
Impact of Sucrose Consumption on the Metabolic, Immune, and Redox Profile of Mice with Gestational Diabetes Mellitus

 ,
,  ,
,  , , and
, , and
Abstract

1. Introduction
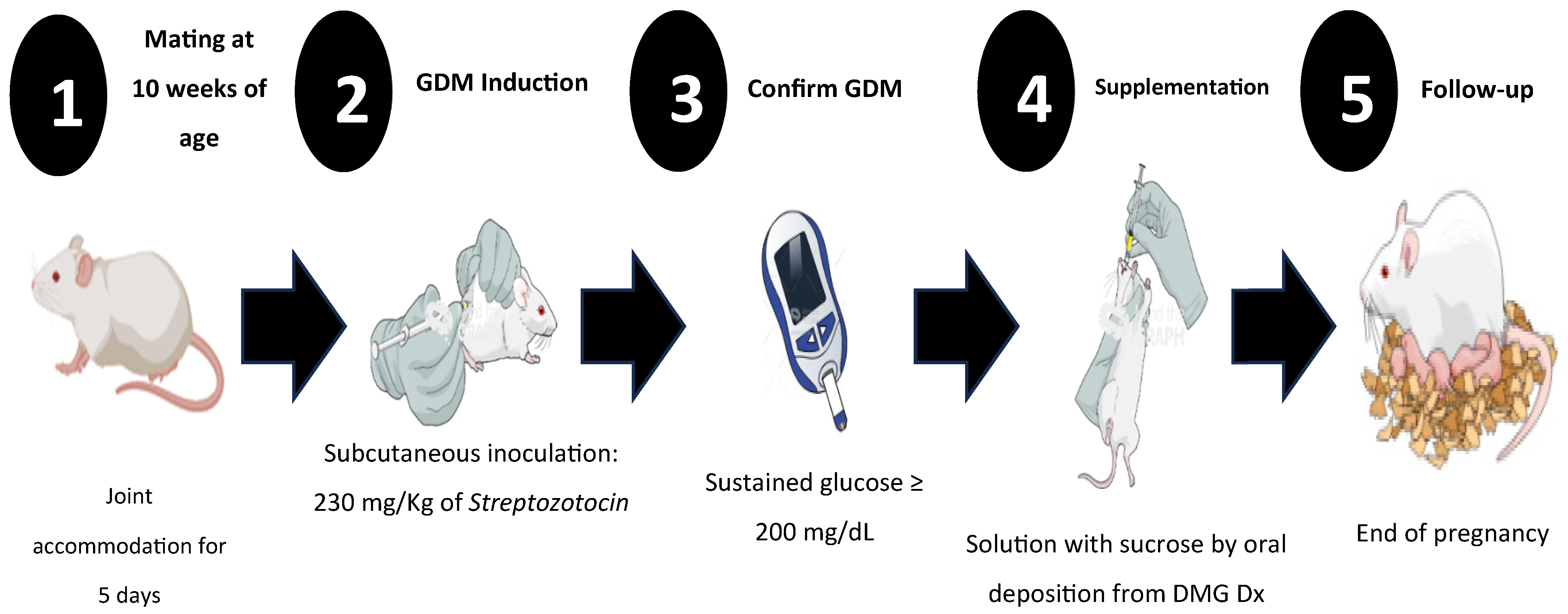
2. Materials and Methods
2.1. Study Design
2.2. Study Groups
2.3. Gestational Diabetes Mellitus Induction
2.4. Sucrose Supplementation
2.5. Calculation and Determination of Metabolic Parameters
2.6. Immunity Components Quantification
2.7. Redox Capacity Quantification
2.8. Statistical Analysis
3. Results
3.1. Morphometric Values and Glycemia
3.2. Metabolic Parameters
3.3. Immune Parameters
3.4. Redox Activity
4. Discussion
4.1. Changes in Weight and BMI Related to GDM
4.2. Insulin Resistance and GDM
4.3. Metabolic Changes During GDM
4.4. Immune Response in GDM
4.5. Changes in Humoral Response During GDM
4.6. Modifications of Redox Mechanisms During GDM
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DM | Diabetes Mellitus |
| T1DM | Type 1 Diabetes Mellitus |
| T2DM | Type 2 Diabetes Mellitus |
| GDM | Gestational Diabetes Mellitus |
| IR | Insulin Receptor |
| ISR-1 | Insulin Receptor Substrate 1 |
| GLUT4 | Glucose Transporter Protein Type 4 |
| BMI | Body Mass Index |
| IL-1β | Interleukin-1 Beta |
| IL-6 | Interleukin-6 |
| INF-γ | Interferon Gamma |
| TNF-α | Tumor Necrosis Factor-Alpha |
| ROS | Reactive Oxygen Species |
| OxS | Oxidant Stress |
| AGEs | Advanced Glycation End Products |
| TBARS | Thiobarbituric Acid Reactive Species |
| ARRIVE | Animal Research: Reporting of In Vivo Experiments |
| CL (−) | Control Group without Sucrose Supplementation |
| CL (+) | Control Group with Sucrose Supplementation |
| GDM (−) | Gestational Diabetes Mellitus group without Sucrose Supplementation |
| GDM (+) | Gestational Diabetes Mellitus group with Sucrose Supplementation |
| ELISA | Enzyme-Linked Immunosorbent Assay |
| HOMA Index | Homeostasis Model Assessment—Insulin Resistance Index |
| TAC | Total Antioxidant Capacity |
| SOD | Superoxide Dismutase |
| IgA | Immunoglobulin A |
| IgG | Immunoglobulin G |
References
- Kanguru, L.; Bezawada, N.; Hussein, J.; Bell, J. The burden of diabetes mellitus during pregnancy in low- and middle-income countries: A systematic review. Glob. Health Action 2014, 7, 23987. [Google Scholar] [CrossRef] [PubMed]
- Silva-Zolezzi, I.; Samuel, T.M.; Spieldenner, J. Maternal nutrition: Opportunities in the prevention of gestational diabetes. Nutr. Rev. 2017, 75, 32–50. [Google Scholar] [CrossRef]
- International Diabetes Federation. IDF Diabetes Atlas, 8th ed.; International Diabetes Federation: Brussels, Belgium, 2017; Available online: http://www.diabetesatlas.org (accessed on 20 July 2020).
- Moon, J.H.; Kwak, S.H.; Jang, H.C. Prevention of type 2 diabetes mellitus in women with previous gestational diabetes mellitus. Korean J. Intern. Med. 2017, 32, 26–41. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association Professional Practice Committee. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes—2022. Diabetes Care 2022, 45 (Suppl. S1), S17–S38. [Google Scholar] [CrossRef]
- Ye, W.; Luo, C.; Huang, J.; Li, C.; Liu, Z.; Liu, F. Gestational diabetes mellitus and adverse pregnancy outcomes: Systematic review and meta-analysis. BMJ 2022, 377, e067946. [Google Scholar] [CrossRef]
- Greco, E.; Calanducci, M.; Nicolaides, K.H.; Barry, E.V.H.; Huda, M.S.B.; Iliodromiti, S. Gestational diabetes mellitus and adverse maternal and perinatal outcomes in twin and singleton pregnancies: A systematic review and meta-analysis. Am. J. Obstet. Gynecol. 2024, 230, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Ornoy, A.; Becker, M.; Weinstein-Fudim, L.; Ergaz, Z. Diabetes during Pregnancy: A Maternal Disease Complicating the Course of Pregnancy with Long-Term Deleterious Effects on the Offspring. A Clinical Review. Int. J. Mol. Sci. 2021, 22, 2965. [Google Scholar] [CrossRef]
- Trumbo, P.; Schlicker, S.; Yates, A.A.; Poos, M. Food and Nutrition Board of the Institute of Medicine, The National Academies. Dietary reference intakes for energy, carbohydrate, fiber, fat, fatty acids, cholesterol, protein and amino acids. J. Am. Diet. Assoc. 2002, 102, 1621–1630. [Google Scholar] [CrossRef]
- Petersohn, I.; Hellinga, A.H.; van Lee, L.; Keukens, N.; Bont, L.; Hettinga, K.A.; Feskens, E.J.M.; Brouwer-Brolsma, E.M. Maternal diet and human milk composition: An updated systematic review. Front. Nutr. 2024, 10, 1320560. [Google Scholar] [CrossRef]
- Lovegrove, A.; Edwards, C.H.; De Noni, I.; Patel, H.; El, S.N.; Grassby, T.; Zielke, C.; Ulmius, M.; Nilsson, L.; Butterworth, P.J.; et al. Role of polysaccharides in food, digestion, and health. Crit. Rev. Food Sci. Nutr. 2017, 57, 237–253. [Google Scholar] [CrossRef]
- Mustad, V.A.; Huynh, D.T.; López-Pedrosa, J.M.; Campoy, C.; Rueda, R. The role of dietary carbohydrates in Gestational Diabetes. Nutrients 2020, 12, 385. [Google Scholar] [CrossRef]
- Lekva, T.; Norwitz, E.R.; Aukrust, P.; Ueland, T. Impact of systemic inflammation on the progression of gestational diabetes mellitus. Curr. Diabetes Rep. 2016, 16, 26. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.C.; Carpenter, M.W. Inflammatory mediators in gestational diabetes mellitus. Obstet. Gynecol. Clin. N. Am. 2007, 34, 213–224. [Google Scholar] [CrossRef]
- Saito, S.; Nakashima, A.; Shima, T.; Ito, M. Th1/Th2/Th17 and regulatory T-Cell paradigm in pregnancy. Am. J. Reprod. Immunol. 2010, 63, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Mor, G. Introduction to the immunology of pregnancy. Immunol. Rev. 2022, 308, 5–8. [Google Scholar] [CrossRef]
- Barbour, L.A.; McCurdy, C.E.; Hernandez, T.L.; Kirwan, J.P.; Catalano, P.M.; Friedman, J.E. Cellular mechanisms for insulin resistance in normal pregnancy and gestational diabetes. Diabetes Care 2007, 30, S112–S119. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Bachmann, R.A.; Chen, J. Interleukin-6 and insulin resistance. Vitam. Horm. 2009, 80, 613–633. [Google Scholar] [CrossRef]
- Kirwan, J.P.; Hauguel-De Mouzon, S.; Lepercq, J.; Challier, J.C.; Huston-Presley, L.; Friedman, J.E.; Kalhan, S.C.; Catalano, P.M. TNF-alpha is a predictor of insulin resistance in human pregnancy. Diabetes 2002, 51, 2207–2213. [Google Scholar] [CrossRef]
- Khalid, M.; Petroianu, G.; Adem, A. Advanced Glycation End Products and Diabetes Mellitus: Mechanisms and Perspectives. Biomolecules 2022, 12, 542. [Google Scholar] [CrossRef]
- Harsem, N.K.; Braekke, K.; Torjussen, T.; Hanssen, K.; Staff, A.C. Advanced glycation end products in pregnancies complicated with diabetes mellitus or preeclampsia. Hypertense Pregnancy 2008, 24, 374–386. [Google Scholar] [CrossRef]
- [NOM-062-ZOO-1999]. Diario Oficial de la Federación, México. 15. Organización Mundial de Sanidad Animal. Available online: https://www.gob.mx/cms/uploads/attachment/file/203498/NOM-062-ZOO-1999_220801.pdf (accessed on 17 November 2024).
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. Br. J. Pharmacol. 2020, 177, 3617–3624. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Carrillo, B.E.; Rosales-Gómez, C.A.; Ramírez-Durán, N.; Reséndiz-Albor, A.A.; Escoto-Herrera, J.A.; Mondragón-Velásquez, T.; Valdés-Ramos, R.; Castillo-Cardiel, A. Effect of Chronic Consumption of Sweeteners on Microbiota and Immunity in the Small Intestine of Young Mice. Int. J. Food Sci. 2019, 2019, 9619020. [Google Scholar] [CrossRef]
- Gutiérrez-Pliego, L.E.; Martínez-Carrillo, B.E.; Reséndiz-Albor, A.A.; Arciniega-Martínez, I.M.; Escoto-Herrera, J.A.; Rosales-Gómez, C.A.; Valdés-Ramos, R. Effect of Supplementation with n-3 Fatty Acids Extracted from Microalgae on Inflammation Biomarkers from Two Different Strains of Mice. J. Lipids 2018, 2018, 4765358. [Google Scholar] [CrossRef] [PubMed]
- Rosales-Gómez, C.A.; Martínez-Carrillo, B.E.; Guadarrama-López, A.L.; Reséndiz-Albor, A.A.; Arciniega-Martínez, I.M.; Aguilar-Rodríguez, E. Pharmacological induction of diabetes mellitus in pregnant female mice: A comparison of two doses and routes of administration. Eur. Rev. Med. Pharmacol. Sci. 2024, 28, 3275–3286. [Google Scholar] [CrossRef] [PubMed]
- ElSayed, N.A.; Aleppo, G.; Aroda, V.R.; Bannuru, R.R.; Brown, F.M.; Bruemmer, D.; Collins, B.S.; Gaglia, J.L.; Hilliard, M.E.; Isaacs, D.; et al. 2. Classification and Diagnosis of Diabetes: Standards of Care in Diabetes—2023. Diabetes Care 2023, 46 (Suppl. S1), S19–S40. [Google Scholar] [CrossRef] [PubMed]
- Rosales-Gómez, C.A.; Martínez-Carrillo, B.E.; Reséndiz-Albor, A.A.; Ramírez-Durán, N.; Valdés-Ramos, R.; Mondragón-Velásquez, T.; Escoto-Herrera, J.A. Chronic Consumption of Sweeteners and Its Effect on Glycaemia, Cytokines, Hormones, and Lymphocytes of GALT in CD1 Mice. BioMed Res. Int. 2018, 2018, 1345282. [Google Scholar] [CrossRef] [PubMed]
- Vilela, B.S.; Vasques, A.C.; Cassani, R.S.; Forti, A.C.; Pareja, J.C.; Tambascia, M.A.; BRAMS Investigators; Geloneze, B. The HOMA-Adiponectin (HOMA-AD) Closely Mirrors the HOMA-IR Index in the Screening of Insulin Resistance in the Brazilian Metabolic Syndrome Study (BRAMS). PLoS ONE 2016, 11, e0158751. [Google Scholar] [CrossRef]
- Carrizales-Luna, J.E.; Reséndiz-Albor, A.A.; Arciniega-Martínez, I.M.; Gómez-López, M.; Campos-Rodríguez, R.; Pacheco-Yépez, J.; Drago-Serrano, M.E. Outcomes of nicotinic modulation on markers of intestinal IgA antibody response. Biomed. Rep. 2022, 18, 13. [Google Scholar] [CrossRef]
- García-Iniesta, L.; Martínez-Carrillo, B.E.; Valdés-Ramos, R.; Jarillo-Luna, R.A.; Escoto-Herrera, J.A.; Reséndiz-Albor, A.A. Relationship between Prolonged Sweetener Consumption and Chronic Stress in the Production of Carbonylated Proteins in Blood Lymphocytes. Eur. J. Nutr. Food Saf. 2017, 7, 220–232. [Google Scholar] [CrossRef]
- Soma-Pillay, P.; Nelson-Piercy, C.; Tolppanen, H.; Mebazaa, A. Physiological changes in pregnancy. Cardiovasc. J. Afr. 2016, 27, 89–94. [Google Scholar] [CrossRef]
- Baz, B.; Riveline, J.P.; Gautier, J.F. Endocrinology of Pregnancy: Gestational diabetes mellitus: Definition, aetiological and clinical aspects. Eur. J. Endocrinol. 2016, 174, R43–R51. [Google Scholar] [CrossRef] [PubMed]
- Chandra, M.; Paray, A.A. Natural Physiological Changes During Pregnancy. Yale J. Biol. Med. 2024, 97, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Plows, J.F.; Stanley, J.L.; Baker, P.N.; Clare, M.R.; Vickers, M.H. The Pathophysiology of Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2018, 19, 3342. [Google Scholar] [CrossRef]
- Ryckman, K.K.; Borowski, K.S.; Parikh, N.I.; Saftlas, A.F. Pregnancy Complications and the Risk of Metabolic Syndrome for the Offspring. Curr. Cardiovasc. Risk Rep. 2013, 7, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Xiang, A.H.; Li, B.H.; Black, M.H.; Sacks, D.A.; Buchanan, T.A.; Jacobsen, S.J.; Lawrence, J.M. Racial and ethnic disparities in diabetes risk after gestational diabetes mellitus. Diabetologia 2011, 54, 3016–3021. [Google Scholar] [CrossRef]
- Pintaudi, B.; Fresa, R.; Dalfra, M.; Dodesini, A.R.; Vitacolonna, E.; Tumminia, A.; Sciacca, L.; Lencioni, C.; Marcone, T.; Lucisano, G.; et al. The risk stratification of adverse neonatal outcomes in women with gestational diabetes (STRONG) study. Acta Diabetol. 2018, 55, 1261–1273. [Google Scholar] [CrossRef]
- Mousa, A.; Naqash, A.; Lim, S. Macronutrient and Micronutrient Intake during Pregnancy: An Overview of Recent Evidence. Nutrients 2019, 11, 443. [Google Scholar] [CrossRef]
- Jovanovic-Peterson, L.; Peterson, C.M.; Reed, G.F.; Metzger, B.E.; Mills, J.L.; Knopp, R.H.; Aarons, J.H. The National Institute of Child Health and Human Development—Diabetes in Early Pregnancy Study Maternal postprandial glucose levels and infant birth weight: The Diabetes in Early Pregnancy Study. Am. J. Obstet. Gynecol. 1991, 164, 103–111. [Google Scholar] [CrossRef]
- Gou, B.H.; Guan, H.M.; Bi, Y.X.; Ding, B.J. Gestational diabetes: Weight gain during pregnancy and its relationship to pregnancy outcomes. Chin. Med. J. 2019, 132, 154–160. [Google Scholar] [CrossRef]
- Rayanagoudar, G.; Hashi, A.A.; Zamora, J.; Khan, K.S.; Hitman, G.A.; Thangaratinam, S. Quantification of the type 2 diabetes risk in women with gestational diabetes: A systematic review and meta-analysis of 95,750 women. Diabetologia 2016, 59, 1403–1411. [Google Scholar] [CrossRef]
- Mat, S.H.C.; Yaacob, N.M.; Hussain, S. Rate of weight gain and its association with homeostatic model assessment-insulin resistance (HOMA-IR) among obese children attending Paediatric Endocrine Clinic, Hospital Universiti Sains Malaysia. J. ASEAN Fed. Endocr. Soc. 2021, 36, 149–155. [Google Scholar] [CrossRef]
- Duo, Y.; Song, S.; Zhang, Y.; Qiao, X.; Xu, J.; Zhang, J.; Peng, Z.; Chen, Y.; Nie, X.; Sun, Q.; et al. Predictability of HOMA-IR for Gestational Diabetes Mellitus in Early Pregnancy Based on Different First Trimester BMI Values. J. Pers. Med. 2022, 13, 60. [Google Scholar] [CrossRef] [PubMed]
- Powe, C.E.; Allard, C.; Battista, M.C.; Doyon, M.; Bouchard, L.; Ecker, J.L.; Perron, P.; Florez, J.C.; Thadhani, R.; Hivert, M.-F. Heterogeneous contribution of insulin sensitivity and secretion defects to gestational Diabetes Mellitus. Diabetes Care 2016, 39, 1052–1055. [Google Scholar] [CrossRef] [PubMed]
- Cheney, C.; Shragg, P.; Hollingsworth, D. Demonstration of heterogeneity in gestational Diabetes by a 400-kcal breakfast meal tolerance test. Obstet. Gynecol. 1985, 65, 17–23. [Google Scholar] [PubMed]
- Hashemipour, S.; Zohal, M.; Modarresnia, L.; Kolaji, S.; Panahi, H.; Badri, M.; Chopani, S.M.; Kelishomi, S.E.; Ghasemi, A.; Ghafelehbashi, S.H. The yield of early-pregnancy homeostasis of model assessment-insulin resistance (HOMA-IR) for predicting gestational diabetes mellitus in different body mass index and age groups. BMC Pregnancy Childbirth 2023, 23, 822. [Google Scholar] [CrossRef]
- Rieck, S.; Kaestner, K.H. Expansion of beta-cell mass in response to pregnancy. Trends Endocrinol. Metab. 2010, 21, 151–158. [Google Scholar] [CrossRef]
- Usman, T.O.; Chhetri, G.; Yeh, H.; Dong, H.H. Beta-cell compensation and gestational diabetes. J. Biol. Chem. 2023, 299, 105405. [Google Scholar] [CrossRef]
- Inoue, S.; Kozuma, Y.; Miyahara, M.; Yoshizato, T.; Tajiri, Y.; Hori, D.; Ushijima, K. Pathophysiology of gestational Diabetes Mellitus in lean Japanese pregnant women in relation to insulin secretion or insulin resistance. Diabetol. Int. 2020, 11, 269–273. [Google Scholar] [CrossRef]
- Štambuk, T.; Kifer, D.; Smirčić-Duvnjak, L.; Vučić Lovrenčić, M.; Gornik, O. Associations between plasma protein, IgG and IgA N-glycosylation and metabolic health markers in pregnancy and gestational diabetes. PLoS ONE 2023, 18, e0284838. [Google Scholar] [CrossRef]
- Luo, N.; Liu, J.; Chung, B.H.; Yang, Q.; Klein, R.L.; Garvey, W.T.; Fu, Y. Macrophage Adiponectin Expression Improves Insulin Sensitivity and Protects Against Inflammation and Atherosclerosis. Diabetes 2010, 59, 791–799. [Google Scholar] [CrossRef]
- McElwain, C.J.; McCarthy, F.P.; McCarthy, C.M. Gestational Diabetes Mellitus and Maternal Immune Dysregulation: What We Know So Far. Int. J. Mol. Sci. 2021, 22, 4261. [Google Scholar] [CrossRef] [PubMed]
- Atègbo, J.M.; Grissa, O.; Yessoufou, A.; Hichami, A.; Dramane, K.L.; Moutairou, K.; Miled, A.; Grissa, A.; Jerbi, M.; Tabka, Z.; et al. Modulation of Adipokines and Cytokines in Gestational Diabetes and Macrosomia. J. Clin. Endocrinol. Metab. 2006, 91, 4137–4143. [Google Scholar] [CrossRef]
- Christian, L.M.; Porter, K. Longitudinal changes in serum proinflammatory markers across pregnancy and postpartum: Effects of maternal body mass index. Cytokine 2014, 70, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Moyce Gruber, B.L.; Dolinsky, V.W. The Role of Adiponectin during Pregnancy and Gestational Diabetes. Life 2023, 13, 301. [Google Scholar] [CrossRef]
- Pérez-Pérez, A.; Vilariño-García, T.; Guadix, P.; Dueñas, J.L.; Sánchez-Margalet, V. Leptin and nutrition in gestational diabetes. Nutrients 2020, 12, 1970. [Google Scholar] [CrossRef]
- De Gennaro, G.; Palla, G.; Battini, L.; Simoncini, T.; Del Prato, S.; Bertolotto, A.; Bianchi, C. The role of adipokines in the pathogenesis of gestational diabetes mellitus. Gynecol. Endocrinol. 2019, 35, 737–751. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.; Baecker, A.; Song, Y.; Kiely, M.; Liu, S.; Zhang, C. Adipokine levels during the first or early second trimester of pregnancy and subsequent risk of gestational diabetes mellitus: A systematic review. Metabolism 2015, 64, 756–764. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, Y.H.; Chen, Y.P.; Yuan, X.L.; Wang, J.; Zhu, H.; Lu, C.M. Maternal circulating concentrations of tumor necrosis factor-alpha, leptin, and adiponectin in gestational diabetes mellitus: A systematic review and meta-analysis. Sci. World J. 2014, 2014, 926932. [Google Scholar] [CrossRef]
- Mor, G.; Cardenas, I.; Abrahams, V.; Guller, S. Inflammation and pregnancy: The role of the immune system at the implantation site. Ann. N. Y. Acad. Sci. 2011, 1221, 80–87. [Google Scholar] [CrossRef]
- Berbudi, A.; Rahmadika, N.; Tjahjadi, A.I.; Ruslami, R. Type 2 diabetes and its impact on the immune system. Curr. Diabetes Rev. 2020, 16, 442–449. [Google Scholar] [CrossRef]
- O'Neill, L.A.; Kishton, R.J.; Rathmell, J. A guide to immunometabolism for immunologists. Nat. Rev. Immunol. 2016, 16, 553–565. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hu, T.; Liu, C.H.; Lei, M.; Zeng, Q.; Li, L.; Tang, H.; Zhang, N. Metabolic regulation of the immune system in health and diseases: Mechanisms and interventions. Signal Transduct. Target. Ther. 2024, 9, 268. [Google Scholar] [CrossRef] [PubMed]
- Cascone, T.; McKenzie, J.A.; Mbofung, R.M.; Punt, S.; Wang, Z.; Xu, C.; Williams, L.J.; Wang, Z.; Bristow, C.A.; Carugo, A.; et al. Increased Tumor Glycolysis Characterizes Immune Resistance to Adoptive T Cell Therapy. Cell Metab. 2018, 27, 977–987.e4. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation, metaflammation and immunometabolic disorders. Nature 2017, 542, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Sheu, A.; Chan, Y.; Ferguson, A.; Bakhtyari, M.B.; Hawke, W.; White, C.; Chan, Y.F.; Bertolino, P.J.; Woon, H.G.; Palendira, U.; et al. A proinflammatory CD4+ T cell phenotype in gestational diabetes mellitus. Diabetologia 2018, 61, 1633–1643. [Google Scholar] [CrossRef]
- Abell, S.K.; De Courten, B.; Boyle, J.A.; Teede, H.J. Inflammatory and Other Biomarkers: Role in Pathophysiology and Prediction of Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2015, 16, 13442–13473. [Google Scholar] [CrossRef]
- Sifnaios, E.; Mastorakos, G.; Psarra, K.; Panagopoulos, N.D.; Panoulis, K.; Vitoratos, N.; Rizos, D.; Creatsas, G. Gestational Diabetes and T-cell (Th1/Th2/Th17/Treg) immune profile. In Vivo 2019, 33, 31–40. [Google Scholar] [CrossRef]
- Seck, A.; Hichami, A.; Doucouré, S.; Diallo Agne, F.; Bassène, H.; Ba, A.; Sokhna, C.; Khan, N.A.; Samb, A. Th1/Th2 Dichotomy in Obese Women with Gestational Diabetes and Their Macrosomic Babies. J. Diabetes Res. 2018, 2018, 8474617. [Google Scholar] [CrossRef] [PubMed]
- Winer, D.A.; Winer, S.; Chng, M.H.; Shen, L.; Engleman, E.G. B Lymphocytes in obesity-related adipose tissue inflammation and insulin resistance. Cell. Mol. Life Sci. 2014, 71, 1033–1043. [Google Scholar] [CrossRef]
- Nikolajczyk, B.S.; Jagannathan-Bogdan, M.; Shin, H.; Gyurko, R. State of the union between metabolism and the immune system in type 2 diabetes. Genes Immun. 2011, 12, 239–250. [Google Scholar] [CrossRef]
- Palming, J.; Gabrielsson, B.G.; Jennische, E.; Smith, U.; Carlsson, B.; Carlsson, L.M.; Lönn, M. Plasma cells and Fc receptors in human adipose tissue—Lipogenic and anti-inflammatory effects of immunoglobulins on adipocytes. Biochem. Biophys. Res. Commun. 2006, 343, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Qi, Y.; Yi, H.; Mao, C.; Meng, Q.; Wang, H.; Zheng, C. The Roles of Adipose Tissue Macrophages in Human Disease. Front. Immunol. 2022, 13, 908749. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Zhang, J.; Li, Y.; Gu, H.; Zhao, J.; Sun, Y.; Wang, R.; Zhang, C.; Chen, W.; Weng, J.; et al. B Lymphocytes Are Predictors of Insulin Resistance in Women with Gestational Diabetes Mellitus. Endocr. Metab. Immune Disord. Drug Targets 2019, 19, 358–366. [Google Scholar] [CrossRef]
- Luck, H.; Khan, S.; Kim, J.H.; Copeland, J.K.; Revelo, X.S.; Tsai, S.; Chakraborty, M.; Cheng, K.; Chan, Y.T.; Nøhr, M.K.; et al. Gut-associated IgA+ immune cells regulate obesity-related insulin resistance. Nat. Commun. 2019, 10, 3650. [Google Scholar] [CrossRef]
- Nishikawa, T.; Edelstein, D.; Du, X.L.; Yamagishi, S.; Matsumura, T.; Kaneda, Y.; Yorek, M.A.; Beebe, D.J.; Oates, P.J.; Hammes, H.-P.; et al. Normalizing mitochondrial superoxide production blocks three pathways of hyperglycaemic damage. Nature 2000, 404, 787–790. [Google Scholar] [CrossRef]
- Sadowska, J.; Dudzińska, W.; Dziaduch, I. Effects of different models of sucrose intake on the oxidative status of the uterus and ovary of rats. PLoS ONE 2021, 16, e0251789. [Google Scholar] [CrossRef] [PubMed]
- Saucedo, R.; Ortega-Camarillo, C.; Ferreira-Hermosillo, A.; Díaz-Velázquez, M.F.; Meixueiro-Calderón, C.; Valencia-Ortega, J. Role of Oxidative Stress and Inflammation in Gestational Diabetes Mellitus. Antioxidants 2023, 12, 1812. [Google Scholar] [CrossRef]
- Batty, M.; Bennett, M.R.; Yu, E. The Role of Oxidative Stress in Atherosclerosis. Cells 2022, 11, 3843. [Google Scholar] [CrossRef]
- Maciejczyk, M.; Matczuk, J.; Żendzian-Piotrowska, M.; Niklińska, W.; Fejfer, K.; Szarmach, I.; Ładny, J.R.; Zieniewska, I.; Zalewska, A. Eight-Week Consumption of High-Sucrose Diet Has a Pro-Oxidant Effect and Alters the Function of the Salivary Glands of Rats. Nutrients 2018, 10, 1530. [Google Scholar] [CrossRef]
- Sadowska, J.; Dudzińska, W.; Skotnicka, E.; Sielatycka, K.; Daniel, I. The impact of a diet containing sucrose and systematically repeated starvation on the oxidative status of the uterus and ovary of rats. Nutrients 2019, 11, 1544. [Google Scholar] [CrossRef]
- Gasparini, P.V.F.; Matias, A.M.; Torezani-Sales, S.; Kobi, J.B.B.S.; Siqueira, J.S.; Corrêa, C.R.; Leopoldo, A.P.L. High-Fat and Combined High-Fat and Sucrose Diets Promote Cardiac Oxidative Stress Independent of Nox2 Redox Regulation and Obesity in Rats. Cell Physiol. Biochem. 2021, 55, 618–634. [Google Scholar] [CrossRef] [PubMed]
- Jarukamjorn, K.; Jearapong, N.; Pimson, C.; Chatuphonprasert, W. A High-Fat, High-Fructose Diet Induces Antioxidant Imbalance and Increases the Risk and Progression of Nonalcoholic Fatty Liver Disease in Mice. Scientifica 2016, 2016, 5029414. [Google Scholar] [CrossRef] [PubMed]
- Busserolles, J.; Rock, E.; Gueux, E.; Mazur, A.; Grolier, P.; Rayssiguier, Y. Short-term consumption of a high-sucrose diet has a pro-oxidant effect in rats. Br. J. Nutr. 2002, 87, 337–342. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| CL (−) Sucrose | CL (+) Sucrose | DMG (−) Sucrose | DMG (+) Sucrose | ||
|---|---|---|---|---|---|
| Mean ± SD (n = 6) | Mean ± SD (n = 6) | Mean ± SD (n = 6) | Mean ± SD (n = 6) | p * Value | |
| Weight (g) | 31.52 ± 2.67 | 27.65 ± 0.60 | 22.60 ± 1.68 | 25.38 ± 2.19 | 0.001 * |
| BMI (g/cm2) | 0.315 ± 0.03 | 0.263 ± 0.01 | 0.220 ± 0.02 | 0.233 ± 0.02 | 0.001 * |
| Median | Median | Median | Median | ||
| Glycemia (mg/dL) | 207 | 219 | 577 | 586 | 0.001 * |
| CL (−) Sucrose | CL (+) Sucrose | DMG (−) Sucrose | DMG (+) Sucrose | ||
|---|---|---|---|---|---|
| Mean ± SD (n = 6) | Mean ± SD (n = 6) | Mean ± SD (n = 6) | Mean ± SD (n = 6) | p Value * | |
| * 10th day, beginning of supplementation | |||||
| Insulin (pg/mL) | 0.743 ± 0.012 | 0.743 ± 0.012 | 0.743 ± 0.008 | 0.743 ± 0.008 | 0.994 |
| HOMA-IR Index | 0.239 ± 0.257 | 0.2557 ± 0.257 | 0.209 ± 0.031 | 0.226 ± 0.029 | 0.062 |
| Adiponectin (pg/mL) | 8.5178 ± 1.031 | 8.76 ± 0.883 | 9.29 ± 2.4 | 10.53 ± 0.751 | 0.102 |
| Leptin (pg/mL) | 0.058 ± 0.031 | 0.074 ± 0.009 | 0.028 ± 0.012 | 0.038 ± 0.009 | 0.001 * |
| & 23rd day, end of gestation and supplementation | |||||
| Insulin (pg/mL) | 0.438 ± 0.018 | 0.436 ± 0.014 | 0.443 ± 0.024 | 0.445 ± 0.005 | 0.792 |
| HOMA-IR Index | 0.394 ± 0.1564 | 0.397 ± 0.1050 | 1.026 ± 0.0718 | 1.027 ± 0.0886 | 0.001 * |
| Adiponectin (pg/mL) | 9.574 ± 0.778 | 7.792 ± 2.154 | 6.622 ± 3.139 | 7.631 ± 2.162 | 0.175 |
| Leptin (pg/mL) | 0.077 ± 0.029 | 0.101 ± 0.051 | 0.027 ± 0.018 | 0.043 ± 0.018 | 0.003 * |
| CL (−) Sucrose | CL (+) Sucrose | DMG (−) Sucrose | DMG (+) Sucrose | ||
|---|---|---|---|---|---|
| Mean ± SD (n = 6) | Mean ± SD (n = 6) | Mean ± SD (n = 6) | Mean ± SD (n = 6) | p Value * | |
| Cell Immunity | |||||
| CD3+ (%) | 26.41 ± 0.85 | 27.59 ± 0.71 | 26.05 ± 0.42 | 27.46 ± 0.95 | 0.004 * |
| CD3+/CD4+ (%) | 44.45 ± 0.77 | 42.80 ± 0.43 | 44.15 ± 0.91 | 41.78 ± 0.41 | 0.001 * |
| CD3+/CD8+ (%) | 26.37 ± 0.79 | 30.40 ± 0.85 | 27.05 ± 0.30 | 26.92 ± 0.78 | 0.001 * |
| IL-1β (pg/mL) | 0.46 ± 0.20 | 8.28 ± 0.35 | 10.63 ± 0.56 | 12.50 ± 0.20 | 0.001 * |
| IL-6 (pg/mL) | 1.08 ± 0.16 | 10.89 ± 0.80 | 7.39 ± 0.11 | 10.72 ± 0.19 | 0.001 * |
| INF-γ (pg/mL) | 0.20 ± 0.09 | 5.04 ± 0.46 | 6.36 ± 0.24 | 10.76 ± 0.51 | 0.001 * |
| TNF-α (pg/mL) | 0.33 ± 0.14 | 4.31 ± 0.08 | 5.49 ± 0.18 | 8.32 ± 0.31 | 0.001 * |
| Humoral Immunity | |||||
| IgA (OD) | 0.298 ± 0.037 | 0.537 ± 0.078 | 1.061 ± 0.627 | 0.939 ± 0.176 | 0.002 * |
| IgG (OD) | 0.247 ± 0.093 | 0.421 ± 0.080 | 0.545 ± 0.249 | 0.422 ± 0.083 | 0.017 * |
| CL (−) Sucrose | CL (+) Sucrose | DMG (−) Sucrose | DMG (+) Sucrose | ||
|---|---|---|---|---|---|
| Median (n = 6) | Median (n = 6) | Median (n = 6) | Median (n = 6) | p * Value | |
| Antioxidant Parameters | |||||
| TAC (µM) | 0.133 | 0.167 | 0.159 | 0.179 | 0.025 * |
| Catalase (U/L) | 0.182 | 0.168 | 0.193 | 0.172 | 0.138 |
| SOD (U/mL) | 1.274 | 0.925 | 1.823 | 2.047 | 0.019 * |
| GR (µM) | 98.86 | 92.06 | 87.53 | 83 | 0.049 * |
| Oxidant Parameters | |||||
| TBARS (µM) | 0.0218 | 0.0075 | 0.0625 | 0.1430 | 0.006 * |
| AGE (pM/dL) | 1218 | 1791 | 725 | 3034 | 0.001 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosales-Gómez, C.Á.; Martínez-Carrillo, B.E.; Guadarrama-López, A.L.; Reséndiz-Albor, A.A.; Arciniega-Martínez, I.M.; Aguilar-Rodríguez, E. Impact of Sucrose Consumption on the Metabolic, Immune, and Redox Profile of Mice with Gestational Diabetes Mellitus. Life 2025, 15, 989. https://doi.org/10.3390/life15070989
Rosales-Gómez CÁ, Martínez-Carrillo BE, Guadarrama-López AL, Reséndiz-Albor AA, Arciniega-Martínez IM, Aguilar-Rodríguez E. Impact of Sucrose Consumption on the Metabolic, Immune, and Redox Profile of Mice with Gestational Diabetes Mellitus. Life. 2025; 15(7):989. https://doi.org/10.3390/life15070989
Chicago/Turabian StyleRosales-Gómez, Cristian Ángel, Beatriz Elina Martínez-Carrillo, Ana Laura Guadarrama-López, Aldo Arturo Reséndiz-Albor, Ivonne Maciel Arciniega-Martínez, and Efrén Aguilar-Rodríguez. 2025. "Impact of Sucrose Consumption on the Metabolic, Immune, and Redox Profile of Mice with Gestational Diabetes Mellitus" Life 15, no. 7: 989. https://doi.org/10.3390/life15070989
APA StyleRosales-Gómez, C. Á., Martínez-Carrillo, B. E., Guadarrama-López, A. L., Reséndiz-Albor, A. A., Arciniega-Martínez, I. M., & Aguilar-Rodríguez, E. (2025). Impact of Sucrose Consumption on the Metabolic, Immune, and Redox Profile of Mice with Gestational Diabetes Mellitus. Life, 15(7), 989. https://doi.org/10.3390/life15070989