CrossRef]
- Ray, D.; Bharadwaj, P.K. A coumarin-derived fluorescence probe selective for magnesium. Inorg. Chem. 2008, 47, 2252–2254. [Google Scholar] [CrossRef]
- Li, J.; Zhang, C.F.; Ming, Z.Z.; Hao, G.F.; Yang, W.C.; Yang, G.F. Coumarin-based novel fluorescent zinc ion probe in aqueous solution. Tetrahedron 2013, 69, 4743–4748. [Google Scholar] [CrossRef]
- Mead, J.A.; Smith, J.N.; Williams, R.T. Studies in detoxication. 67. The biosynthesis of the glucuronides of the glucuronides of umbelliferone and 4-methylumbelliferone and their use in fluorimetric determination of b-glucuronidase. Biochem. J. 1955, 61, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D. Fluorimetric determination of glycosidases in the locust (Locusta migratoria) and other insects. Comp. Biochem. Physiol. 1964, 12, 95–105. [Google Scholar] [CrossRef]
- Robinson, D. The fluorimetric determination of beta-glucosidase: Its occurrence in the tissues of animals, including insects. Biochem. J. 1956, 63, 39–44. [Google Scholar]
- Sista, R.; Eckhardt, A.E.; Wang, T.; Séllos-Moura, M.; Pamula, V.K. Rapid, single-step assay for Hunter syndrome in dried blood spots using digital microfluidics. Clin. Chim. Acta 2011, 412, 1895–1897. [Google Scholar] [CrossRef]
- Stirling, J.L.; Robinson, D.; Fensom, A.H.; Benson, P.F.; Baker, J.E. Fluorimetric assay for prenatal detection of Hurler and Scheie homozygoes or heterozygotes. Lancet 1978, 311, 147. [Google Scholar] [CrossRef]
- Hopwood, J.J.; Muller, V.; Smithson, A.; Baggett, N. A fluorometric assay using 4-methylumbelliferyl α-l-iduronide for the estimation of α-l-iduronidase activity and the detection of Hurler and Scheie syndromes. Clin. Chim. Acta 1979, 92, 257–265. [Google Scholar] [CrossRef]
- Minami, R.; Suzuki, M.; Kudoh, T.; Sato, S.; Oyanagi, K. alpha-L-Iduronidase activity in established lymphoblastoid cells from patients with Hurler and Scheie syndromes transformed by Epstein-Barr virus. Tohoku J. Exp. Med. 1977, 122, 393–396. [Google Scholar] [CrossRef]
- Chan, M.J.; Liao, H.C.; Gelb, M.H.; Chuang, C.K.; Liu, M.Y.; Chen, H.J.; Kao, S.M.; Lin, H.Y.; Huang, Y.H.; Kumar, A.B.; et al. Taiwan national newborn screening program by tandem mass spectrometry for mucopolysaccharidoses types I, II, and VI. J. Pediatr. 2019, 205, 176–182. [Google Scholar] [CrossRef]
- Lee, K.; Jun, S.H.; Song, S.H.; Park, H.D.; Park, K.U.; Song, J. Direct assay of iduronate-2-sulfatase for Hunter disease using UPLC-tandem mass spectrometry and fluorogenic substrate. Clin. Biochem. 2015, 48, 1350–1353. [Google Scholar] [CrossRef]
- Chamoles, N.A.; Blanco, M.; Gaggioli, D. Diagnosis of α-L-iduronidase deficiency in dried blood spots on filter paper: The possibility of newborn diagnosis. Clin. Chem. 2001, 47, 780–781. [Google Scholar] [CrossRef]
- Chamoles, N.A.; Blanco, M.; Gaggioli, D.; Casentini, C. Gaucher and Niemann-Pick diseases--enzymatic diagnosis in dried blood spots on filter paper: Retrospective diagnoses in newborn-screening cards. Clin. Chim. Acta. 2002, 317, 191–197. [Google Scholar] [CrossRef]
- Chamoles, N.A.; Niizawa, G.; Blanco, M.; Gaggioli, D.; Casentini, C. Glycogen storage disease type II: Enzymatic screening in dried blood spots on filter paper. Clin. Chim. Acta 2004, 347, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-P.; Lin, H.-Y.; Wang, T.-J.; Chang, C.-Y.; Lin, C.-H.; Huang, S.-F.; Tsai, C.-C.; Liu, H.-L.; Keutzer, J.; Chuang, C.-K. A pilot newborn screening program for Mucopolysaccharidosis type I in Taiwan. Orphanet J. Rare Dis. 2013, 8, 147. [Google Scholar] [CrossRef] [PubMed]
- Sista, R.S.; Wang, T.; Wu, N.; Graham, C.; Eckhardt, A.; Winger, T.; Srinivasan, V.; Bali, D.; Millington, D.S.; Pamula, V.K. Multiplex newborn screening for Pompe, Fabry, Hunter, Gaucher, and Hurler diseases using a digital microfluidic platform. Clin. Chim. Acta 2013, 424, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Sista, R.S.; Eckhardt, A.E.; Wang, T.; Graham, C.; Rouse, J.L.; Norton, S.M.; Srinivasan, V.; Pollack, M.G.; Tolun, A.A.; Bali, D.; et al. Digital microfluidic platform for multiplexing enzyme assays: Implications for lysosomal storage disease screening in newborns. Clin. Chem. 2011, 57, 1444–1451. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration DEN150035 Evaluation of Automatic Class III Designation for SEEKER System DECISION SUMMARY Quantitative Fluorimetric Enzymatic Activity Assay. Available online: https://www.accessdata.fda.gov/cdrh_docs/reviews/DEN150035.pdf (accessed on 29 October 2019).
- Fink, D.W.; Koehler, W.R. pH effects on fluorescence of umbelliferone. Anal. Chem. 1970, 42, 990–993. [Google Scholar] [CrossRef]
- Zhi, H.; Wang, J.; Wang, S.; Wei, Y. Fluorescent properties of hymecromone and fluorimetric analysis of hymecromone in compound dantong capsule. J. Spectrosc. 2013. [Google Scholar] [CrossRef]
- Voznyi, Y.V.; Karpova, E.A.; Dudukina, T.V.; Tsvetkova, I.V.; Boer, A.M.; Janse, H.C.; van Diggelen, O.P. A fluorimetric enzyme assay for the diagnosis of Sanfilippo disease C (MPS III C). J. Inherit. Metab. Dis. 1993, 16, 465–472. [Google Scholar] [CrossRef]
- Wood, T.C.; Harvey, K.; Beck, M.; Burin, M.G.; Chien, Y.H.; Church, H.J.; D’Almeida, V.; Van Diggelen, O.P.; Fietz, M.; Giugliani, R.; et al. Diagnosing mucopolysaccharidosis IVA. J. Inherit. Metab. Dis. 2013, 36, 293–307. [Google Scholar] [CrossRef]
- Cé, J.; Rodrigues, M.T.; Käfer, E.T.; da Costa Moraes, V.; Coelho, J.C. Beta-glucuronidase activity in dried blood spots: Reduced technique with biochemical parameters determined. Clin. Biochem. 2017, 50, 1243–1248. [Google Scholar] [CrossRef] [PubMed]
- Uribe, A.; Ayala, A.; España, M.; Arevalo, I.; Pacheco, N.; Garcia, L.M.J. b-Galactosidase Deficiency in Colombia: Report of 20 Patients Detected Using Dried Blood Spot Samples. J. Inborn Errors Metab. Screen. 2015, 3. [Google Scholar] [CrossRef]
- Regier, D.S.; Tifft, C.J. GLB1-Related Disorders; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Singh, R.; Chopra, S.; Norton, S.; Pamula, V. Demonstration of a Digital Microfluidic Platform for the High Throughput Analysis of 12 Discrete Fluorimetric Enzyme Assays Using a Single Newborn Dried Blood Spot Punch. Available online: https://worldsymposia.org/wp-content/uploads/WORLDSymposium-2018-Poster-List.pdf (accessed on 9 May 2020).
- Voznyi, Y.V.; Keulemans, J.L.M.; Van Diggelen, O.P. A fluorimetric enzyme assay for the diagnosis of MPS II (hunter disease). J. Inherit. Metab. Dis. 2001, 24, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Tolun, A.A.; Graham, C.; Shi, Q.; Sista, R.S.; Wang, T.; Eckhardt, A.E.; Pamula, V.K.; Millington, D.S.; Bali, D.S. A novel fluorometric enzyme analysis method for Hunter syndrome using dried blood spots. Mol. Genet. Metab. 2012, 105, 519–521. [Google Scholar] [CrossRef]
- Civallero, G.; Michelin, K.; de Mari, J.; Viapiana, M.; Burin, M.; Coelho, J.C.; Giugliani, R. Twelve different enzyme assays on dried-blood filter paper samples for detection of patients with selected inherited lysosomal storage diseases. Clin. Chim. Acta. 2006, 372, 98–102. [Google Scholar] [CrossRef]
- Kresse, H.; Paschke, E.; von Figura, K.; Gilberg, W.; Fuchs, W. Sanfilippo disease type D: Deficiency of N-acetylglucosamine-6-sulfate sulfatase required for heparan sulfate degradation. Proc. Natl. Acad. Sci. USA 1980, 77, 6822–6826. [Google Scholar] [CrossRef]
- Freeman, C.; Hopwood, J. Lysosomal degradation of heparin and heparan sulphate. Adv. Exp. Med. Biol. 1992, 313, 121–134. [Google Scholar]
- He, W.; Voznyi, Y.V.; Boer, A.M.; Kleijer, W.J.; van Diggelen, O.P. A fluorimetric enzyme assay for the diagnosis of Sanfilippo disease type D (MPS IIID). J. Inherit. Metab. Dis. 1993, 16, 935–941. [Google Scholar] [CrossRef]
- Van Diggelen, O.P.; Zhao, H.; Kleijer, W.J.; Janse, H.C.; Poorthuis, B.J.H.M.; van Pelt, J.; Kamerling, J.P.; Galjaard, H. A fluorimetric enzyme assay for the diagnosis of Morquio disease type A (MPS IV A). Clin. Chim. Acta 1990, 187, 131–139. [Google Scholar] [CrossRef]
- Camelier, M.V.; Burin, M.G.; De Mari, J.; Vieira, T.A.; Marasca, G.; Giugliani, R. Practical and reliable enzyme test for the detection of Mucopolysaccharidosis IVA (Morquio Syndrome type A) in dried blood samples. Clin. Chim. Acta 2011, 412, 1805–1808. [Google Scholar] [CrossRef]
- Karpova, E.A.; Voznyi, Y.A.; Keulemans, J.L.M.; Hoogeveen, A.T.; Winchester, B.; Tsvetkova, I.V.; Van Diggelen, O.P. A fluorimetric enzyme assay for the diagnosis of Sanfilippo disease type A (MPS IIIA). J. Inherit. Metab. Dis. 1996, 19, 278–285. [Google Scholar] [CrossRef]
- Ruzo, A.; Marcó, S.; García, M.; Villacampa, P.; Ribera, A.; Ayuso, E.; Maggioni, L.; Mingozzi, F.; Haurigot, V.; Bosch, F. Correction of pathological accumulation of glycosaminoglycans in central nervous system and peripheral tissues of MPSIIIA mice through systemic AAV9 gene transfer. Hum. Gene Ther. 2012, 23, 1237–1246. [Google Scholar] [CrossRef] [PubMed]
- Fraldi, A.; Hemsley, K.; Crawley, A.; Lombardi, A.; Lau, A.; Sutherland, L.; Auricchio, A.; Ballabio, A.; Hopwood, J.J. Functional correction of CNS lesions in an MPS-IIIA mouse model by intracerebral AAV-mediated delivery of sulfamidase and SUMF1 genes. Hum. Mol. Genet. 2007, 16, 2693–2702. [Google Scholar] [CrossRef] [PubMed]
- Langford-Smith, A.; Wilkinson, F.L.; Langford-Smith, K.J.; Holley, R.J.; Sergijenko, A.; Howe, S.J.; Bennett, W.R.; Jones, S.A.; Wraith, J.E.; Merry, C.L.R.; et al. Hematopoietic stem cell and gene therapy corrects primary neuropathology and behavior in mucopolysaccharidosis IIIA mice. Mol. Ther. 2012, 20, 1610–1621. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, B.J.; Ghomashchi, F.; Kim, T.; Abam, C.A.; Sadilek, M.; Jack, R.; Thompson, J.N.; Scott, C.R.; Gelb, M.H.; Turecek, F. New substrates and enzyme Assays for the detection of mucopolysaccharidosis III (Sanfilippo Syndrome) Types A, B, C, and D by tandem mass spectrometry. Bioconjug. Chem. 2012, 23, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Yi, F.; Gelb, M.H.; Zalatan, J.; Nelson, A. Development of Newborn Screening Methods for Mucopolysaccharidosis III Type A and Type B in Dried Blood Spot Using Tandem Mass Spectrometry. Ph.D. Thesis, University of Washington, Seattle, WA, USA, 2018. [Google Scholar]
- Choi, Y.; Tuzikov, A.B.; Ovchinnikova, T.V.; Bovin, N.V.; Pshezhetsky, A.V. Novel Direct Assay for Acetyl-CoA:α-Glucosaminide N-Acetyltransferase Using BODIPY-Glucosamine as a Substrate. JIMD Rep. 2016, 28, 11–18. [Google Scholar]
- Briciu-Burghina, C.; Heery, B.; Regan, F. Continuous fluorometric method for measuring β-glucuronidase activity: Comparative analysis of three fluorogenic substrates. Analyst 2015, 140, 5953–5964. [Google Scholar] [CrossRef]
- Kumar, A.B.; Masi, S.; Ghomashchi, F.; Chennamaneni, N.K.; Ito, M.; Scott, C.R.; Turecek, F.; Gelb, M.H.; Spacil, Z. Tandem mass spectrometry has a larger analytical range than fluorescence assays of lysosomal enzymes: Application to newborn screening and diagnosis of mucopolysaccharidoses types II, IVA, and VI. Clin. Chem. 2015, 61, 1363–1371. [Google Scholar] [CrossRef]
- Oemardien, L.F.; Boer, A.M.; Ruijter, G.J.G.; van der Ploeg, A.T.; de Klerk, J.B.C.; Reuser, A.J.J.; Verheijen, F.W. Hemoglobin precipitation greatly improves 4-methylumbelliferone-based diagnostic assays for lysosomal storage diseases in dried blood spots. Mol. Genet. Metab. 2011, 102, 44–48. [Google Scholar] [CrossRef]
- Franco, P.G.; Pérez, M.J.; Aranda, C.; Adamo, A.; Silvestroff, L. Improving arylsulfatase activity determination in dried blood spots: Screening and diagnostic approaches for Maroteaux-Lamy syndrome (MPS VI). Clin. Chim. Acta 2015, 446, 86–92. [Google Scholar] [CrossRef]
- Chang, P.L.; Rosa, N.E.; Davidson, R.G. Differential assay of arylsulfatase A and B activities: A sensitive method for cultured human cells. Anal. Biochem. 1981, 117, 382–389. [Google Scholar] [CrossRef]
- Delvin, E.E.; Pottier, A.; Glorieux, F.H. Comparative activity of arylsulphatases A and B on two synthetic substrates. Biochem. J. 1976, 157, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Ullal, A.J.; Millington, D.S.; Bali, D.S. Development of a fluorometric microtiter plate-based enzyme assay for arylsulfatase B (MPS VI) using dried blood spots. Mol. Genet. Metab. Reports 2014, 1, 465–467. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, A.; Sokolsky, T.; Wyman, S.K.; Reese, M.G.; Puffenberger, E.; Strauss, K.; Morton, H.; Parad, R.B.; Naylor, E.W. Development of DNA confirmatory and high-risk diagnostic testing for newborns using targeted next-generation DNA sequencing. Genet. Med. 2015, 17, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Burlina, A.B.; Polo, G.; Rubert, L.; Gueraldi, D.; Cazzorla, C.; Duro, G.; Salviati, L.; Burlina, A.P. Implementation of second-tier tests in newborn screening for lysosomal disorders in North Eastern Italy. Int. J. Neonatal Screen. 2019, 5, 24. [Google Scholar] [CrossRef]
- Duffey, T.A.; Bellamy, G.; Elliott, S.; Fox, A.C.; Glass, M.; Turecek, F.; Gelb, M.H.; Scott, C.R. A tandem mass spectrometry triplex assay for the detection of fabry, pompe, and mucopolysaccharidosis-I (Hurler). Clin. Chem. 2010, 56, 1854–1861. [Google Scholar] [CrossRef]
- Spacil, Z.; Tatipaka, H.; Barcenas, M.; Scott, C.R.; Turecek, F.; Gelb, M.H. High-throughput assay of 9 lysosomal enzymes for newborn screening. Clin. Chem. 2013, 59, 502–511. [Google Scholar] [CrossRef]
- Gelb, M.H.; Lukacs, Z.; Ranieri, E.; Schielen, P.C.J.I. Newborn screening for lysosomal storage disorders: Methodologies for measurement of enzymatic activities in dried blood spots. Int. J. Neonatal Screen. 2019, 5, 1. [Google Scholar] [CrossRef]
- Burton, B.K.; Hoganson, G.E.; Grange, D.K.; Braddock, S.R.; Christensen, K.M.; Hitchins, L.; Hickey, R.; Shao, R.; Basheeruddin, K.; Basheeruddin, K. Newborn screening for mucopolysaccharidosis type II (MPS II) in Illinois: The first year’s experience. Mol. Genet. Metab. 2019, 126, S34. [Google Scholar] [CrossRef]
- FLEXMAP 3D® System | Advanced and Versatile Multiplexing Platform. Available online: https://www.luminexcorp.com/flexmap-3d/ (accessed on 27 January 2020).
- Motabar, O.; Shi, Z.D.; Goldin, E.; Liu, K.; Southall, N.; Sidransky, E.; Austin, C.P.; Griffiths, G.L.; Zheng, W. A new resorufin-based α-glucosidase assay for high-throughput screening. Anal. Biochem. 2009, 390, 79–84. [Google Scholar] [CrossRef][Green Version]
- Ou, L.; Herzog, T.L.; Wilmot, C.M.; Whitley, C.B. Standardization of α-L-iduronidase enzyme assay with Michaelis-Menten kinetics. Mol. Genet. Metab. 2014, 111, 113–115. [Google Scholar] [CrossRef] [PubMed]
- Whyte, L.S.; Hopwood, J.J.; Hemsley, K.M.; Lau, A.A. Variables influencing fluorimetric N-sulfoglucosamine sulfohydrolase (SGSH) activity measurement in brain homogenates. Mol. Genet. Metab. Reports 2015, 5, 60–62. [Google Scholar] [CrossRef] [PubMed]
- Gray, G.; Claridge, P.; Jenkinson, L.; Green, A. Quantitation of urinary glycosaminoglycans using dimethylene blue as a screening technique for the diagnosis of mucopolysaccharidoses: An evaluation. Ann. Clin. Biochem. 2007, 44, 360–363. [Google Scholar] [CrossRef]
- Kubaski, F.; Mason, R.W.; Nakatomi, A.; Shintaku, H.; Xie, L.; van Vlies, N.N.; Church, H.; Giugliani, R.; Kobayashi, H.; Yamaguchi, S.; et al. Newborn screening for mucopolysaccharidoses: A pilot study of measurement of glycosaminoglycans by tandem mass spectrometry. J. Inherit. Metab. Dis. 2017, 40, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Kubaski, F.; Suzuki, Y.; Orii, K.; Giugliani, R.; Church, H.J.; Mason, R.W.; Dũng, V.C.; Ngoc, C.T.B.; Yamaguchi, S.; Kobayashi, H.; et al. Glycosaminoglycan levels in dried blood spots of patients with mucopolysaccharidoses and mucolipidoses. Mol. Genet. Metab. 2017, 120, 247–254. [Google Scholar] [CrossRef]
- Tomatsu, S.; Montaño, A.M.; Oguma, T.; Dung, V.C.; Oikawa, H.; Gutiérrez, M.L.; Yamaguchi, S.; Suzuki, Y.; Fukushi, M.; Barrera, L.A.; et al. Validation of disaccharide compositions derived from dermatan sulfate and heparan sulfate in mucopolysaccharidoses and mucolipidoses II and III by tandem mass spectrometry. Mol. Genet. Metab. 2010, 99, 124–131. [Google Scholar] [CrossRef]
- Tomatsu, S.; Shimada, T.; Mason, R.W.; Montaño, A.M.; Kelly, J.; LaMarr, W.A.; Kubaski, F.; Giugliani, R.; Guha, A.; Yasuda, E.; et al. Establishment of glycosaminoglycan assays for mucopolysaccharidoses. Metabolites 2014, 4, 655–679. [Google Scholar] [CrossRef]
- Stapleton, M.; Kubaski, F.; Mason, R.W.; Shintaku, H.; Kobayashi, H.; Yamaguchi, S.; Taketani, T.; Suzuki, Y.; Orii, K.; Orii, T.; et al. Newborn screening for mucopolysaccharidoses: Measurement of glycosaminoglycans by LC-MS/MS. Mol. Genet. Metab. Reports 2020, 22, 100563. [Google Scholar] [CrossRef]
- De Ruijter, J.; de Ru, M.H.; Wagemans, T.; Ijlst, L.; Lund, A.M.; Orchard, P.J.; Schaefer, G.B.; Wijburg, F.A.; van Vlies, N. Heparan sulfate and dermatan sulfate derived disaccharides are sensitive markers for newborn screening for mucopolysaccharidoses types I, II and III. Mol. Genet. Metab. 2012, 107, 705–710. [Google Scholar] [CrossRef]
- Arunkumar, N.; Langan, T.J.; Stapleton, M.; Kubaski, F.; Mason, R.W.; Singh, R.; Kobayashi, H.; Yamaguchi, S.; Suzuki, Y.; Orii, K.; et al. Newborn screening of mucopolysaccharidoses: Past, present, and future. J. Hum. Genet. 2020. [Google Scholar] [CrossRef]
- Volpi, N.; Galeotti, F.; Yang, B.; Linhardt, R.J. Analysis of glycosaminoglycan-derived, precolumn, 2-aminoacridone-labeled disaccharides with LC-fluorescence and LC-MS detection. Nat. Protoc. 2014, 9, 541–558. [Google Scholar] [CrossRef] [PubMed]
- Maccari, F.; Galeotti, F.; Mantovani, V.; Zampini, L.; Padella, L.; Rigon, L.; Concolino, D.; Fiumara, A.; Pascale, E.; Pittalà, A.; et al. Composition and structure of glycosaminoglycans in DBS from 2-3-day-old newborns for the diagnosis of mucopolysaccharidosis. Anal. Biochem. 2018, 557, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Warttinger, U.; Giese, C.; Harenberg, J.; Holmer, E.; Krämer, R. A fluorescent probe assay (Heparin Red) for direct detection of heparins in human plasma. Anal. Bioanal. Chem. 2016, 408, 8241–8251. [Google Scholar] [CrossRef] [PubMed]
- Osago, H.; Shibata, T.; Hara, N.; Kuwata, S.; Kono, M.; Uchio, Y.; Tsuchiya, M. Quantitative analysis of glycosaminoglycans, chondroitin/dermatan sulfate, hyaluronic acid, heparan sulfate, and keratan sulfate by LC-ESI-MS/MS. Anal. Biochem. 2014, 467, 62–74. [Google Scholar] [CrossRef]
- Lawrence, R.; Brown, J.R.; Lorey, F.; Dickson, P.I.; Crawford, B.E.; Esko, J.D. Glycan-based biomarkers for mucopolysaccharidoses. Mol. Genet. Metab. 2014, 111, 73–83. [Google Scholar] [CrossRef]
- Alonso-Fernández, J.R.; Fidalgo, J.; Colón, C. Neonatal screening for mucopolysaccharidoses by determination of glycosaminoglycans in the eluate of urine-impregnated paper: Preliminary results of an improved DMB-based procedure. J. Clin. Lab. Anal. 2010, 24, 149–153. [Google Scholar] [CrossRef]
- Clarke, L.A.; Atherton, A.M.; Burton, B.K.; Day-Salvatore, D.L.; Kaplan, P.; Leslie, N.D.; Scott, C.R.; Stockton, D.W.; Thomas, J.A.; Muenzer, J. Mucopolysaccharidosis Type I newborn screening: Best practices for diagnosis and management. J. Pediatr. 2017, 182, 363–370. [Google Scholar] [CrossRef]
- Donati, M.A.; Pasquini, E.; Spada, M.; Polo, G.; Burlina, A. Newborn screening in mucopolysaccharidoses. Ital. J. Pediatr. 2018, 44 (Suppl 2), 25–34. [Google Scholar] [CrossRef]
- Bravo, H.; Neto, E.C.; Schulte, J.; Pereira, J.; Filho, C.S.; Bittencourt, F.; Sebastião, F.; Bender, F.; de Magalhães, A.P.S.; Guidobono, R.; et al. Investigation of newborns with abnormal results in a newborn screening program for four lysosomal storage diseases in Brazil. Mol. Genet. Metab. Reports 2017, 12, 92–97. [Google Scholar] [CrossRef]
- ClinVar. Available online: https://www.ncbi.nlm.nih.gov/clinvar/ (accessed on 3 March 2020).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MPS | OMIM# | Gene | Deficient Enzyme (EC#) | Elevated GAG | Key Disease Features | FDA-Approved Therapies |
|---|---|---|---|---|---|---|
| MPS I (Hurler, Scheie, Hurler/Scheie) | 607014, 607015, 607016 | IDUA | α-iduronidase (3.2.1.76) | DS, HS | Corneal clouding, skeletal abnormalities, organ enlargement, heart disease, mental retardation, death in childhood | Aldurazyme |
| MPS II (Hunter) | 300823 | IDS | α-iduronide sulfatase (3.1.6.13) | DS, HS | Elaprase | |
| MPS IIIA (Sanfilippo A) | 252900 | SGSH | N-sulfoglucosamine sulfohydrolase (3.10.1.1) | HS | Profound mental deterioration, hyperactivity, and mild somatic manifestations | |
| MPS IIIB (Sanfilippo B) | 252920 | NAGLU | Acetyl α-glucosaminidase (3.2.1.50) | HS | ||
| MPS IIIC (Sanfilippo C) | 252930 | HGSNAT | α-glucosaminide N-acetyl transferase (2.3.1.78) | HS | ||
| MPS IIID (Sanfilippo D) | 252940 | GNS | N-acetyl glucosamine-6-sulfatase (3.1.6.14) | HS | ||
| MPS IVA (Morquio A) | 253000 | GALNS | N-acetyl galactosamine -6-sulfatase (3.1.6.4) | KS, CS | Skeletal abnormalities, loose ligaments, degenerative joint disease, corneal clouding, heart disease, death in childhood or young adulthood | Vimzim |
| MPS IVB (Morquio B) | 253010 | GLB1 | β-galactosidase (3.2.1.23) | KS | ||
| MPS VI (Maroteaux-Lamy) | 253200 | ARSB | N-acetyl galactosamine 4-sulfatase (3.1.6.1) | DS, CS | Corneal clouding, skeletal abnormalities, organ enlargement, heart disease, death in childhood | Naglazyme |
| MPS VII (Sly) | 253220 | GUSB | β-glucuronidase (3.2.1.31) | DS, HS, CS | Corneal clouding, skeletal abnormalities, organ enlargement, heart disease, mental retardation, death in childhood | Mepsevii |
| Condition | Fluorescent Substrate | Assay | Sample Types |
|---|---|---|---|
| MPS I | 4MU-α-l-iduronide (4MU-IdoA) | 1 step + pH change | DBS, recombinant (r)IDUA, fibroblasts, leukocytes |
| MPS II | 4MU-α-l-iduronide-2-sulfate (4MU-IdoA2S) | 2 steps + pH change | DBS, rIDS, fibroblasts, leukocytes |
| MPS IIIA | 4MU-N-sulpho-α-d-glucosaminide (4MU-Glc2NS) | 2 steps + pH change | rSGSH, fibroblasts, leukocytes, cell lysates |
| MPS IIIB | 4MU-N-acetyl-α-d-glucosaminide (4MU-GlcNAc) | 1 step + pH change | DBS, rNAGLU, fibroblasts, leukocytes |
| MPS IIIC | 4MU-2-amino-deoxy glucose (4MU-GlcN) | 2 steps + pH change | rHGSNAT, leukocytes, fibroblasts, cell lysates |
| MPS IIID | 4MU-N-acetyl-α-d-glucosaminide-6-sulfate (4MU-GlcNAcS) | 2 steps + pH change | rGNS, fibroblasts, leukocytes |
| MPS IVA | 4MU-N-acetyl-α-d-galactoseaminide-6-sulfate (4MU-GalNAc-6S) | 2 steps + pH change | rGALNS, DBS, leukocytes |
| MPS IVB | 4MU-β-d-galactoside (4MU-Gal) | 1 step + pH change | rβGal, DBS |
| MPS VI | 4MU-N-acetyl-α-d-galactoseaminide-4-sulfate (4MU-GalNAc-4S) | 2 steps + pH change | rARSB, DBS |
| 4MUS | 1 step + pH change | ||
| MPS VII | 4MU-β-d-glucuronide (4MU-GlcA) | 1 step + pH change | rβGlc, DBS |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, R.; Chopra, S.; Graham, C.; Langer, M.; Ng, R.; Ullal, A.J.; Pamula, V.K. Emerging Approaches for Fluorescence-Based Newborn Screening of Mucopolysaccharidoses. Diagnostics 2020, 10, 294. https://doi.org/10.3390/diagnostics10050294
Singh R, Chopra S, Graham C, Langer M, Ng R, Ullal AJ, Pamula VK. Emerging Approaches for Fluorescence-Based Newborn Screening of Mucopolysaccharidoses. Diagnostics. 2020; 10(5):294. https://doi.org/10.3390/diagnostics10050294
Chicago/Turabian StyleSingh, Rajendra, Shaileja Chopra, Carrie Graham, Melissa Langer, Rainer Ng, Anirudh J. Ullal, and Vamsee K. Pamula. 2020. "Emerging Approaches for Fluorescence-Based Newborn Screening of Mucopolysaccharidoses" Diagnostics 10, no. 5: 294. https://doi.org/10.3390/diagnostics10050294
APA StyleSingh, R., Chopra, S., Graham, C., Langer, M., Ng, R., Ullal, A. J., & Pamula, V. K. (2020). Emerging Approaches for Fluorescence-Based Newborn Screening of Mucopolysaccharidoses. Diagnostics, 10(5), 294. https://doi.org/10.3390/diagnostics10050294