
Bending Elasticity Modulus of Giant Vesicles Composed of Aeropyrum Pernix K1 Archaeal Lipid






Abstract
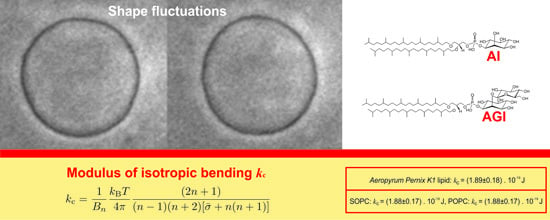
:
1. Introduction
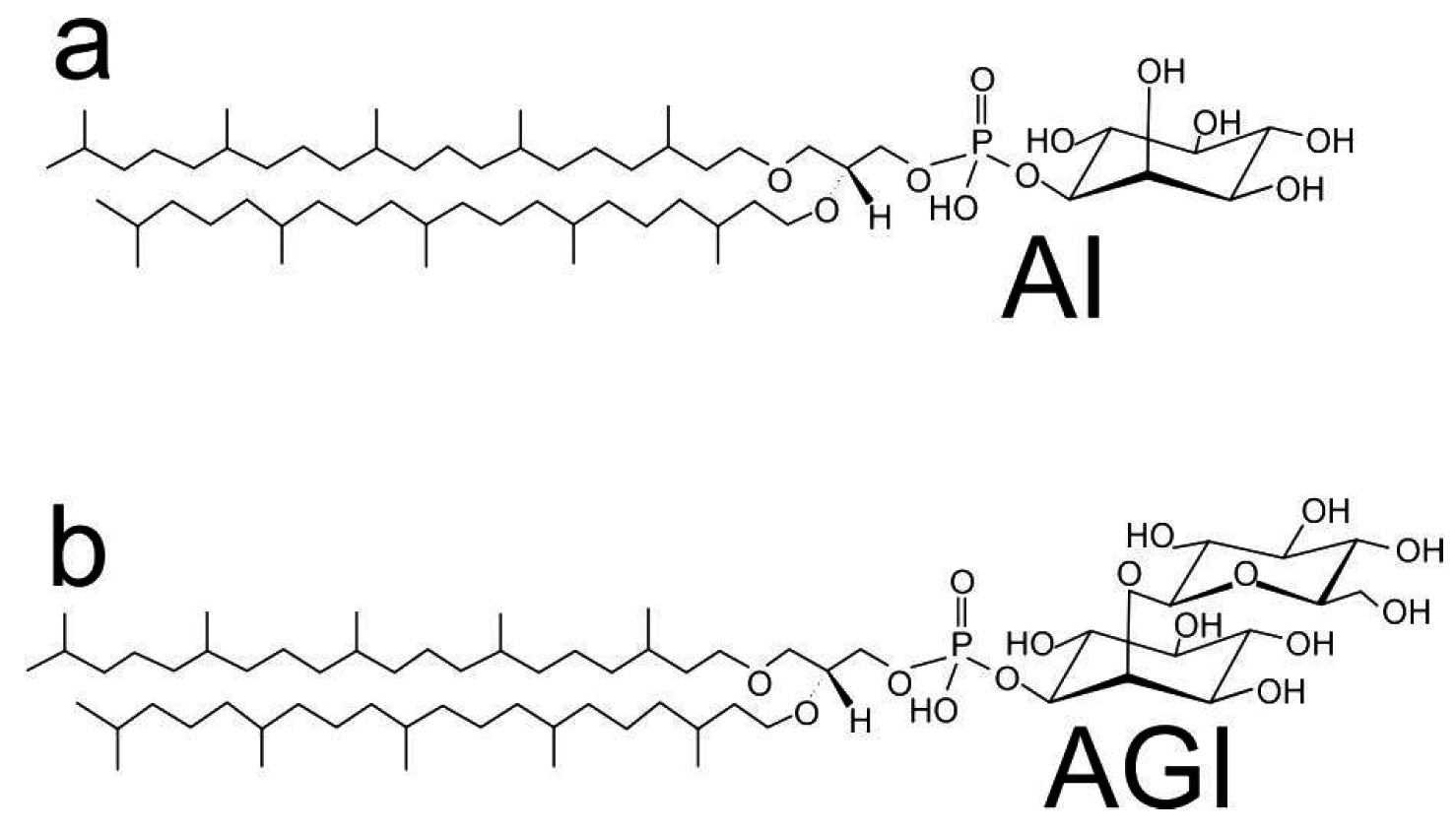
2. Materials and Methods
2.1. Bending Elasticity of Lipid Membrane
2.2. Thermally Induced Shape Fluctuation Method
2.3. Experimental Section
2.3.1. Chemicals
2.3.2. Growth of Aeropyrum Pernix K1
2.3.3. Isolation and Purification of Lipids
2.3.4. Formaton of Giant Vesicles

2.3.5. Observation and Recording of Giant Vesicles

3. Results and Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Lipid | Weighted Mean Value of the Bending Elasticity Modulus kc (± Standard Deviation) |
|---|---|
| Aeropyrum pernix K1 archaeal lipid | (1.89 ± 0.18) × 10−19 J |
| SOPC | (1.88 ± 0.17) × 10−19 J |
| POPC | (2.00 ± 0.21) × 10−19 J |
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Moghimipour, E.; Kargar, M.; Ramezani, Z.; Handali, S. The potent in vitro skin permeation of archaeosome made from lipids extracted of sulfolobus acidocaldarius. Archaea 2013, 2013. [Google Scholar] [CrossRef]
- Corcelli, A.; Lobasso, S. Characterization of lipids of halophilic archaea. Methods Microbiol. 2006, 35, 585–613. [Google Scholar]
- Angelova, M.I.; Soléau, S.; Méléard, P.; Faucon, F.; Bothorel, P. Preparation of giant vesicles by external AC electric fields. Kinetics and applications. In Trends in Colloid and Interface Science VI; Helm, C., Lösche, M., Möhwald, H., Eds.; Steinkopff: Darmstadt, Germany, 1992; pp. 127–131. [Google Scholar]
- Bagatolli, L.A.; Parasassi, T.; Gratton, E. Giant phospholipid vesicles: Comparison among the whole lipid sample characteristics using different preparation methods: A two photon fluorescence microscopy study. Chem. Phys. Lipids 2000, 105, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Peterlin, P.; Arrigler, V. Electroformation in a flow chamber with solution exchange as a means of preparation of flaccid giant vesicles. Colloids Surf. B Biointerfaces 2008, 64, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Pavlič, J.I.; Genova, J.; Zheliaskova, A.; Iglič, A.; Mitov, M.D. Electroformation of neutral and negatively charged phospholipid giant vesicles under physiological conditions. C. R. L’Academie Bulg. Sci. 2010, 63, 497–502. [Google Scholar]
- Vitkova, V.; Mader, M.A.; Polack, B.; Misbach, C.; Podgorski, T. Micro-macro link in rheology of erythrocyte and vesicle suspensions. Biophys. J. 2006, 95, L33–L35. [Google Scholar] [CrossRef]
- Genova, J.; Zheliaskova, A.; Mitov, M.D. Monosaccharides (fructose, glucose) and disaccharides (sucrose, trehalose) influence the elasticity of SOPC membranes. J. Optoelec. Adv. Mater. 2007, 9, 427–430. [Google Scholar]
- Mitkova, D.; Marukovich, N.; Ermakov, Y.A.; Vitkova, V. Bending rigidity of phosphatidylserine-containing lipid bilayers in acidic aqueous solutions. Colloids Surf. A Physicochem. Eng. Asp. 2014, 460, 71–78. [Google Scholar] [CrossRef]
- Ulrih, N.P.; Gmajner, D.; Raspor, P. Structural and physicochemical properties of polar lipids from thermophilic archaea. Appl. Microbiol. Biotechnol. 2009, 84, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Gmajner, D.; Ota, A.; Sentjurc, M.; Ulrih, N.P. Stability of diether C-25,C-25 liposomes from the hyperthermophilic archaeon Aeropyrum pernix K1. Chem. Phys. Lipids 2011, 164, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Morii, H.; Yagi, H.; Akutsu, H.; Nomura, N.; Sako, Y.; Koga, Y. A novel phosphoglycolipid archaetidyl(glucosyl)inositol with two sesterterpanyl chains from the aerobic hyperthermophilic archaeon Aeropyrum pernix K1. Biochim. Biophys. Acta 1999, 1436, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Sprott, G.D.; Tolson, D.L.; Patel, G.B. Archaeosomes as novel antigen delivery systems. FEMS Microbiol. Lett. 1997, 154, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Patel, G.B.; Sprott, G.D. Archaeobacterial ether lipid liposomes (archaeosomes) as novel vaccine and drug delivery systems. Crit. Rev. Biotechnol. 1999, 19, 317–357. [Google Scholar] [CrossRef] [PubMed]
- Bagatolli, L.; Gratton, E.; Khan, T.K.; Chong, P.L.G. Two-photon fluorescence microscopy studies of bipolar tetraether giant liposomes from thermoacidophilic archaebacteria Sulfolobus acidocaldarius. Biophys. J. 2000, 79, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Sustar, V.; Zelko, J.; Lopalco, P.; Lobasso, S.; Ota, A.; Ulrih, N.P.; Corcelli, A.; Kralj-iglic, V. Morphology, biophysical properties and protein-mediated fusion of archaeosomes. PLoS One 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Sprott, G.D.; Dicaire, C.J.; Cote, J.-P.; Whitfield, D.M. Adjuvant potential of archaeal synthetic glycolipid mimetics critically depends on the glyco head group structure. Glycobiology 2008, 18, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Kamath, A.T.; Rochat, A.-F.; Christensen, D.; Agger, E.M.; Andersen, P.; Lambert, P.-H.; Siegrist, C.-A. A Liposome-based mycobacterial vaccine induces potent adult and neonatal multifunctional T cells through the exquisite targeting of dendritic cells. PLoS One 2009, 4. [Google Scholar] [CrossRef] [PubMed]
- Bivas, I.; Hanusse, P.; Bothorel, P.; Lalanne, J.; Aguerrechariol, O. An application of the optical microscopy to the determination of the curvature elastic-modulus of biological and model membranes. J. Phys. 1987, 48, 855–867. [Google Scholar] [CrossRef]
- Bivas, I.; Bivolarski, L.; Mitov, M.; Derzhanski, A. Correlations between the form fluctuation modes of flaccid quasispherical lipid vesicles and their role in the calculation of the curvature elastic modulus of the vesicle membrane. Numerical results. J. Phys. II 1992, 2, 1423–1438. [Google Scholar]
- Bivas, I.; Meleard, P. Bending elasticity and bending fluctuations of lipid bilayer containing an additive. Phys. Rev. E 2003, 67. [Google Scholar] [CrossRef]
- Pontes, B.; Ayala, Y.; Fonseca, A.C.C.; Romao, L.F.; Amaral, R.F.; Salgado, L.T.; Lima, F.R.; Farina, M.; Viana, N.B.; Moura-Neto, V.; et al. Membrane elastic properties and cell function. PLoS One 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Bivas, I. Shape fluctuations of nearly spherical lipid vesicles and emulsion droplets. Phys. Rev. E 2010, 81. [Google Scholar] [CrossRef]
- Faucon, J.; Mitov, M.; Meleard, P.; Bivas, I.; Bothorel, P. Bending elasticity and thermal fluctuations of lipid-membranes—Theoretical and experimental requirements. J. Phys. 1989, 50, 2389–2414. [Google Scholar] [CrossRef]
- Meleard, P.; Gerbeaud, C.; Pott, T.; Fernandez-Puente, L.; Bivas, I.; Mitov, M.D.; Dufourcq, J.; Bothorel, P. Bending elasticities of model membranes: Influences of temperature and sterol content. Biophys. J. 1997, 72, 2616–2629. [Google Scholar] [CrossRef] [PubMed]
- Vitkova, V.; Genova, J.; Meleard, P. Influence of alamethicin on the passive water permeability of model lipid membranes and on the morphology of giant lipid vesicles. J. Mater. Sci. 2003, 14, 819–820. [Google Scholar]
- Genova, J.; Zheliaskova, A.; Vitkova, V.; Mitov, M.D. Stroboscopic illumination study of the dynamics of fluctuating vesicles. J. Optoelectron. Adv. Mater. 2009, 11, 1222–1225. [Google Scholar]
- Helfrich, W. Elastic properties of lipid bilayers—Theory and possible experiments. Z. Naturforschung C 1973, 28, 693–703. [Google Scholar]
- Milner, S.; Safran, S. Dynamic fluctuations of droplet microemulsions and vesicles. Phys. Rev. A 1987, 36, 4371–4379. [Google Scholar] [CrossRef] [PubMed]
- Mitov, M.D.; Faucon, J.F.; Meleard, P.; Bivas, I.; Bothorel, P. Thermal fluctuations of membranes. In Advances in Supramolecular Chemistry; Gokel, G.W., Ed.; JAI Press: Greenwich, CN, USA, 1992; Volume 2, pp. 93–139. [Google Scholar]
- Genova, J.; Vitkova, V.; Aladjem, L.; Meleard, P.; Mitov, M. Stroboscopic illumination gives new opportunities and improves the precision of bending elastic modulus measurements. J. Optoelectron. Adv. Mater. 2005, 7, 257–260. [Google Scholar]
- Genova, J.; Pavlič, J.I. Realization of Marin Mitov idea for the stroboscopic illumination used in optical microscopy. Bulg. J. Phys. 2012, 39, 65–71. [Google Scholar]
- Milek, I.; Cigić, B.; Skrt, M.; Kaletunç, G.; Ulrih, N.P. Optimization of growth for the hyperthermophilic archaeon Aeropyrum pernix on a small-batch scale. Can. J. Microbiol. 2005, 51, 805–809. [Google Scholar] [CrossRef] [PubMed]
- Genova, J.; Vitkova, V.; Bivas, I. Registration and analysis of the shape fluctuations of nearly spherical lipid vesicles. Phys. Rev. E 2013, 88. [Google Scholar] [CrossRef]
- Fernandez-Puente, L.; Bivas, I.; Mitov, M.D.; Meleard, P. Temperature and chain length effects on bending elasticity of phosphatidylcholine bilayers. Europhys. Lett. 1994, 28, 181–186. [Google Scholar] [CrossRef]
- Bedina Zavec, A.; Ota, A.; Zupančič, T.; Komel, R.; Poklar Ulrih, N.; Liović, M. Archeosomes can efficiently deliver different types of cargo into epithelial cells grown in vitro. J. Biotechnol. 2014, 192, 130–135. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Genova, J.; Ulrih, N.P.; Kralj-Iglič, V.; Iglič, A.; Bivas, I. Bending Elasticity Modulus of Giant Vesicles Composed of Aeropyrum Pernix K1 Archaeal Lipid. Life 2015, 5, 1101-1110. https://doi.org/10.3390/life5021101
Genova J, Ulrih NP, Kralj-Iglič V, Iglič A, Bivas I. Bending Elasticity Modulus of Giant Vesicles Composed of Aeropyrum Pernix K1 Archaeal Lipid. Life. 2015; 5(2):1101-1110. https://doi.org/10.3390/life5021101
Chicago/Turabian StyleGenova, Julia, Nataša Poklar Ulrih, Veronika Kralj-Iglič, Aleš Iglič, and Isak Bivas. 2015. "Bending Elasticity Modulus of Giant Vesicles Composed of Aeropyrum Pernix K1 Archaeal Lipid" Life 5, no. 2: 1101-1110. https://doi.org/10.3390/life5021101
APA StyleGenova, J., Ulrih, N. P., Kralj-Iglič, V., Iglič, A., & Bivas, I. (2015). Bending Elasticity Modulus of Giant Vesicles Composed of Aeropyrum Pernix K1 Archaeal Lipid. Life, 5(2), 1101-1110. https://doi.org/10.3390/life5021101