
The Effect of Environment on the Evolution and Proliferation of Protocells of Increasing Complexity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Dry Phase: Non-Enzymatic Replication
2.2. Dry Phase: Enzymatic Replication Using Ribozymes
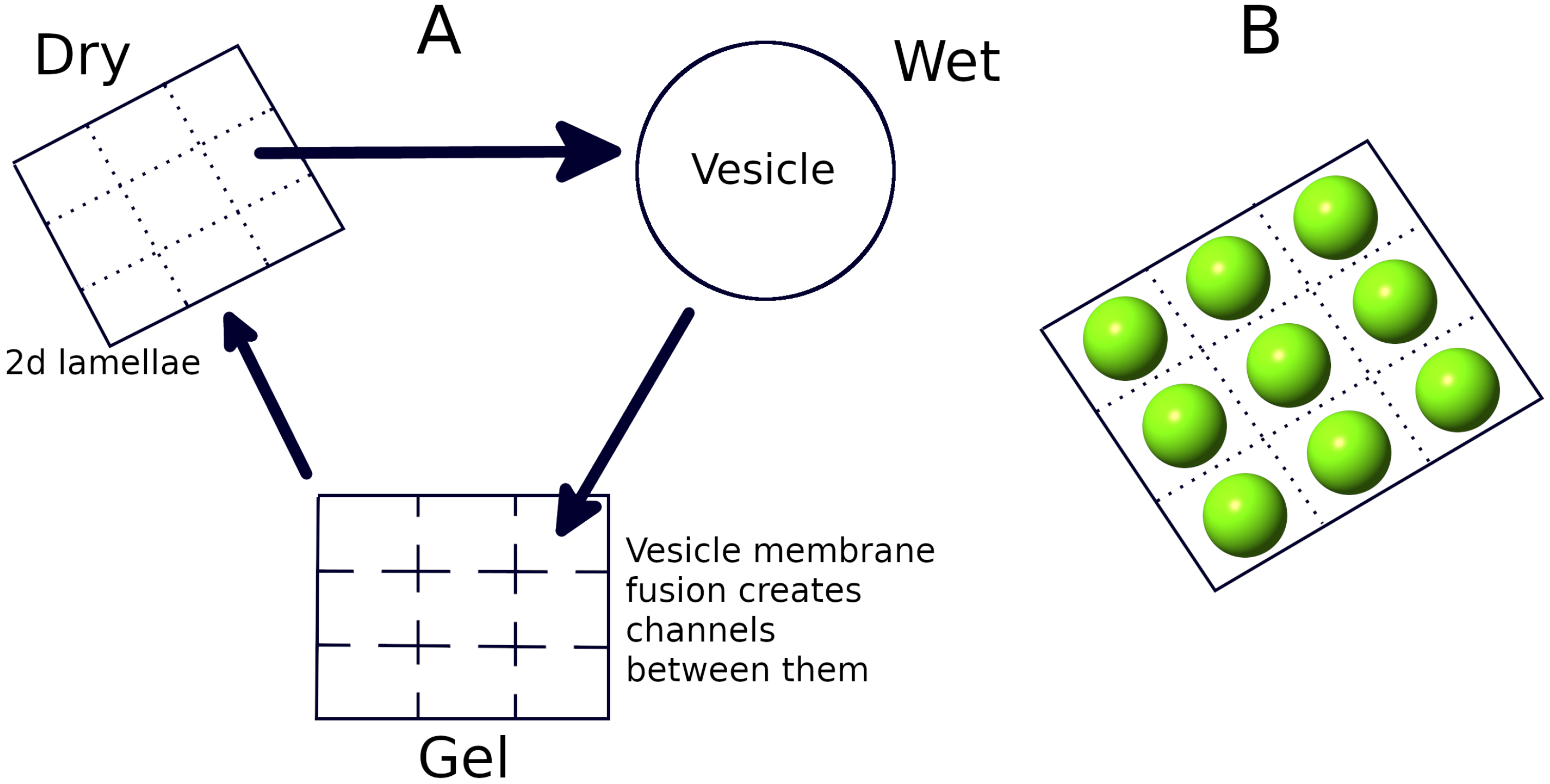
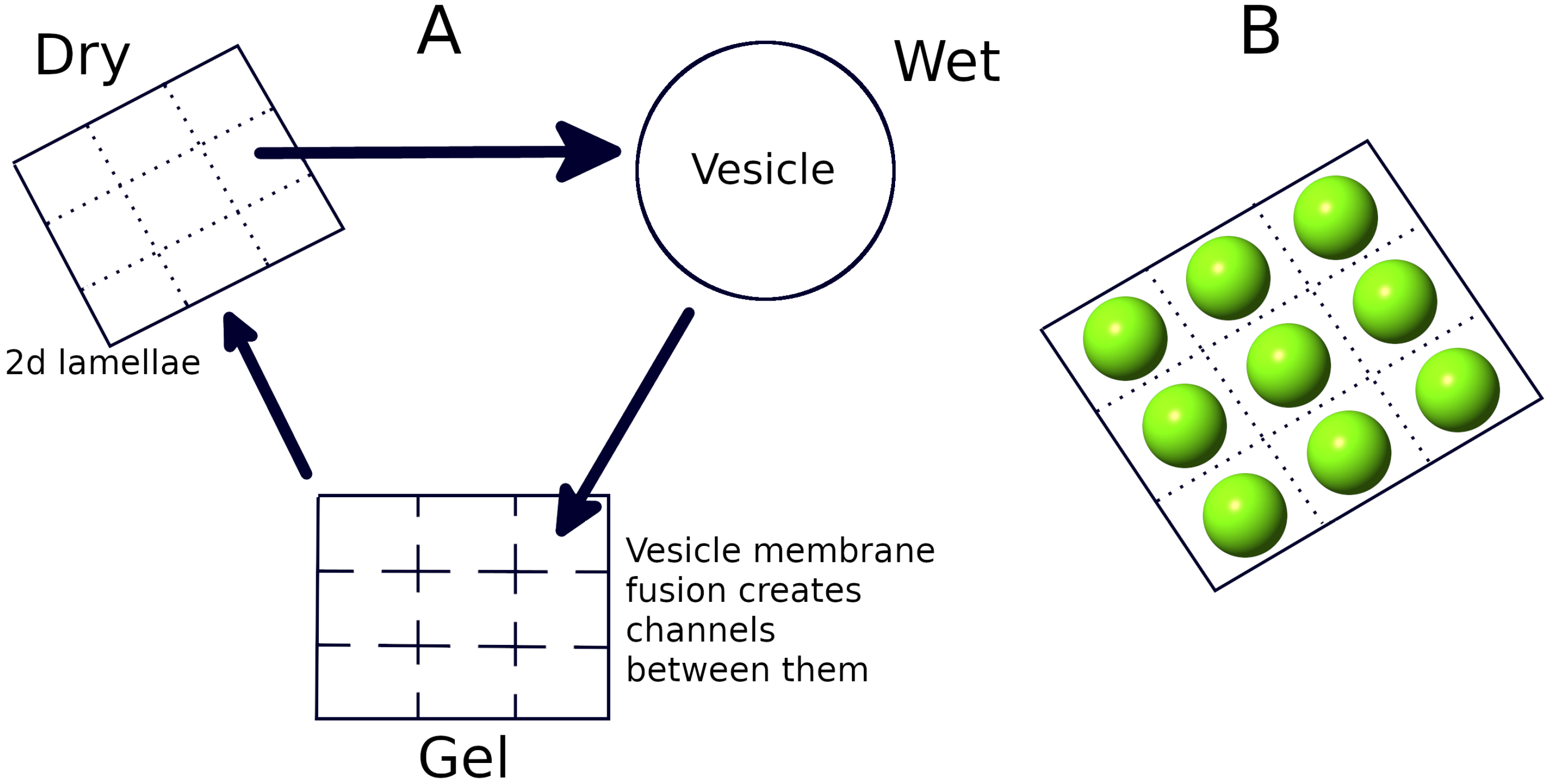
2.3. Wet Phase: Protocell Division
2.4. Gel Phase: Long Strand Diffusion
3. Results
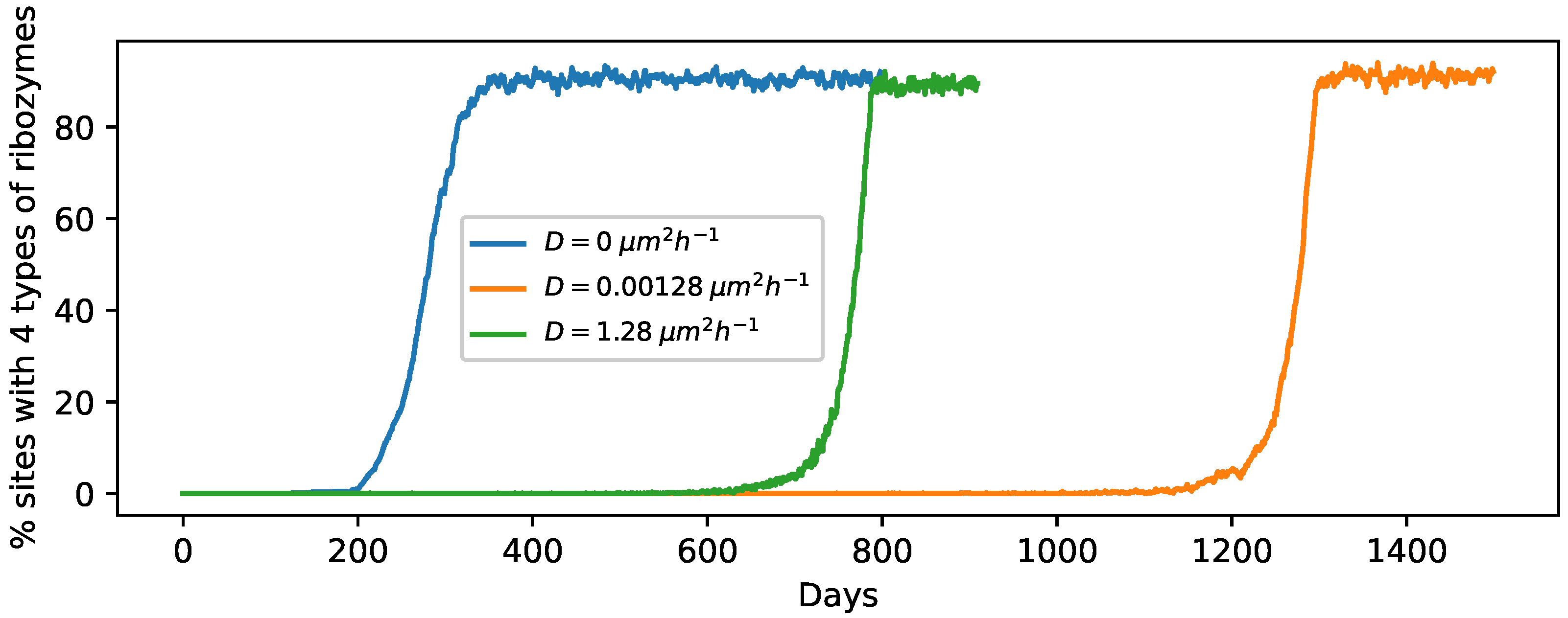
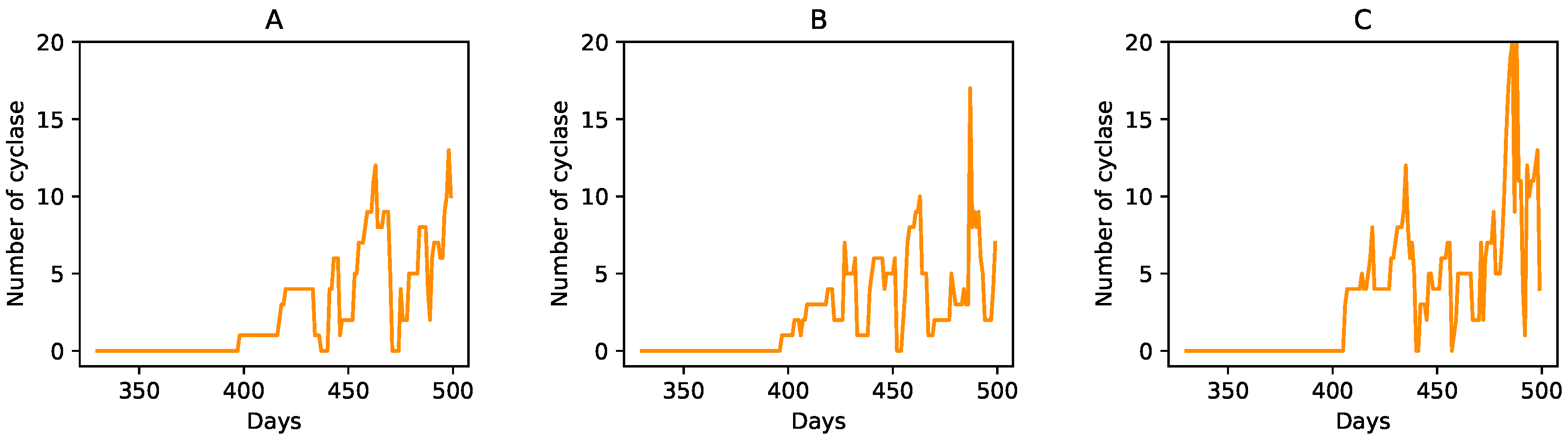
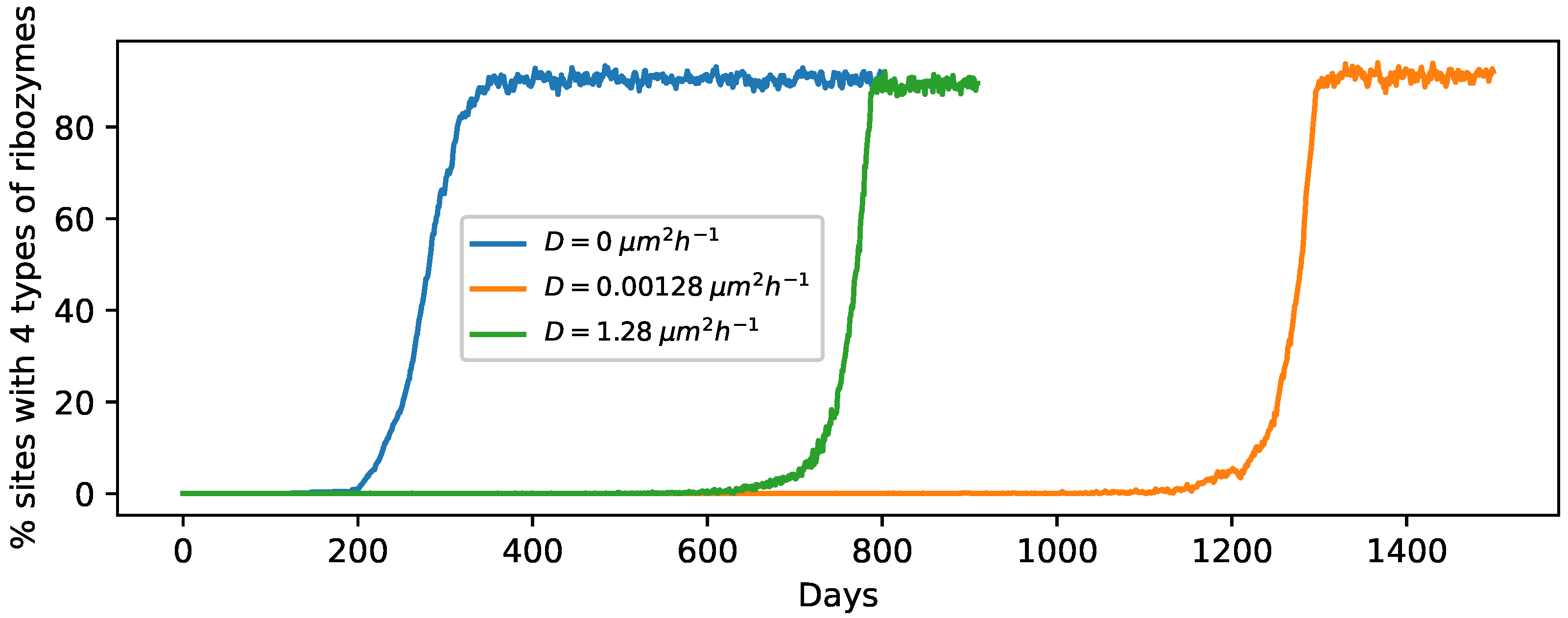
3.1. Effect of the Individual Phases
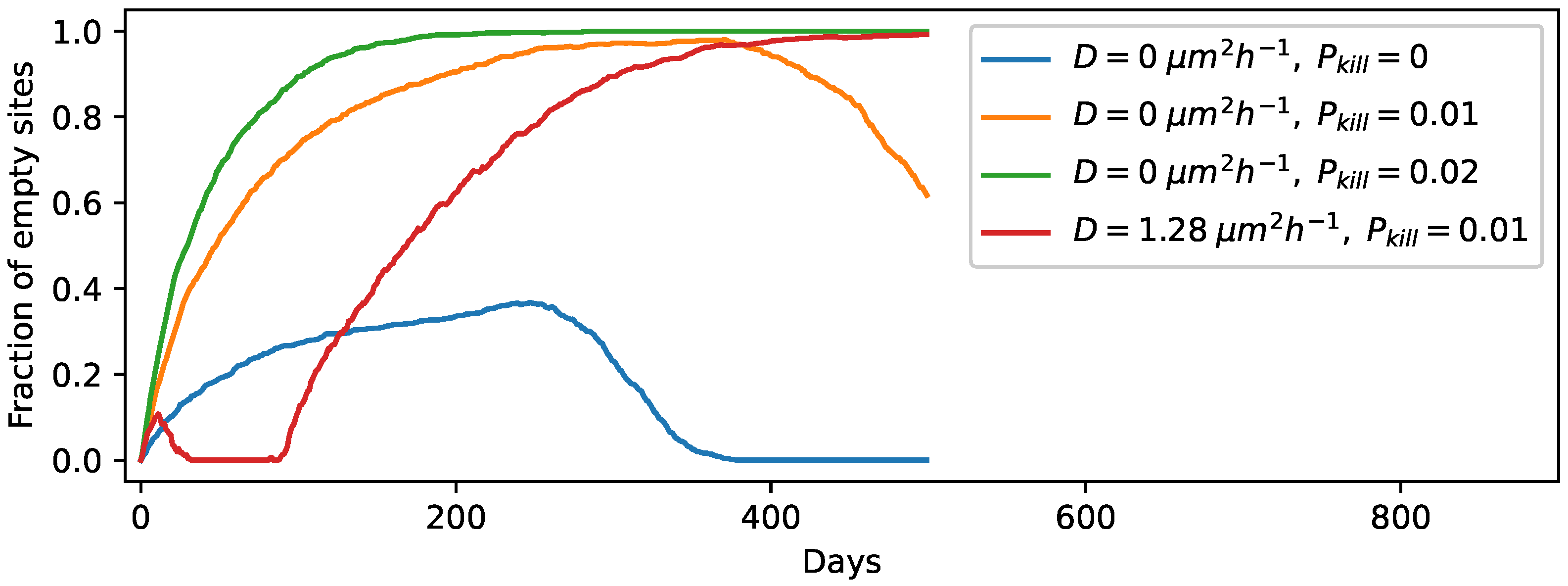
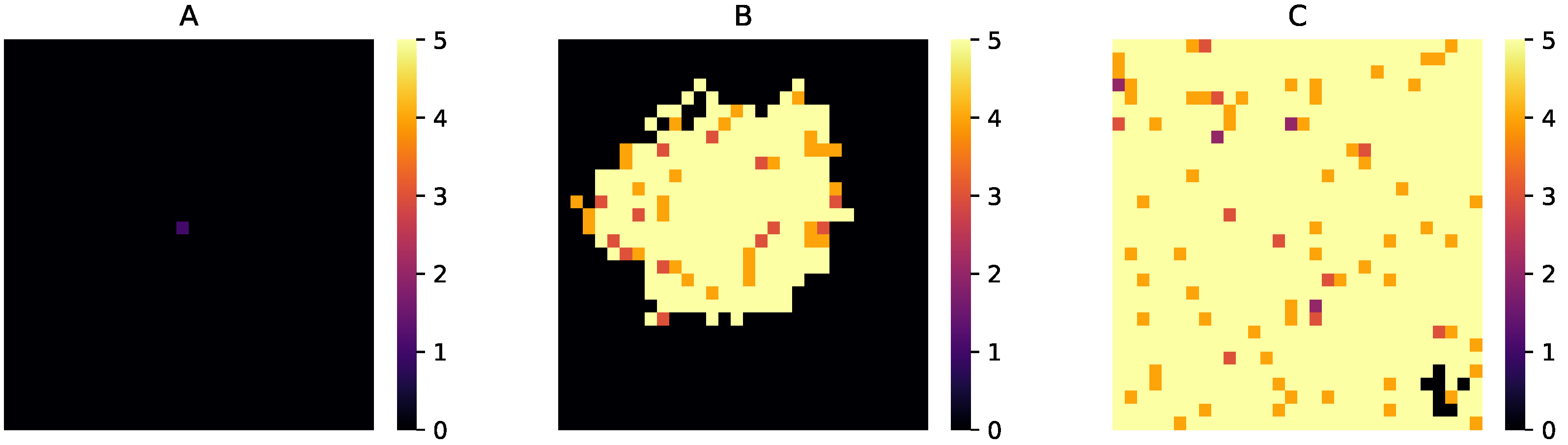
3.2. Relocation of Protocells in the Wet Phase
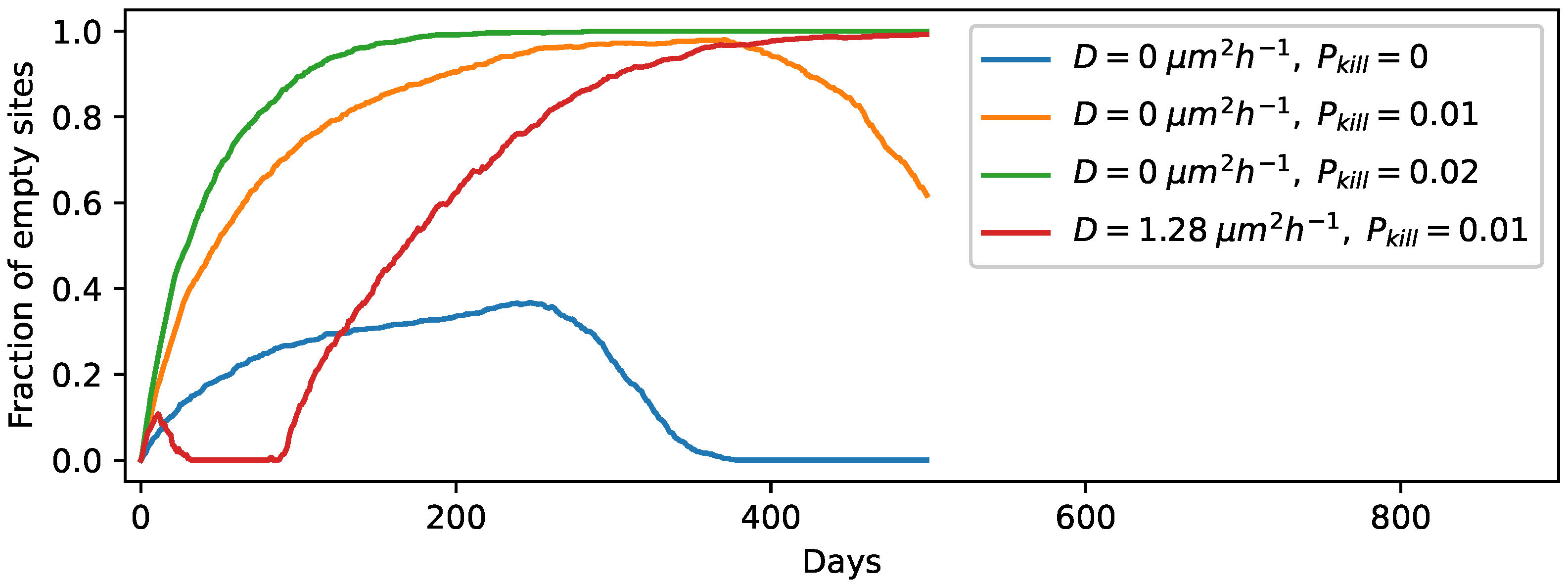
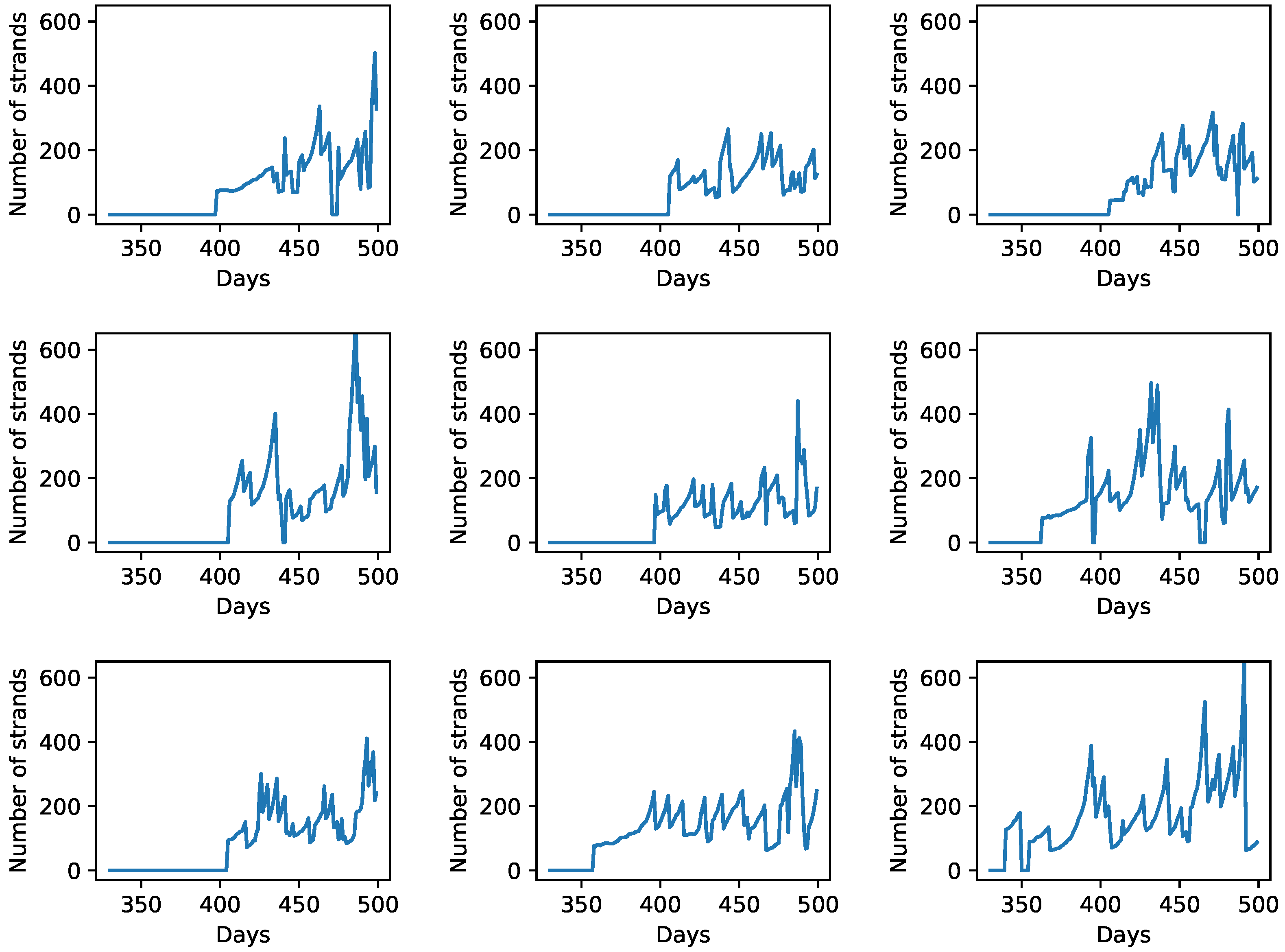
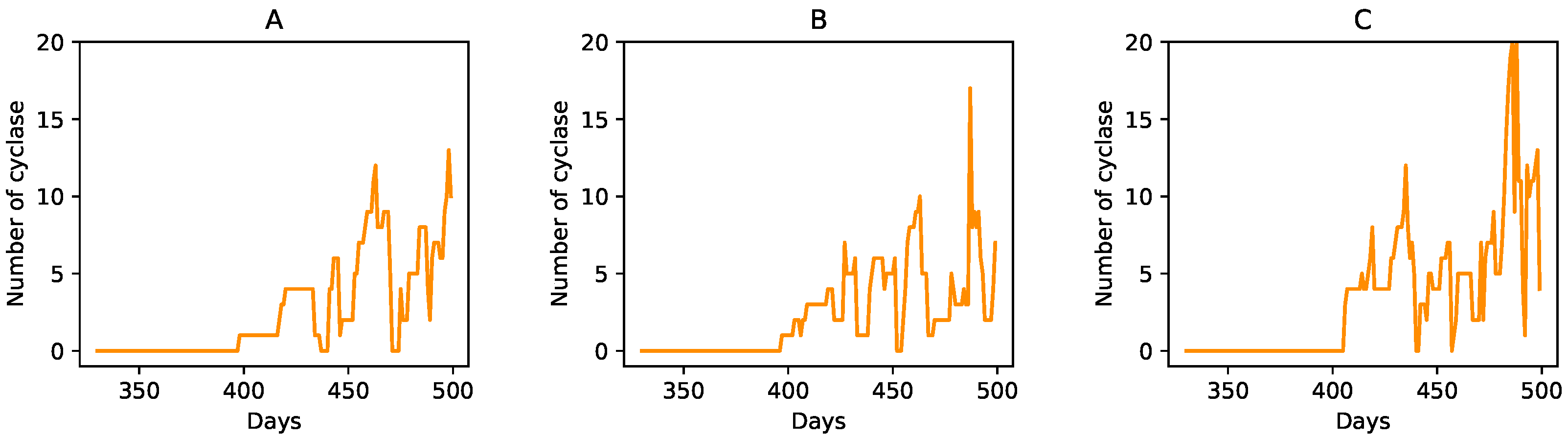
3.3. Degradation of Entire Protocells in the Wet Phase
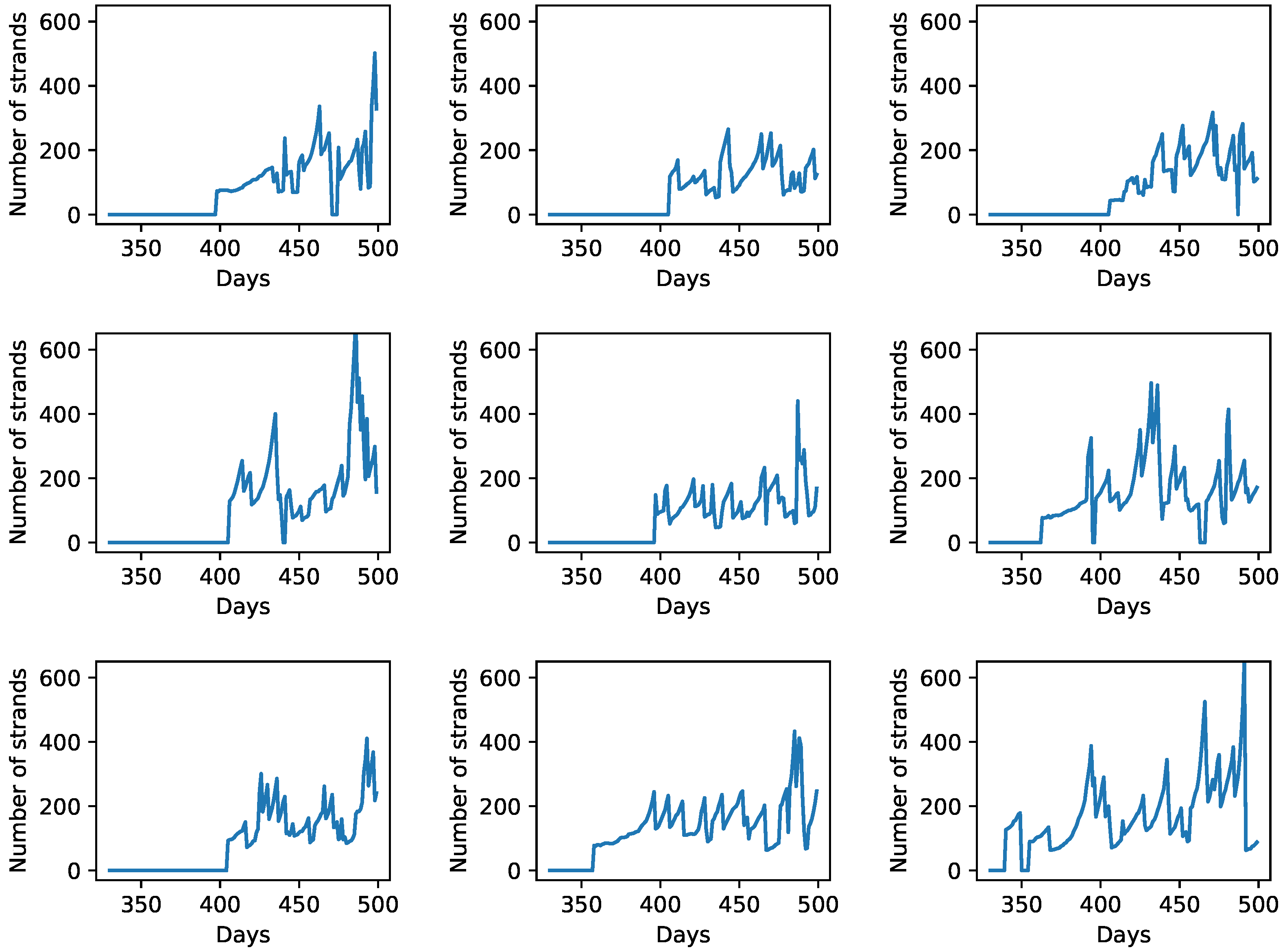
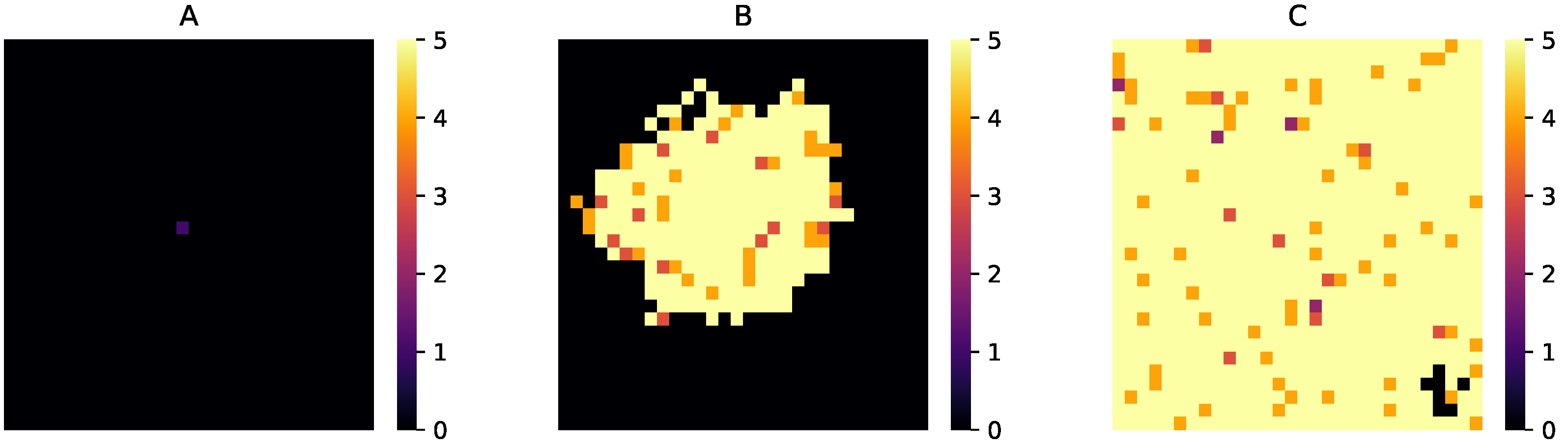
3.4. Spatial Expansion of Protocell Population
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Powner, M.W.; Gerland, B.; Sutherland, J.D. Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions. Nature 2009, 459, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Cafferty, B.J.; Fialho, D.M.; Khanam, J.; Krishnamurthy, R.; Hud, N.V. Spontaneous formation and base pairing of plausible prebiotic nucleotides in water. Nat. Commun. 2016, 7, 11328. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.; Feldmann, J.; Wiedemann, S.; Okamura, H.; Schneider, C.; Iwan, K.; Crisp, A.; Rossa, M.; Amatov, T.; Carell, T. Unified prebiotically plausible synthesis of pyrimidine and purine RNA ribonucleotides. Science 2019, 366, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Kruger, K.; Grabowski, P.J.; Zaug, A.J.; Sands, J.; Gottschling, D.E.; Cech, T.R. Self-splicing RNA: Autoexcision and autocyclization of the ribosomal RNA intervening sequence of tetrahymena. Cell 1982, 31, 147–157. [Google Scholar] [CrossRef]
- Stark, B.C.; Kole, R.; Bowman, E.J.; Altman, S. Ribonuclease P: An enzyme with an essential RNA component. Proc. Natl. Acad. Sci. USA 1978, 75, 3717–3721. [Google Scholar] [CrossRef]
- Guerrier-Takada, C.; Gardiner, K.; Marsh, T.; Pace, N.; Altman, S. The RNA moiety of ribonuclease P is the catalytic subunit of the enzyme. Cell 1983, 35, 849–857. [Google Scholar] [CrossRef]
- Robertson, D.L.; Joyce, G.F. Selection in vitro of an RNA enzyme that specifically cleaves single-stranded DNA. Nature 1990, 344, 467–468. [Google Scholar] [CrossRef]
- Jaeger, L.; Wright, M.C.; Joyce, G.F. A complex ligase ribozyme evolved in vitro from a group I ribozyme domain. Proc. Natl. Acad. Sci. USA 1999, 96, 14712–14717. [Google Scholar] [CrossRef]
- Horning, D.P.; Joyce, G.F. Amplification of RNA by an RNA polymerase ribozyme. Proc. Natl. Acad. Sci. USA 2016, 113, 9786–9791. [Google Scholar] [CrossRef]
- Miller, S.L.; Urey, H.C. Organic Compound Synthesis on the Primitive Earth. Science 1959, 130, 245–251. [Google Scholar] [CrossRef]
- Hargreaves, W.R.; Mulvihill, S.J.; Deamer, D.W. Synthesis of phospholipids and membranes in prebiotic conditions. Nature 1977, 266, 78–80. [Google Scholar] [CrossRef] [PubMed]
- Bangham, A.; Horne, R. Negative staining of phospholipids and their structural modification by surface-active agents as observed in the electron microscope. J. Mol. Biol. 1964, 8, 660-IN10. [Google Scholar] [CrossRef]
- Gebicki, J.M.; Hicks, M. Ufasomes are Stable Particles surrounded by Unsaturated Fatty Acid Membranes. Nature 1973, 243, 232–234. [Google Scholar] [CrossRef] [PubMed]
- Rajamani, S.; Vlassov, A.; Benner, S.; Coombs, A.; Olasagasti, F.; Deamer, D. Lipid-assisted Synthesis of RNA-like Polymers from Mononucleotides. Orig. Life Evol. Biosph. 2007, 38, 57–74. [Google Scholar] [CrossRef]
- Bianconi, G.; Zhao, K.; Chen, I.A.; Nowak, M.A. Selection for Replicases in Protocells. PLoS Comput. Biol. 2013, 9, e1003051. [Google Scholar] [CrossRef]
- Joyce, G.F.; Szostak, J.W. Protocells and RNA Self-Replication. Cold Spring Harb. Perspect. Biol. 2018, 10, a034801. [Google Scholar] [CrossRef]
- Kamimura, A.; Matsubara, Y.J.; Kaneko, K.; Takeuchi, N. Horizontal transfer between loose compartments stabilizes replication of fragmented ribozymes. PLoS Comput. Biol. 2019, 15, e1007094. [Google Scholar] [CrossRef]
- Black, R.A.; Blosser, M.C.; Stottrup, B.L.; Tavakley, R.; Deamer, D.W.; Keller, S.L. Nucleobases bind to and stabilize aggregates of a prebiotic amphiphile, providing a viable mechanism for the emergence of protocells. Proc. Natl. Acad. Sci. USA 2013, 110, 13272–13276. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.A.; Roberts, R.W.; Szostak, J.W. The Emergence of Competition Between Model Protocells. Science 2004, 305, 1474–1476. [Google Scholar] [CrossRef]
- Cornell, C.E.; Black, R.A.; Xue, M.; Litz, H.E.; Ramsay, A.; Gordon, M.; Mileant, A.; Cohen, Z.R.; Williams, J.A.; Lee, K.K.; et al. Prebiotic amino acids bind to and stabilize prebiotic fatty acid membranes. Proc. Natl. Acad. Sci. USA 2019, 116, 17239–17244. [Google Scholar] [CrossRef]
- Adamala, K.; Szostak, J.W. Competition between model protocells driven by an encapsulated catalyst. Nat. Chem. 2013, 5, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Hanczyc, M.M.; Fujikawa, S.M.; Szostak, J.W. Experimental Models of Primitive Cellular Compartments: Encapsulation, Growth, and Division. Science 2003, 302, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, S.; Spoelstra, W.K.; van Doorn, M.; Kerssemakers, J.; Dekker, C. Mechanical Division of Cell-Sized Liposomes. ACS Nano 2018, 12, 2560–2568. [Google Scholar] [CrossRef]
- Ma, W.; Yu, C.; Zhang, W.; Zhou, P.; Hu, J. The emergence of ribozymes synthesizing membrane components in RNA-based protocells. Biosystems 2010, 99, 201–209. [Google Scholar] [CrossRef]
- Ma, W.; Hu, J. Computer Simulation on the Cooperation of Functional Molecules during the Early Stages of Evolution. PLoS ONE 2012, 7, e35454. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Chen, Y.; Yu, C.; Ma, W. From molecular to cellular form: Modeling the first major transition during the arising of life. BMC Evol. Biol. 2019, 19, 84. [Google Scholar] [CrossRef]
- Shah, V.; de Bouter, J.; Quinn, P.; Tupper, A.; Higgs, P.G. Survival of RNA Replicators is much Easier in Protocells than in Surface-Based, Spatial Systems. Life 2019, 9, 65. [Google Scholar] [CrossRef] [PubMed]
- Tupper, A.S.; Higgs, P.G. Rolling-circle and strand-displacement mechanisms for non-enzymatic RNA replication at the time of the origin of life. J. Theor. Biol. 2021, 527, 110822. [Google Scholar] [CrossRef]
- Shay, J.A.; Huynh, C.; Higgs, P.G. The origin and spread of a cooperative replicase in a prebiotic chemical system. J. Theor. Biol. 2015, 364, 249–259. [Google Scholar] [CrossRef]
- Kim, Y.E.; Higgs, P.G. Co-operation between Polymerases and Nucleotide Synthetases in the RNA World. PLoS Comput. Biol. 2016, 12, e1005161. [Google Scholar] [CrossRef]
- Roy, S.; Bapat, N.V.; Derr, J.; Rajamani, S.; Sengupta, S. Emergence of ribozyme and tRNA-like structures from mineral-rich muddy pools on prebiotic earth. J. Theor. Biol. 2020, 506, 110446. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Sengupta, S. Evolution towards increasing complexity through functional diversification in a protocell model of the RNA world. Proc. R. Soc. Biol. Sci. 2021, 288, 20212098. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Ding, D.; Szostak, J.W. The virtual circular genome model for primordial RNA replication. RNA 2020, 27, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Corliss, J.B.; Baross, J.A.; Hoffman, S.E. An Hypothesis Concerning the Relationships Between Submarine Hot Springs and the Origin of Life on Earth. In Proceedings of the 26 International Geological Congress, Paris, France, 7 July 1980; pp. 59–69. [Google Scholar]
- Damer, B.; Deamer, D. The Hot Spring Hypothesis for an Origin of Life. Astrobiology 2020, 20, 429–452. [Google Scholar] [CrossRef] [PubMed]
- Obermayer, B.; Krammer, H.; Braun, D.; Gerland, U. Emergence of Information Transmission in a Prebiotic RNA Reactor. Phys. Rev. Lett. 2011, 107, 018101. [Google Scholar] [CrossRef] [PubMed]
- Mast, C.B.; Schink, S.; Gerland, U.; Braun, D. Escalation of polymerization in a thermal gradient. Proc. Natl. Acad. Sci. USA 2013, 110, 8030–8035. [Google Scholar] [CrossRef] [PubMed]
- Kreysing, M.; Keil, L.; Lanzmich, S.; Braun, D. Heat flux across an open pore enables the continuous replication and selection of oligonucleotides towards increasing length. Nat. Chem. 2015, 7, 203–208. [Google Scholar] [CrossRef]
- Göppel, T.; Rosenberger, J.H.; Altaner, B.; Gerland, U. Thermodynamic and Kinetic Sequence Selection in Enzyme-Free Polymer Self-Assembly inside a Non-equilibrium RNA Reactor. Life 2022, 12, 567. [Google Scholar] [CrossRef]
- Ferris, J.P.; Hill, A.R.; Liu, R.; Orgel, L.E. Synthesis of long prebiotic oligomers on mineral surfaces. Nature 1996, 381, 59–61. [Google Scholar] [CrossRef]
- Duhr, S.; Braun, D. Why molecules move along a temperature gradient. Proc. Natl. Acad. Sci. USA 2006, 103, 19678–19682. [Google Scholar] [CrossRef]
- Milshteyn, D.; Damer, B.; Havig, J.; Deamer, D. Amphiphilic Compounds Assemble into Membranous Vesicles in Hydrothermal Hot Spring Water but Not in Seawater. Life 2018, 8, 11. [Google Scholar] [CrossRef] [PubMed]
- Damer, B.; Deamer, D. Coupled Phases and Combinatorial Selection in Fluctuating Hydrothermal Pools: A Scenario to Guide Experimental Approaches to the Origin of Cellular Life. Life 2015, 5, 872–887. [Google Scholar] [CrossRef] [PubMed]
- Kranendonk, M.J.V.; Baumgartner, R.; Djokic, T.; Ota, T.; Steller, L.; Garbe, U.; Nakamura, E. Elements for the Origin of Life on Land: A Deep-Time Perspective from the Pilbara Craton of Western Australia. Astrobiology 2021, 21, 39–59. [Google Scholar] [CrossRef] [PubMed]
- Deamer, D.; Damer, B.; Kompanichenko, V. Hydrothermal Chemistry and the Origin of Cellular Life. Astrobiology 2019, 19, 1523–1537. [Google Scholar] [CrossRef] [PubMed]
- Higgs, P. The Effect of Limited Diffusion and Wet–Dry Cycling on Reversible Polymerization Reactions: Implications for Prebiotic Synthesis of Nucleic Acids. Life 2016, 6, 24. [Google Scholar] [CrossRef]
- Deamer, D.W.; Barchfeld, G.L. Encapsulation of macromolecules by lipid vesicles under simulated prebiotic conditions. J. Mol. Evol. 1982, 18, 203–206. [Google Scholar] [CrossRef]
- Damer, B. A Field Trip to the Archaean in Search of Darwin’s Warm Little Pond. Life 2016, 6, 21. [Google Scholar] [CrossRef]
- Joshi, M.; Samanta, A.; Tripathy, G.; Rajamani, S. Formation and Stability of Prebiotically Relevant Vesicular Systems in Terrestrial Geothermal Environments. Life 2017, 7, 51. [Google Scholar] [CrossRef]
- Deamer, D. Where Did Life Begin? Testing Ideas in Prebiotic Analogue Conditions. Life 2021, 11, 134. [Google Scholar] [CrossRef]
- Ross, D.; Deamer, D. Dry/Wet Cycling and the Thermodynamics and Kinetics of Prebiotic Polymer Synthesis. Life 2016, 6, 28. [Google Scholar] [CrossRef]
- Bapat, N.V.; Rajamani, S. Effect of Co-solutes on Template-Directed Nonenzymatic Replication of Nucleic Acids. J. Mol. Evol. 2015, 81, 72–80. [Google Scholar] [CrossRef]
- Hassenkam, T.; Damer, B.; Mednick, G.; Deamer, D. AFM Images of Viroid-Sized Rings That Self-Assemble from Mononucleotides through Wet–Dry Cycling: Implications for the Origin of Life. Life 2020, 10, 321. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, D.K.; Kamat, N.P.; Mirza, F.N.; Li, L.; Prywes, N.; Szostak, J.W. Copying of Mixed-Sequence RNA Templates inside Model Protocells. J. Am. Chem. Soc. 2018, 140, 5171–5178. [Google Scholar] [CrossRef]
- Kusumoto-Matsuo, R.; Kanda, T.; Kukimoto, I. Rolling circle replication of human papillomavirus type 16 DNA in epithelial cell extracts. Genes Cells 2010, 16, 23–33. [Google Scholar] [CrossRef]
- Daròs, J.A.; Elena, S.F.; Flores, R. Viroids: An Ariadne’s thread into the RNA labyrinth. EMBO Rep. 2006, 7, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Flores, R.; Gas, M.E.; Molina-Serrano, D.; Nohales, M.Á.; Carbonell, A.; Gago, S.; la Peña, M.D.; Daròs, J.A. Viroid Replication: Rolling-Circles, Enzymes and Ribozymes. Viruses 2009, 1, 317–334. [Google Scholar] [CrossRef] [PubMed]
- Leu, K.; Kervio, E.; Obermayer, B.; Turk-MacLeod, R.M.; Yuan, C.; Luevano, J.M.; Chen, E.; Gerland, U.; Richert, C.; Chen, I.A. Cascade of Reduced Speed and Accuracy after Errors in Enzyme-Free Copying of Nucleic Acid Sequences. J. Am. Chem. Soc. 2012, 135, 354–366. [Google Scholar] [CrossRef]
- Dingle, K.; Ghaddar, F.; Šulc, P.; Louis, A.A. Phenotype bias determines how natural RNA structures occupy the morphospace of all possible shapes. Mol. Biol. Evol. 2021, 39, msab280. [Google Scholar] [CrossRef]
- Shea, S.M.; Karnovsky, M.J. Brownian Motion: A Theoretical Explanation for the Movement of Vesicles across the Endothelium. Nature 1966, 212, 353–355. [Google Scholar] [CrossRef]
- Azimi, M.; Jamali, Y.; Mofrad, M.R.K. Accounting for Diffusion in Agent Based Models of Reaction-Diffusion Systems with Application to Cytoskeletal Diffusion. PLoS ONE 2011, 6, e25306. [Google Scholar] [CrossRef]
- Robertson, R.M.; Laib, S.; Smith, D.E. Diffusion of isolated DNA molecules: Dependence on length and topology. Proc. Natl. Acad. Sci. USA 2006, 103, 7310–7314. [Google Scholar] [CrossRef] [PubMed]
- Werner, A. Predicting translational diffusion of evolutionary conserved RNA structures by the nucleotide number. Nucleic Acids Res. 2010, 39, e17. [Google Scholar] [CrossRef] [PubMed]
- Kristoffersen, E.L.; Burman, M.; Noy, A.; Holliger, P. Rolling circle RNA synthesis catalyzed by RNA. eLife 2022, 11, e75186. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Term | Definition | Value |
|---|---|---|
| Number of circular ssRNA at site | variable | |
| Number of circular dsRNA at site | variable | |
| Number of open-ended ssRNA at site | variable | |
| Number of replicase at site | variable | |
| Number of cyclase at site | variable | |
| Number of nucleotide-synthase at site | variable | |
| Number of peptidyl-transferase at site | variable | |
| Replicase-catalyzed replication rate, cyclase-catalyzed new circular ssRNA creation rate | ||
| Non-enzymatic replication rate of template (circular ssRNA or dsRNA) at site | variable | |
| V | Reference volume of each lattice site | 100 |
| Monomer dependent rate reduction factor at site | ||
| Instantaneous number of monomer at site in unit of 200 mers | variable | |
| b | Number of monomers created by a nucleotide- synthase in unit of 200 mers | 1 |
| Maximum number of monomer at site in unit of 200 mers | 80 | |
| h | Degradation rate of each strand | |
| Replicase, Cyclase, Nucleotide-synthase and Peptidyl-transferase creation probabilities respectively | 0.03, 0.03, 0.03, 0.03 | |
| Threshold volume for division of vesicle created from site | ||
| Q | Strength of a peptidyl-transferase in increasing the threshold volume for division | 20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roy, S.; Sengupta, S. The Effect of Environment on the Evolution and Proliferation of Protocells of Increasing Complexity. Life 2022, 12, 1227. https://doi.org/10.3390/life12081227
Roy S, Sengupta S. The Effect of Environment on the Evolution and Proliferation of Protocells of Increasing Complexity. Life. 2022; 12(8):1227. https://doi.org/10.3390/life12081227
Chicago/Turabian StyleRoy, Suvam, and Supratim Sengupta. 2022. "The Effect of Environment on the Evolution and Proliferation of Protocells of Increasing Complexity" Life 12, no. 8: 1227. https://doi.org/10.3390/life12081227
APA StyleRoy, S., & Sengupta, S. (2022). The Effect of Environment on the Evolution and Proliferation of Protocells of Increasing Complexity. Life, 12(8), 1227. https://doi.org/10.3390/life12081227