
Ecdysteroids as Potent Enzyme Inhibitors and Verification of Their Activity Using In Vitro and In Silico Docking Studies

 ,
,  ,
,  ,
,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection of the Tested Ecdysteroids
2.2. Determination of the Enzyme Inhibitory Potential of the Selected Ecdysteroids by In Vitro Assays
2.2.1. Cholinesterase (ChE) Inhibitory Activity
2.2.2. Tyrosinase Inhibitory Activity
2.2.3. α-Amylase Inhibitory Activity
2.2.4. Statistical Analysis
2.3. Molecular Modelling Study
2.3.1. Preparation of the Receptor
2.3.2. Preparation of the Ligands
2.3.3. Molecular Docking
2.4. Evaluation of ADMET/TOPKAT Properties
2.5. Multivariate Data Analysis
3. Results and Discussion
3.1. Ecdysteroids Selected in this Study
3.2. Determination of the Enzyme Inhibitory Potential of the Ecdysteroids by In Vitro Assays
3.2.1. Cholinesterase (ChE) Inhibitory Activity
3.2.2. Tyrosinase Inhibitory Activity
3.2.3. α-Amylase Inhibitory Activity
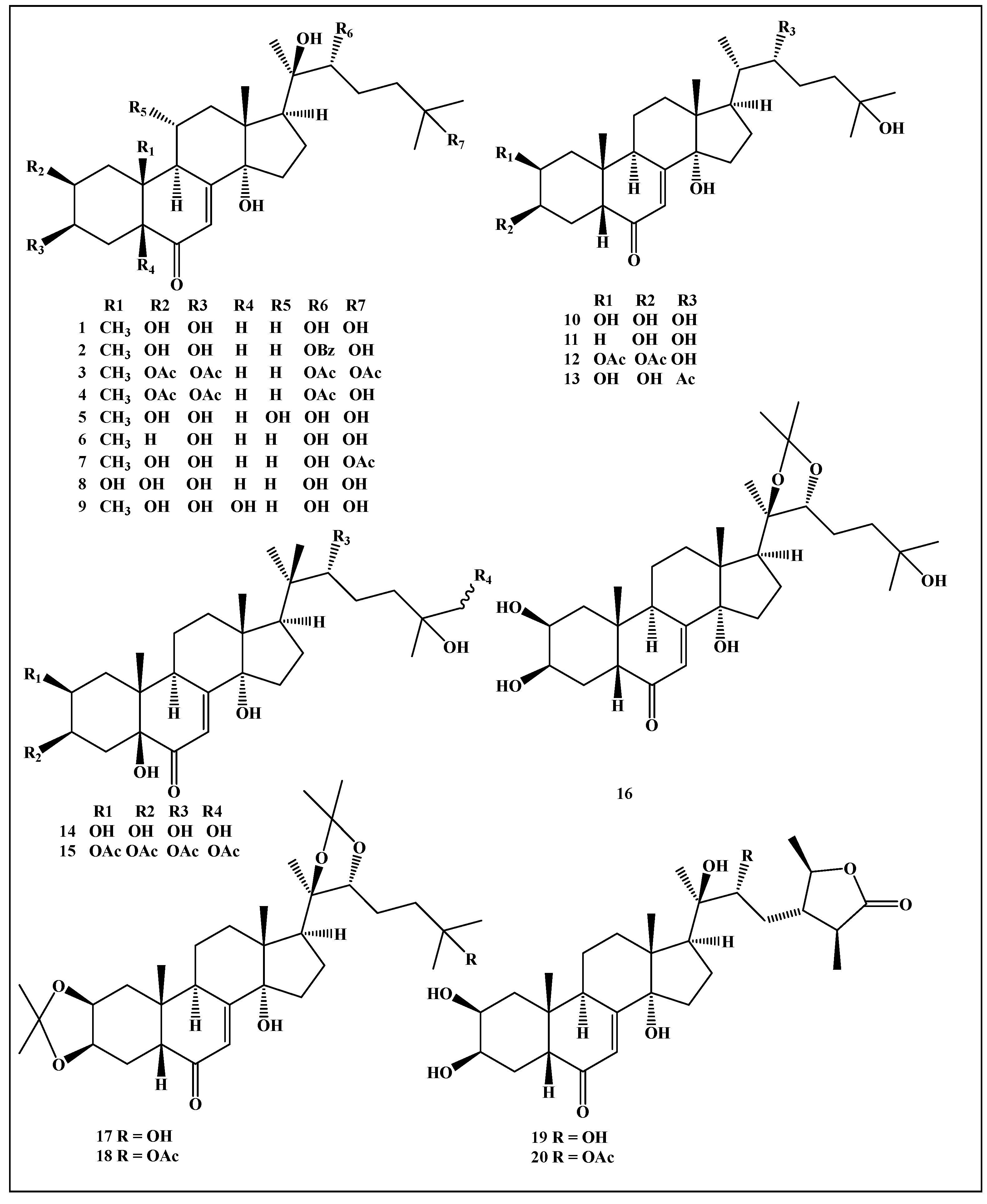
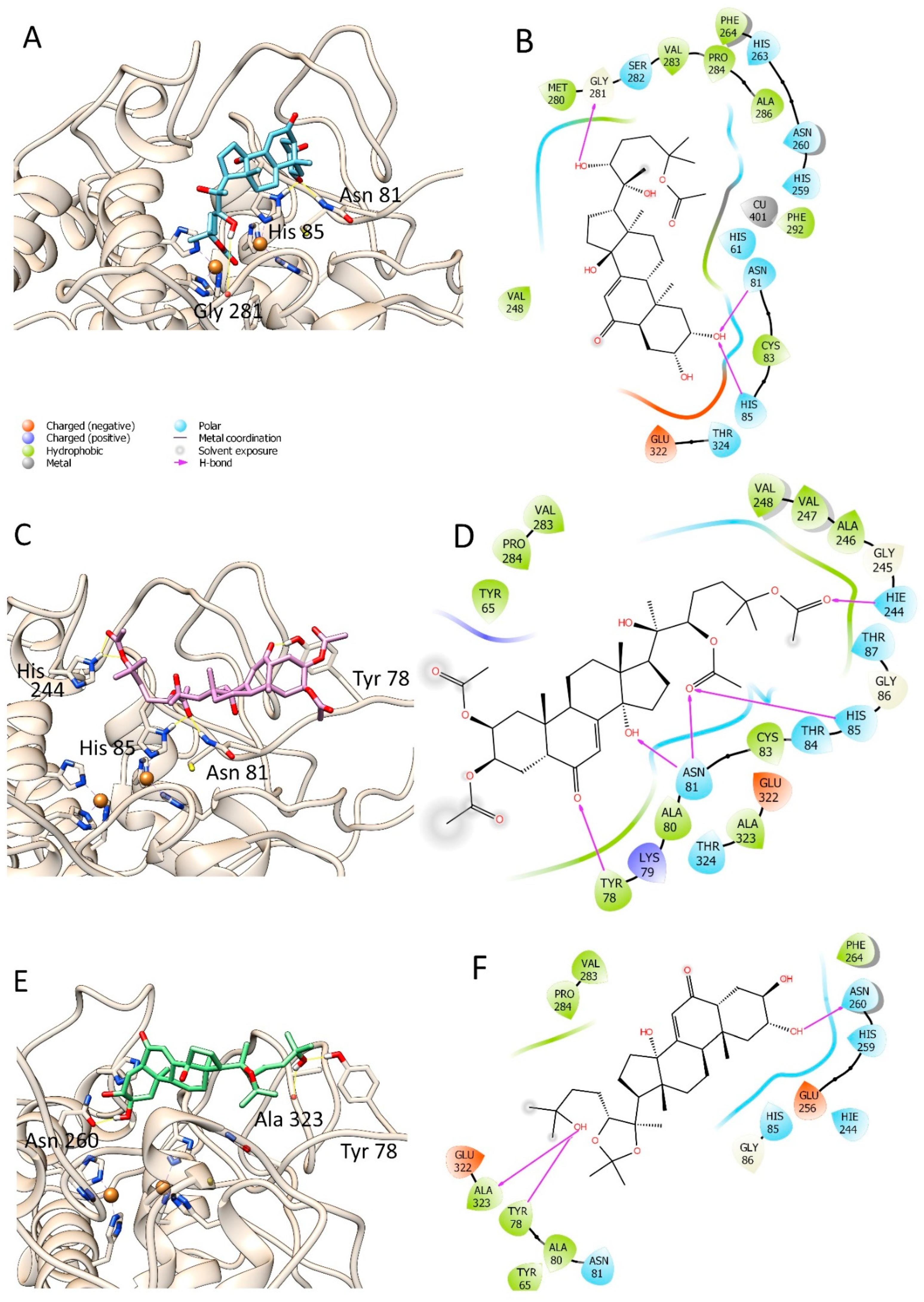
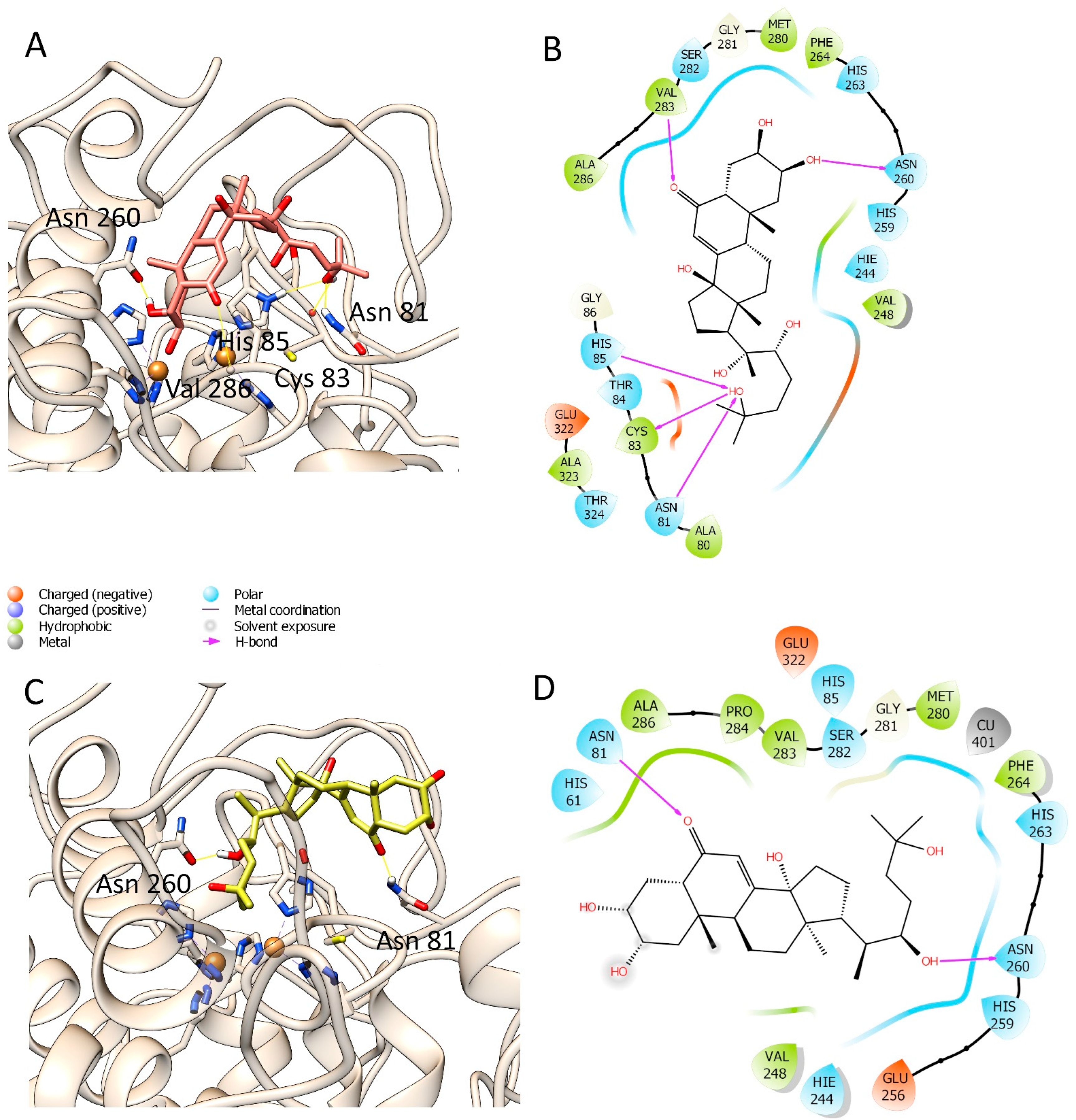
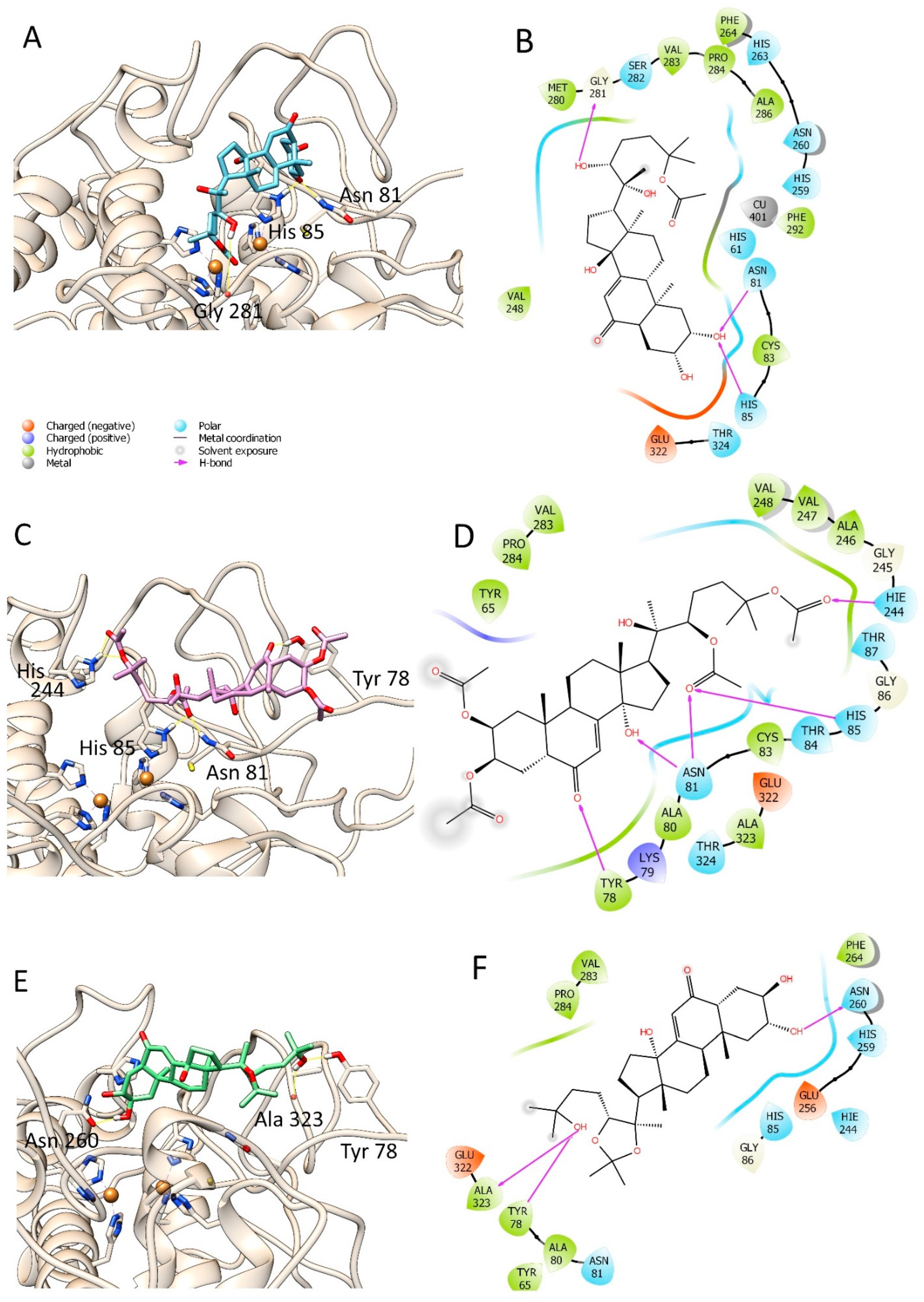
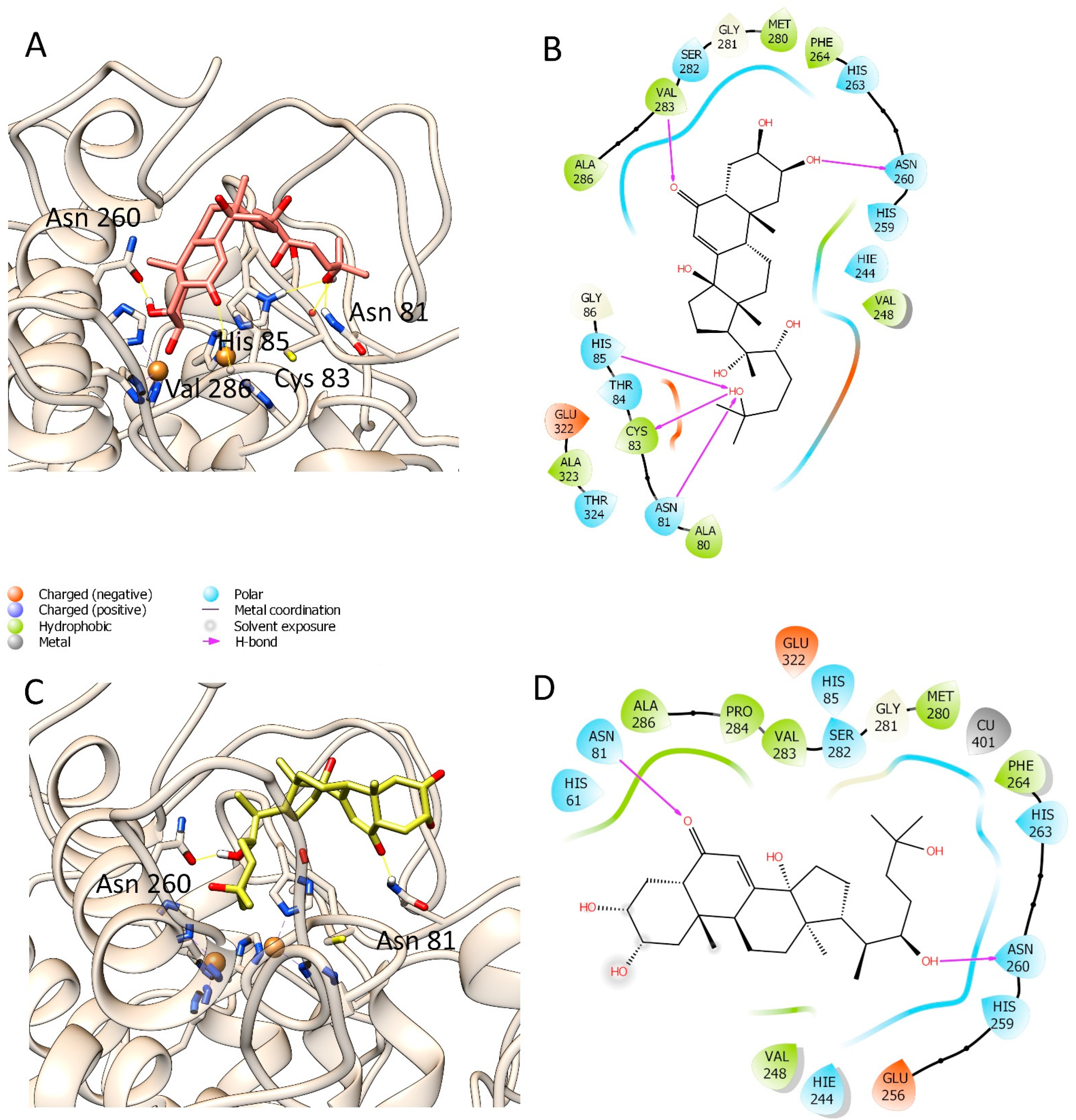
3.3. Molecular Modelling Studies
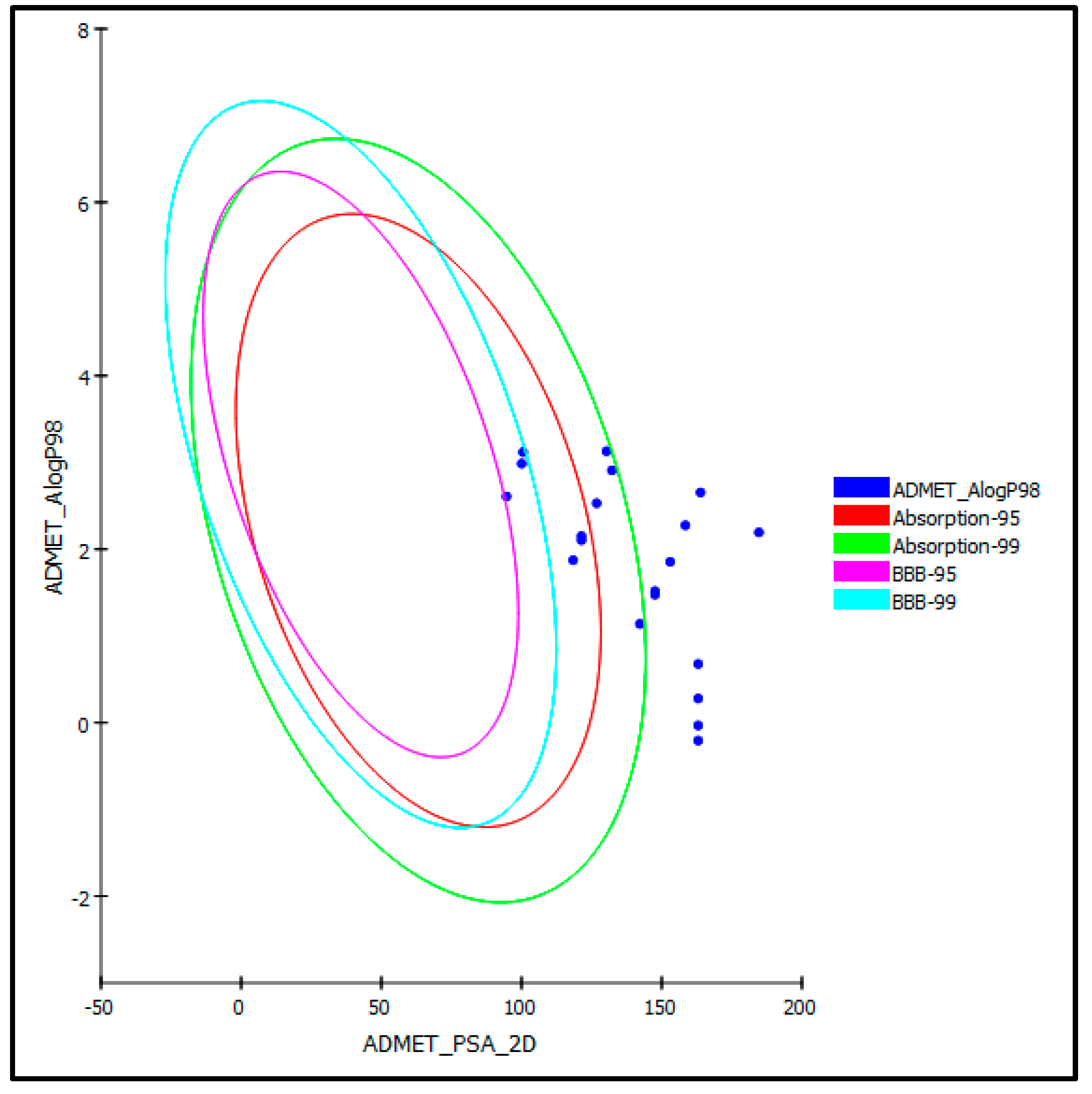
3.4. Evaluation of ADMET/TOPKAT Properties
3.5. Multivariate Data Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the last 25 years. J. Nat. Prod. 2007, 70, 461–477. [Google Scholar] [CrossRef] [Green Version]
- Wink, M. Current understanding of modes of action of multicomponent bioactive phytochemicals: Potential for nutraceuticals and antimicrobials. Annu. Rev. Food Sci. Technol. 2022, 13, 337–359. [Google Scholar] [CrossRef]
- Blow, D.M. Structure and mechanism of chymotrypsin. Acc. Chem. Res. 1976, 9, 145–152. [Google Scholar] [CrossRef]
- Huang, Y.; Jiang, J.; Wang, Y.; Chen, J.; Xi, J. Nanozymes as enzyme inhibitors. Int. J. Nanomed. 2021, 16, 1143–1155. [Google Scholar] [CrossRef]
- Birks, J.S. Cholinesterase inhibitors for Alzheimer’s disease. Cochrane Database Syst. Rev. 2006, CD005593. [Google Scholar] [CrossRef]
- Chang, T.-S. An updated review of tyrosinase inhibitors. Int. J. Mol. Sci. 2009, 10, 2440–2475. [Google Scholar] [CrossRef] [Green Version]
- Dirir, A.M.; Daou, M.; Yousef, A.F.; Yousef, L.F. A review of alpha-glucosidase inhibitors from plants as potential candidates for the treatment of type-2 diabetes. Phytochem. Rev. 2021, 1–31. [Google Scholar] [CrossRef]
- Lafont, R.; Koolman, J. Diversity of ecdysteroids in animal species. In Ecdysone: Structures and Functions; Springer: Dordrecht, The Netherlands, 2009; pp. 47–71. [Google Scholar]
- Dinan, L. Phytoecdysteroids: Biological aspects. Phytochemistry 2001, 57, 325–339. [Google Scholar] [CrossRef]
- Lafont, R.; Harmatha, J.; Marion-Poll, F.; Dinan, L.; Wilson, I. Ecdybase, a Free Ecdysteroid Database; Retrieved May; Cybersales: Prague, Czech Republic, 2002; Volume 26. [Google Scholar]
- Dinan, L.; Lafont, R. Effects and applications of arthropod steroid hormones (ecdysteroids) in mammals. J. Endocrinol. 2006, 191, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Aktumsek, A.; Zengin, G.; Guler, G.O.; Cakmak, Y.S.; Duran, A. Antioxidant potentials and anticholinesterase activities of methanolic and aqueous extracts of three endemic Centaurea L. species. Food Chem. Toxicol. 2013, 55, 290–296. [Google Scholar] [CrossRef]
- Mamadalieva, N.Z.; Youssef, F.S.; Hussain, H.; Zengin, G.; Mollica, A.; Al Musayeib, N.M.; Ashour, M.L.; Westermann, B.; Wessjohann, L.A. Validation of the antioxidant and enzyme inhibitory potential of selected triterpenes using in vitro and in silico studies, and the evaluation of their ADMET properties. Molecules 2021, 26, 6331. [Google Scholar] [CrossRef]
- Zengin, G. A study on in vitro enzyme inhibitory properties of Asphodeline anatolica: New sources of natural inhibitors for public health problems. Ind. Crop. Prod. 2016, 83, 39–43. [Google Scholar] [CrossRef]
- Zengin, G.; Sarikurkcu, C.; Aktumsek, A.; Ceylan, R.; Ceylan, O. A comprehensive study on phytochemical characterization of Haplophyllum myrtifolium Boiss endemic to Turkey and its inhibitory potential against key enzymes involved in Alzheimer, skin diseases and type II diabetes. Ind. Crop. Prod. 2014, 53, 244–251. [Google Scholar] [CrossRef]
- Lazarova, I.; Zengin, G.; Bender, O.; Zheleva-Dimitrova, D.; Uysal, S.; Ceylan, R.; Gevrenova, R.; Aktumsek, A.; Acar, M.; Gunduz, M. A comparative study of Bulgarian and Turkish Asphodeline lutea root extracts: HPLC–UV profiles, enzyme inhibitory potentials and anti-proliferative activities against MCF-7 and MCF-10A cell lines. J. Funct. Foods 2015, 15, 254–263. [Google Scholar] [CrossRef]
- Ismaya, W.T.; Rozeboom, H.J.; Weijn, A.; Mes, J.J.; Fusetti, F.; Wichers, H.J.; Dijkstra, B.W. Crystal structure of Agaricus bisporus mushroom tyrosinase: Identity of the tetramer subunits and interaction with tropolone. Biochemistry 2011, 50, 5477–5486. [Google Scholar] [CrossRef] [Green Version]
- Sinan, K.I.; Dall’Acqua, S.; Ferrarese, I.; Mollica, A.; Stefanucci, A.; Glamočlija, J.; Sokovic, M.; Nenadić, M.; Aktumsek, A.; Zengin, G. LC-MS based analysis and biological properties of Pseudocedrela kotschyi (Schweinf.) Harms extracts: A valuable source of antioxidant, antifungal, and antibacterial compounds. Antioxidants 2021, 10, 1570. [Google Scholar] [CrossRef]
- Harder, E.; Damm, W.; Maple, J.; Wu, C.; Reboul, M.; Xiang, J.Y.; Wang, L.; Lupyan, D.; Dahlgren, M.K.; Knight, J.L.; et al. OPLS3: A force field providing broad coverage of drug-like small molecules and proteins. J. Chem. Theory Comput. 2016, 12, 281–296. [Google Scholar] [CrossRef]
- Schrödinger Release 2017-1; Maestro, Schrödinger, LLC: New York, NY, USA, 2017.
- Zengin, G.; Senkardes, I.; Mollica, A.; Picot-Allain, C.M.N.; Bulut, G.; Dogan, A.; Mahomoodally, M.F. New insights into the in vitro biological effects, in silico docking and chemical profile of clary sage—Salvia sclarea L. Comput. Biol. Chem. 2018, 75, 111–119. [Google Scholar] [CrossRef]
- Youssef, F.S.; Ovidi, E.; Musayeib, N.M.A.; Ashour, M.L. Morphology, anatomy and secondary metabolites investigations of Premna odorata Blanco and evaluation of its anti-tuberculosis activity using in vitro and in silico studies. Plants 2021, 10, 1953. [Google Scholar] [CrossRef]
- Elhady, S.S.; Youssef, F.S.; Alahdal, A.M.; Almasri, D.M.; Ashour, M.L. Anti-hyperglycaemic evaluation of Buddleia indica leaves using in vitro, in vivo and in silico studies and its correlation with the major phytoconstituents. Plants 2021, 10, 2351. [Google Scholar] [CrossRef]
- Altyar, A.E.; Ashour, M.L.; Youssef, F.S. Premna odorata: Seasonal metabolic variation in the essential oil composition of its leaf and verification of its anti-ageing potential via in vitro assays and molecular modelling. Biomolecules 2020, 10, 879. [Google Scholar] [CrossRef] [PubMed]
- Youssef, F.S.; Mamatkhanova, M.A.; Mamadalieva, N.Z.; Zengin, G.; Aripova, S.F.; Alshammari, E.; Ashour, M.L. Chemical profiling and discrimination of essential oils from six Ferula species using GC analyses coupled with chemometrics and evaluation of their antioxidant and enzyme inhibitory potential. Antibiotics 2020, 9, 518. [Google Scholar] [CrossRef] [PubMed]
- Mamadalieva, N.Z.; Böhmdorfer, S.; Zengin, G.; Bacher, M.; Potthast, A.; Akramov, D.K.; Janibekov, A.; Rosenau, T. Phytochemical and biological activities of Silene viridiflora extractives. Development and validation of a HPTLC method for quantification of 20-hydroxyecdysone. Ind. Crop. Prod. 2019, 129, 542–548. [Google Scholar] [CrossRef]
- Mishra, P.; Kumar, A.; Panda, G. Anti-cholinesterase hybrids as multi-target-directed ligands against Alzheimer’s disease (1998–2018). Bioorg. Med. Chem. 2019, 27, 895–930. [Google Scholar] [CrossRef] [PubMed]
- Ben Nejma, A.; Nguir, A.; Ben Jannet, H.; Hamza, M.A.; Daich, A.; Othman, M.; Lawson, A.M. New septanoside and 20-hydroxyecdysone septanoside derivative from Atriplex portulacoides roots with preliminary biological activities. Bioorg. Med. Chem. Lett. 2015, 25, 1665–1670. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Biswas, R.; Sharma, A.; Banerjee, S.; Biswas, S.; Katiyar, C. Validation of medicinal herbs for anti-tyrosinase potential. J. Herb. Med. 2018, 14, 1–16. [Google Scholar] [CrossRef]
- Li, X.; Bai, Y.; Jin, Z.; Svensson, B. Food-derived non-phenolic α-amylase and α-glucosidase inhibitors for controlling starch digestion rate and guiding diabetes-friendly recipes. LWT 2022, 153, 112455. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Tyrosinase Inhibitory Potential | α-Amylase Inhibitory Activity |
|---|---|---|
| 1 | 19.65 ± 2.78 ef | 0.10 ± 0.03 def |
| 2 | 60.50 ± 4.69 bc | 0.16 ± 0.05 cd |
| 3 | 71.41 ± 5.79 ab | 0.17 ± 0.01 cd |
| 4 | 69.10 ± 4.89 ab | 0.14 ± 0.04 de |
| 5 | 10.67 ± 0.92 fg | 0.07 ± 0.01 ef |
| 6 | 20.34 ± 0.39 ef | 0.10 ± 0.03 def |
| 7 | 79.88 ± 1.36 a | 0.14 ± 0.01 de |
| 8 | 37.38 ± 0.09 d | 0.11 ± 0.02 def |
| 9 | 62.49 ± 5.77 bc | 0.14 ± 0.01 de |
| 10 | 13.30 ± 1.10 fg | 0.07 ± 0.01 ef |
| 11 | 6.36 ± 0.20 g | 0.05 ± 0.01 f |
| 12 | 64.45 ± 4.87 bc | 0.24 ± 0.03 bc |
| 13 | 53.19 ± 2.64 c | 0.16 ± 0.02 cd |
| 14 | 54.19 ± 4.86 c | 0.17 ± 0.04 cd |
| 15 | 31.51 ± 2.60 de | 0.15 ± 0.02 de |
| 16 | 69.33 ± 3.04 ab | 0.32 ± 0.01 ab |
| 17 | 61.59 ± 9.30 bc | 0.11 ± 0.01 def |
| 18 | 62.94 ± 7.44 bc | 0.35 ± 0.05 a |
| 19 | 18.60 ± 3.29 efg | 0.10 ± 0.03 def |
| 20 | 11.74 ± 1.49 fg | 0.09 ± 0.01 def |
| Compound | XP Docking Score | ΔG Free Binding Energy |
|---|---|---|
| 3 | −5.451 | −31.91 |
| 4 | −5.336 | −14.26 |
| 7 | −5.716 | −29.26 |
| 12 | −4.840 | −20.67 |
| 16 | −5.375 | −24.75 |
| Compounds | Absorption Level | Solubility Level | BBB Level | PPB Level | CPY2D6 | Hepato-Toxic | Alog p98 | PSA-2D |
|---|---|---|---|---|---|---|---|---|
| 1 | 1 | 3 | 4 | False | NI | NT | 1.14 | 142.193 |
| 2 | 1 | 3 | 4 | False | NI | NT | 2.09 | 130.308 |
| 3 | 3 | 3 | 4 | False | NI | NT | 2.65 | 163.855 |
| 4 | 3 | 3 | 4 | False | NI | NT | 2.28 | 158.440 |
| 5 | 3 | 4 | 4 | False | NI | TOX | −0.03 | 163.009 |
| 6 | 0 | 3 | 4 | False | NI | NT | 2.10 | 121.378 |
| 7 | 2 | 3 | 4 | False | NI | NT | 1.52 | 147.609 |
| 8 | 3 | 4 | 4 | False | NI | TOX | −0.21 | 163.009 |
| 9 | 3 | 4 | 4 | False | NI | NT | 0.28 | 163.009 |
| 10 | 0 | 3 | 4 | False | NI | NT | 2.15 | 121.378 |
| 11 | 0 | 3 | 3 | False | NI | NT | 3.12 | 100.562 |
| 12 | 1 | 3 | 4 | False | NI | NT | 2.91 | 132.209 |
| 13 | 1 | 3 | 4 | False | NI | NT | 2.53 | 126.793 |
| 14 | 3 | 4 | 4 | False | NI | NT | 0.68 | 163.009 |
| 15 | 3 | 3 | 4 | False | NI | NT | 2.19 | 184.67 |
| 16 | 0 | 3 | 4 | False | NI | NT | 1.87 | 118.422 |
| 17 | 0 | 2 | 3 | False | NI | NT | 2.61 | 94.652 |
| 18 | 0 | 2 | 3 | False | NI | NT | 2.99 | 100.067 |
| 19 | 2 | 3 | 4 | False | NI | NT | 1.48 | 147.609 |
| 20 | 3 | 3 | 4 | False | NI | NT | 1.86 | 153.024 |
| Compound | Ames Prediction | Rat Oral LD50 | Rat Chronic LOAEL | Skin Irritancy | Ocular Irritancy | Rat Female FDA | Rat Male FDA |
|---|---|---|---|---|---|---|---|
| 1 | Non-Mutagen | 21.76 | 0.02 | Moderate | Severe | Carcinogen | Carcinogen |
| 2 | Non-Mutagen | 17.04 | 0.03 | Mild | None | Non-Carcinogen | Non-Carcinogen |
| 3 | Non-Mutagen | 12.77 | 0.01 | Moderate | Mild | Carcinogen | Carcinogen |
| 4 | Non-Mutagen | 17.30 | 0.01 | Moderate | Severe | Carcinogen | Carcinogen |
| 5 | Non-Mutagen | 29.14 | 0.03 | Mild | Severe | Carcinogen | Non-Carcinogen |
| 6 | Non-Mutagen | 16.58 | 0.02 | Moderate | Severe | Carcinogen | Non-Carcinogen |
| 7 | Non-Mutagen | 24.02 | 0.02 | Moderate | Moderate | Carcinogen | Carcinogen |
| 8 | Non-Mutagen | 21.25 | 0.03 | Mild | Severe | Carcinogen | Carcinogen |
| 9 | Non-Mutagen | 13.00 | 0.03 | Moderate | Severe | Carcinogen | Non-Carcinogen |
| 10 | Non-Mutagen | 15.44 | 0.02 | Moderate | Severe | Carcinogen | Non-Carcinogen |
| 11 | Non-Mutagen | 11.95 | 0.01 | Moderate | Severe | Carcinogen | Non-Carcinogen |
| 12 | Non-Mutagen | 16.58 | 0.01 | Moderate | Severe | Carcinogen | Carcinogen |
| 13 | Non-Mutagen | 17.09 | 0.01 | Moderate | Moderate | Carcinogen | Non-Carcinogen |
| 14 | Non-Mutagen | 9.75 | 0.03 | Moderate | Severe | Carcinogen | Non-Carcinogen |
| 15 | Non-Mutagen | 6.83 | 0.01 | Moderate | None | Carcinogen | Non-Carcinogen |
| 16 | Non-Mutagen | 7.55 | 0.004 | Moderate | Severe | Carcinogen | Non-Carcinogen |
| 17 | Non-Mutagen | 4.92 | 0.001 | Mild | Severe | Carcinogen | Non-Carcinogen |
| 18 | Non-Mutagen | 4.11 | 0.001 | Moderate | Moderate | Carcinogen | Carcinogen |
| 19 | Non-Mutagen | 8.60 | 0.006 | Moderate | Moderate | Carcinogen | Carcinogen |
| 20 | Non-Mutagen | 6.83 | 0.01 | Moderate | Moderate | Carcinogen | Non-Carcinogen |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mamadalieva, N.Z.; Hussain, H.; Mollica, A.; Zengin, G.; Mamadalieva, R.Z.; Elhady, S.S.; Fadil, S.A.; Ashour, M.L.; Youssef, F.S. Ecdysteroids as Potent Enzyme Inhibitors and Verification of Their Activity Using In Vitro and In Silico Docking Studies. Life 2022, 12, 824. https://doi.org/10.3390/life12060824
Mamadalieva NZ, Hussain H, Mollica A, Zengin G, Mamadalieva RZ, Elhady SS, Fadil SA, Ashour ML, Youssef FS. Ecdysteroids as Potent Enzyme Inhibitors and Verification of Their Activity Using In Vitro and In Silico Docking Studies. Life. 2022; 12(6):824. https://doi.org/10.3390/life12060824
Chicago/Turabian StyleMamadalieva, Nilufar Z., Hidayat Hussain, Adriano Mollica, Gokhan Zengin, Rano Z. Mamadalieva, Sameh S. Elhady, Sana A. Fadil, Mohamed L. Ashour, and Fadia S. Youssef. 2022. "Ecdysteroids as Potent Enzyme Inhibitors and Verification of Their Activity Using In Vitro and In Silico Docking Studies" Life 12, no. 6: 824. https://doi.org/10.3390/life12060824
APA StyleMamadalieva, N. Z., Hussain, H., Mollica, A., Zengin, G., Mamadalieva, R. Z., Elhady, S. S., Fadil, S. A., Ashour, M. L., & Youssef, F. S. (2022). Ecdysteroids as Potent Enzyme Inhibitors and Verification of Their Activity Using In Vitro and In Silico Docking Studies. Life, 12(6), 824. https://doi.org/10.3390/life12060824