
Scum of the Earth: A Hypothesis for Prebiotic Multi-Compartmentalised Environments


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
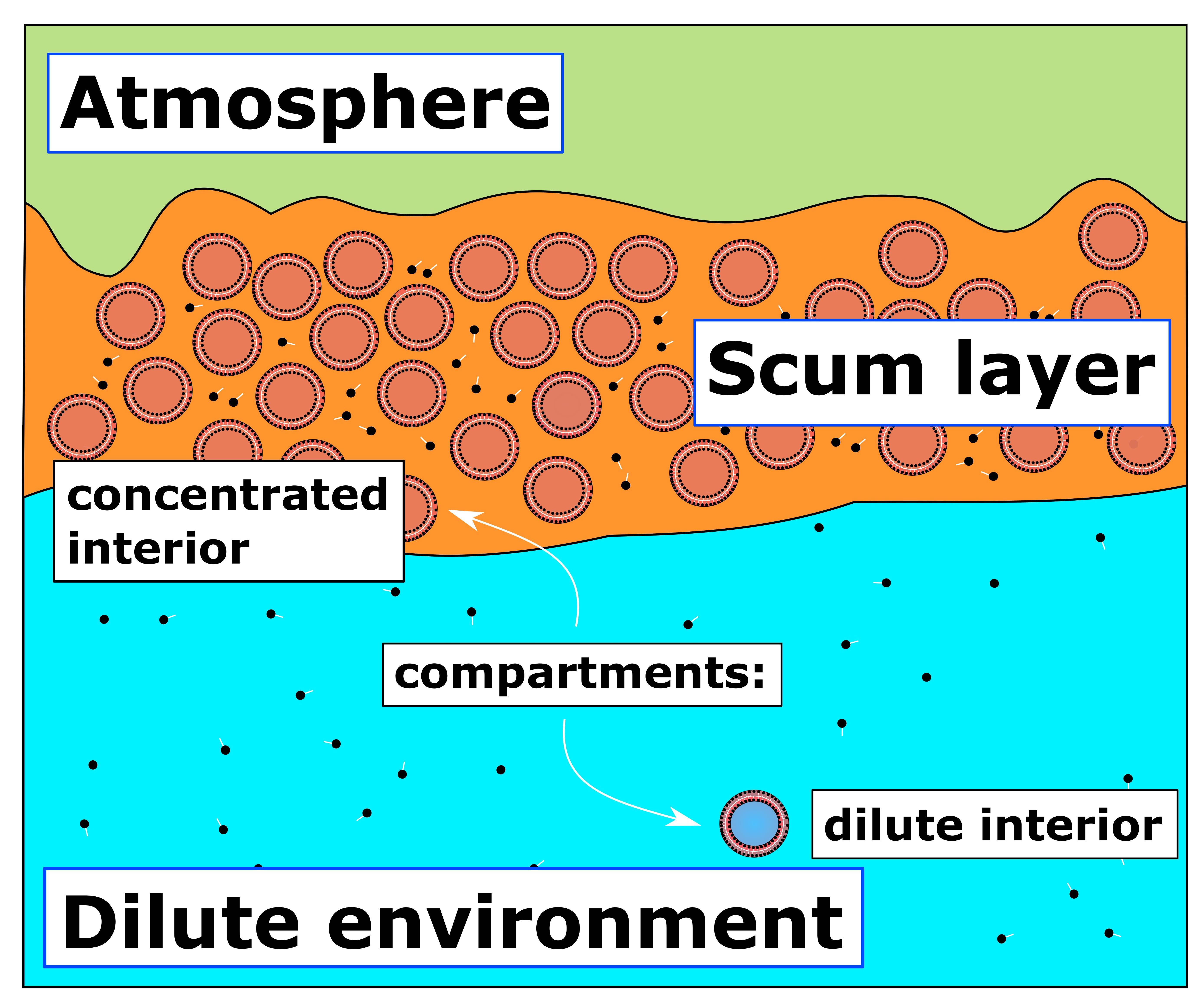
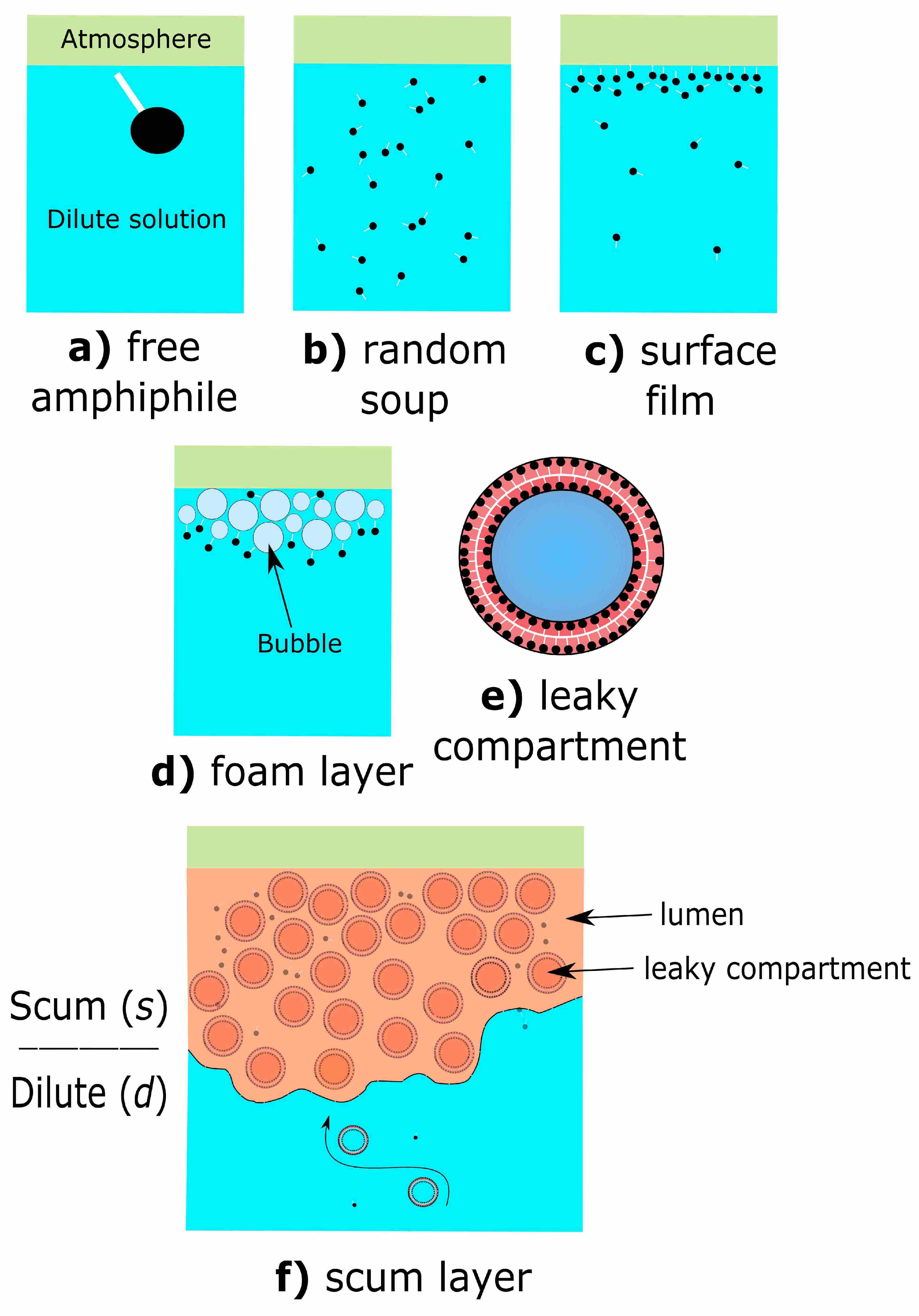
2. The Scum Hypothesis
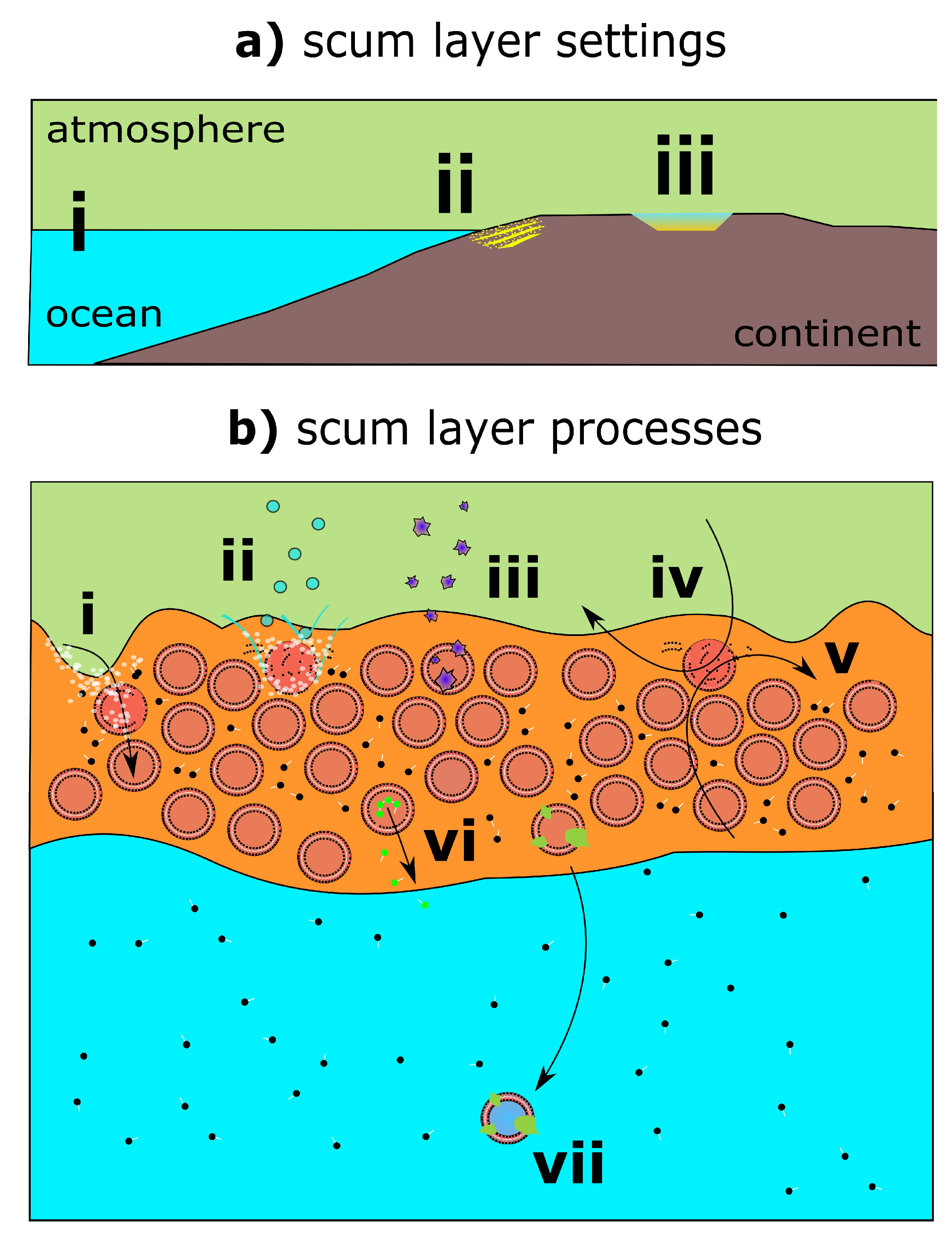
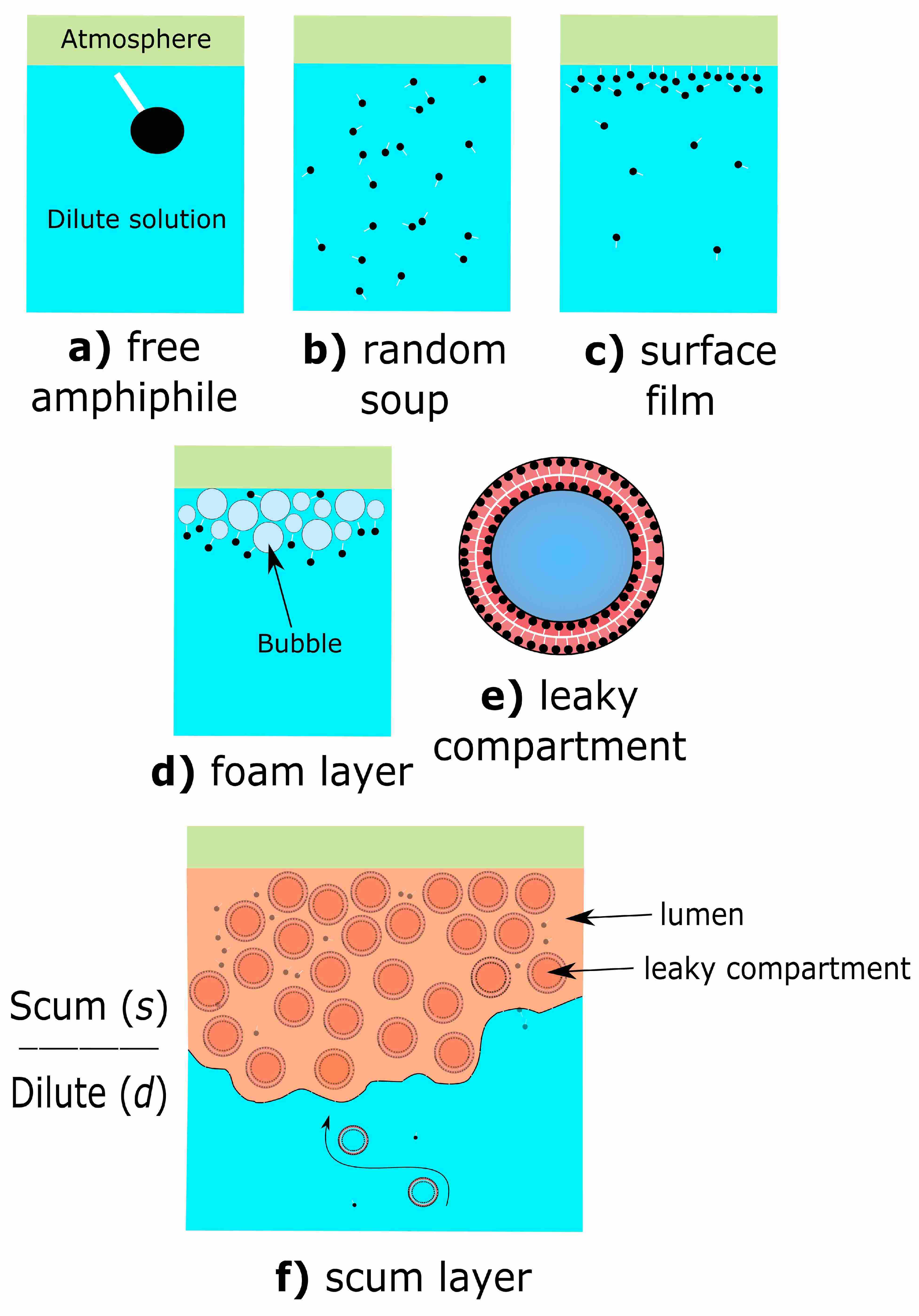
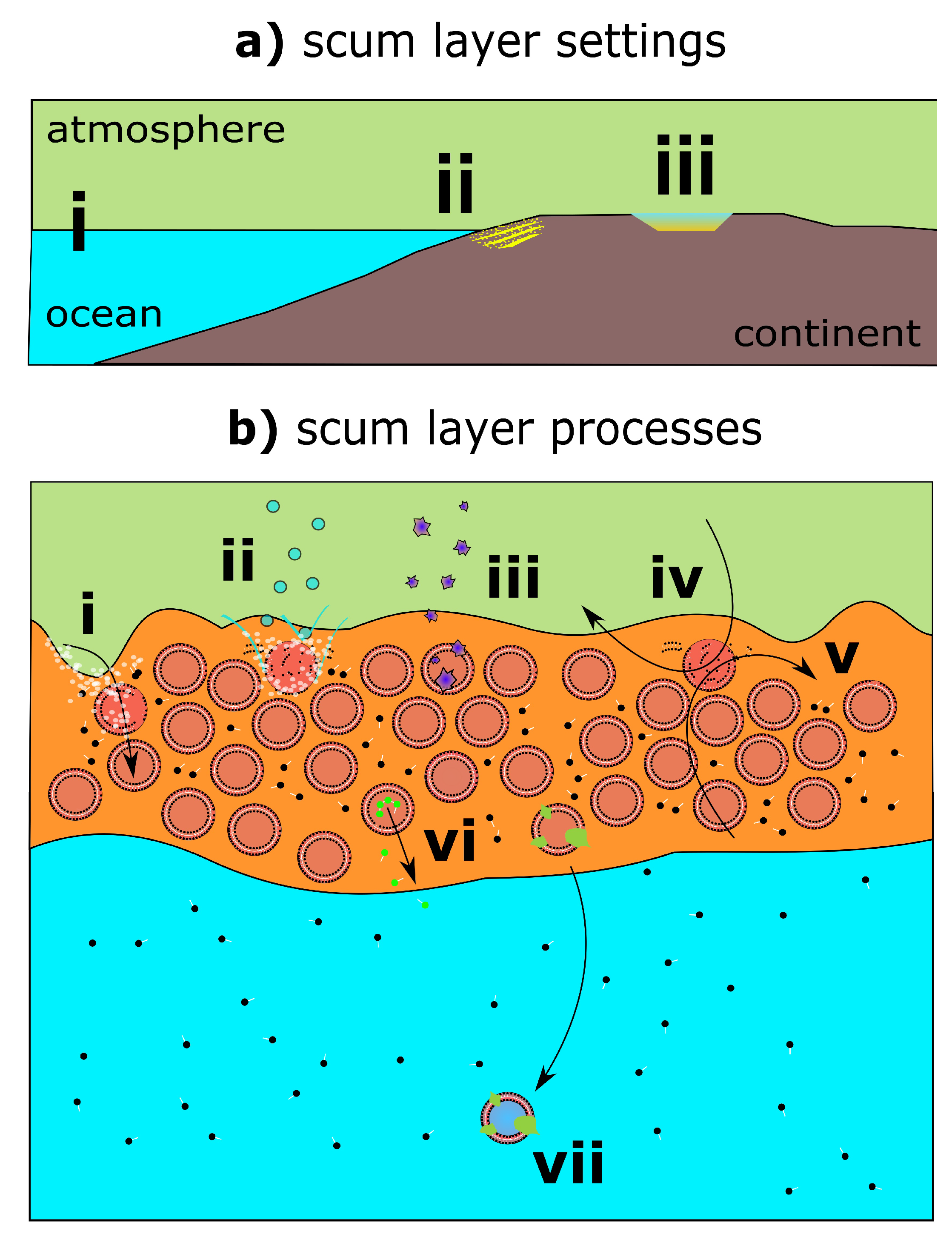
2.1. Prebiotic Plausibility of Scum Layers
2.2. Prebiotic Utility of Scum Layers
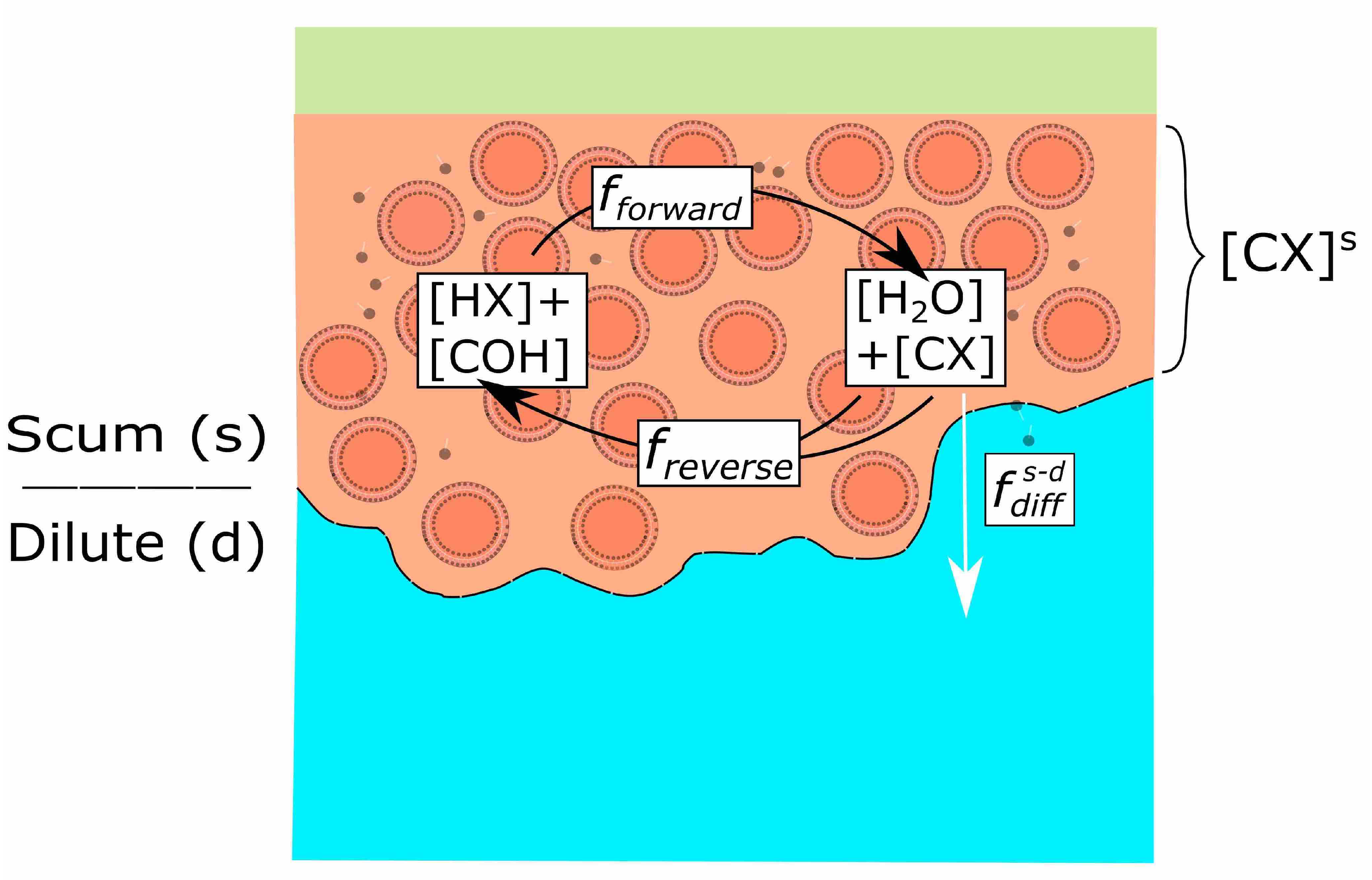
3. A Simple Model of Scum Layer Chemistry
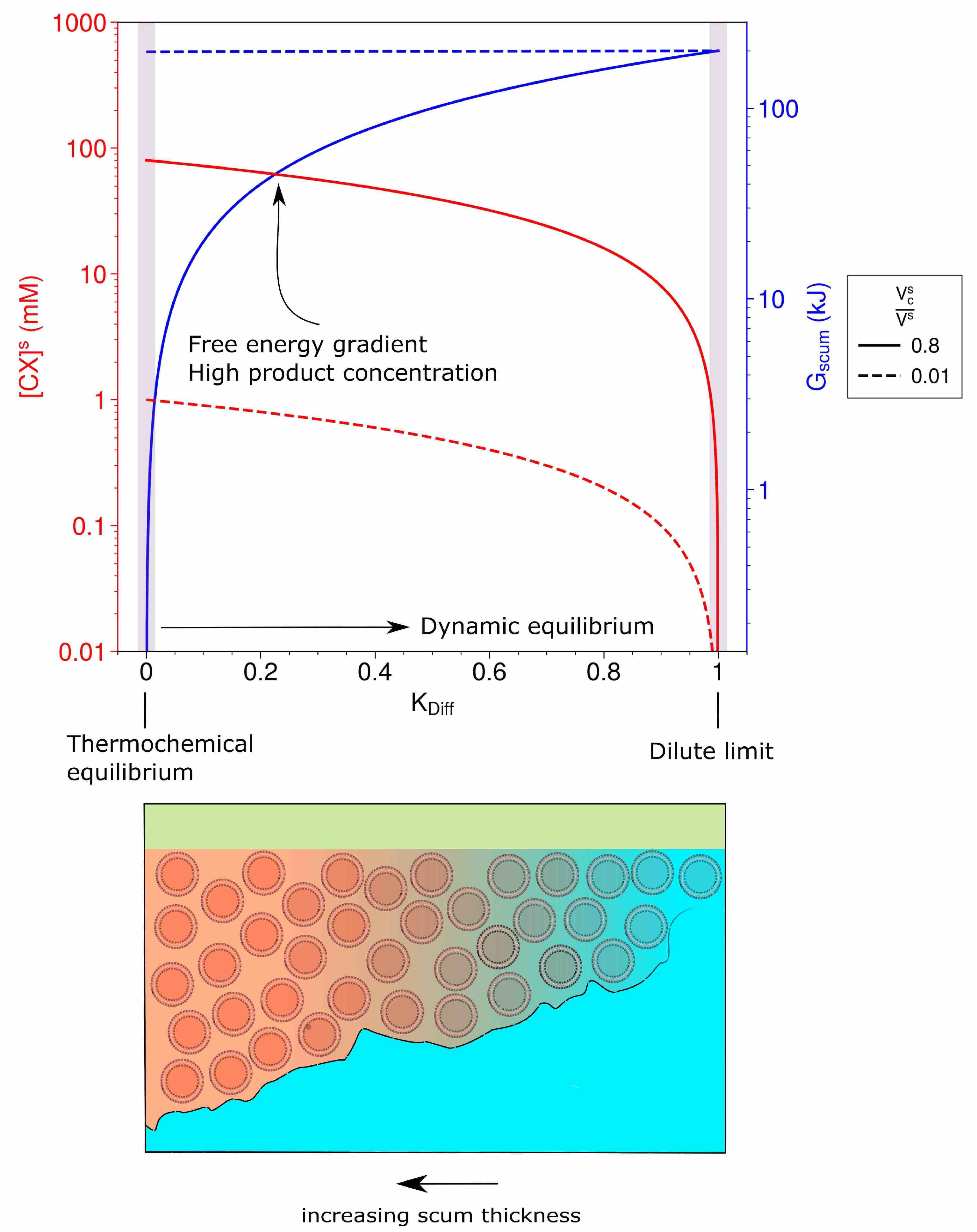
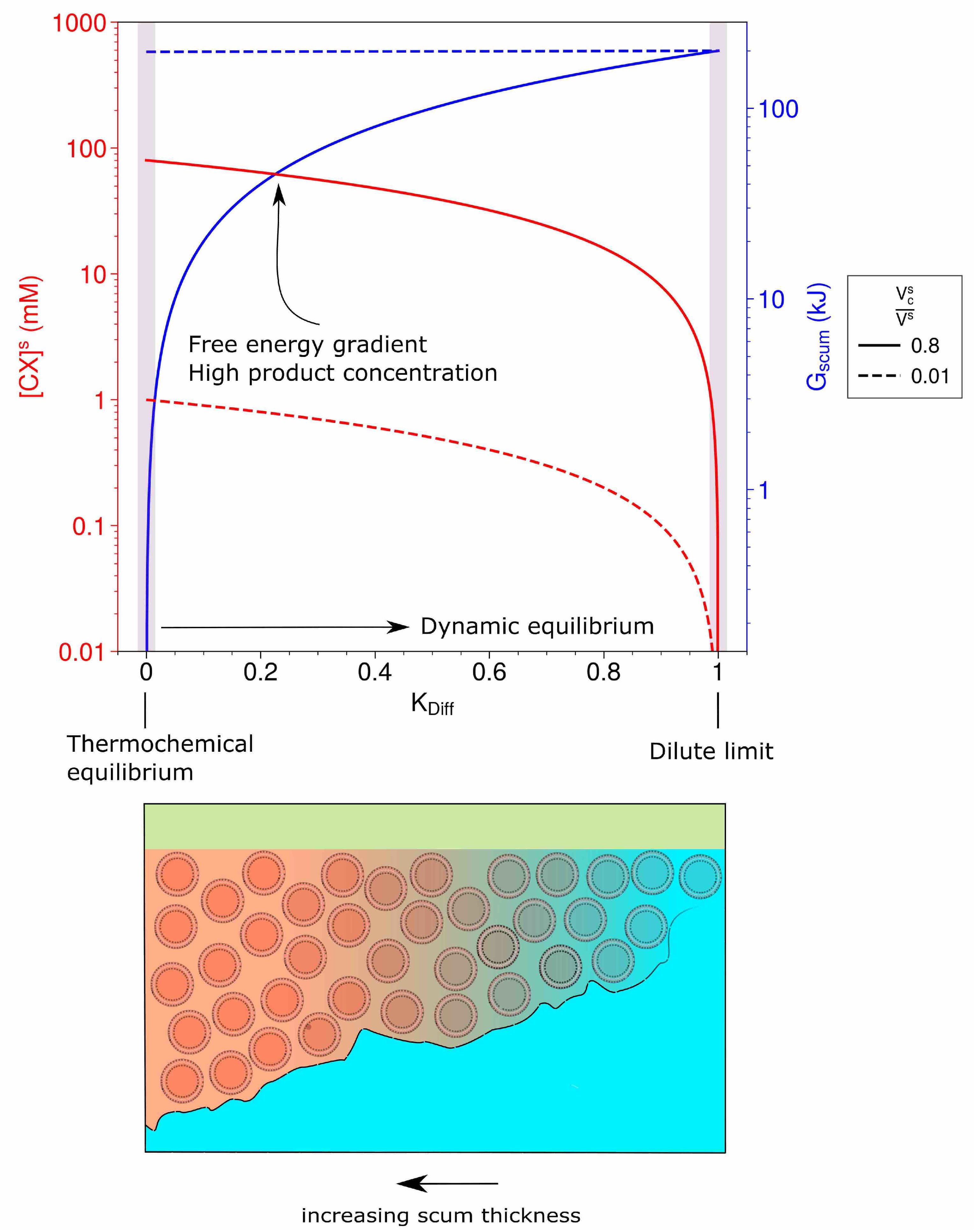
3.1. Dynamic Equilibrium: The Drive to Life
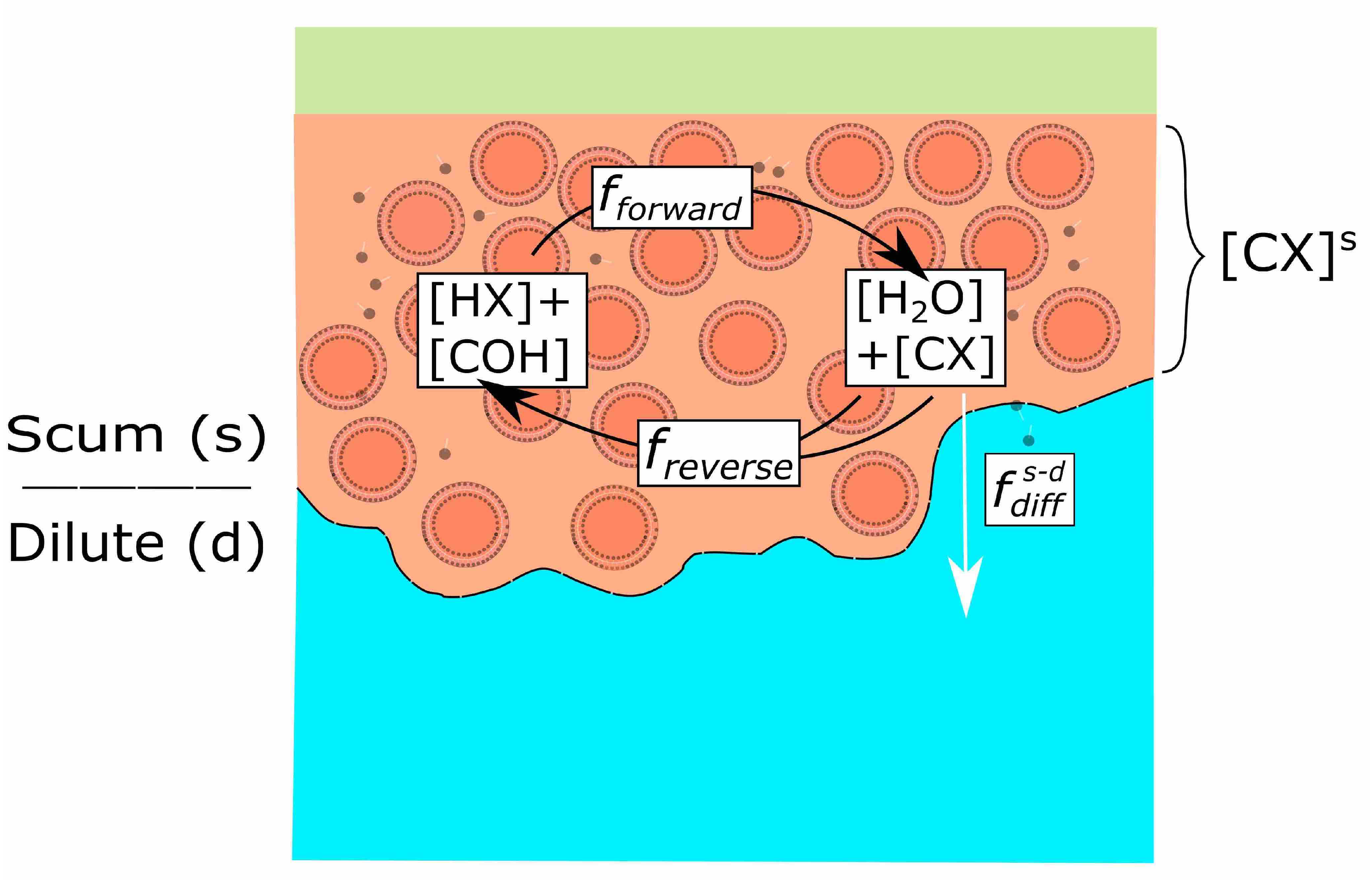
3.2. A Simple Model of Scum Layer Processes
4. Summary and Next Steps
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deal, A.M.; Rapf, R.J.; Vaida, V. Water–Air Interfaces as Environments to Address the Water Paradox in Prebiotic Chemistry: A Physical Chemistry Perspective. J. Phys. Chem. A 2021, 125, 4929–4942. [Google Scholar] [CrossRef]
- Crispino, G.; Ho, P.; Sharpless, K. Selective perhydroxylation of squalene: Taming the arithmetic demon. Science 1993, 259, 64–66. [Google Scholar] [CrossRef]
- Cartwright, J.H.E.; Russell, M.J. The origin of life: The submarine alkaline vent theory at 30. Interface Focus 2019, 9, 20190104. [Google Scholar] [CrossRef]
- Jia, T.Z.; Chandru, K.; Hongo, Y.; Afrin, R.; Usui, T.; Myojo, K.; Cleaves, H.J. Membraneless polyester microdroplets as primordial compartments at the origins of life. Proc. Natl. Acad. Sci. USA 2019, 116, 15830–15835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonfio, C.; Russell, D.A.; Green, N.J.; Mariani, A.; Sutherland, J.D. Activation chemistry drives the emergence of functionalised protocells. Chem. Sci. 2020, 11, 10688–10697. [Google Scholar] [CrossRef] [PubMed]
- Morasch, M.; Liu, J.; Dirscher, C.F.; Ianeselli, A.; Kühnlein, A.; Le Vay, K.; Schwintek, P.; Islam, S.; Corpinot, M.K.; Scheu, B.; et al. Heated gas bubbles enrich, crystallize, dry, phosphorylate and encapsulate prebiotic molecules. Nat. Chem. 2019. [Google Scholar] [CrossRef] [PubMed]
- Bonfio, C.; Godino, E.; Corsini, M.; Biani, F.F.d.; Guella, G.; Mansy, S.S. Prebiotic iron–sulfur peptide catalysts generate a pH gradient across model membranes of late protocells. Nat. Catal. 2018, 1, 616–623. [Google Scholar] [CrossRef]
- Deamer, D.; Damer, B.; Kompanichenko, V. Hydrothermal Chemistry and the Origin of Cellular Life. Astrobiology 2019, 19, 1523–1537. [Google Scholar] [CrossRef]
- Deamer, D. Liquid crystalline nanostructures: Organizing matrices for non-enzymatic nucleic acid polymerization. Chem. Soc. Rev. 2012, 41, 5375–5379. [Google Scholar] [CrossRef]
- Milshteyn, D.; Cooper, G.; Deamer, D. Chemiosmotic energy for primitive cellular life: Proton gradients are generated across lipid membranes by redox reactions coupled to meteoritic quinones. Sci. Rep. 2019, 9, 12447. [Google Scholar] [CrossRef]
- Milshteyn, D.; Damer, B.; Havig, J.; Deamer, D. Amphiphilic Compounds Assemble into Membranous Vesicles in Hydrothermal Hot Spring Water but Not in Seawater. Life 2018, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Rajamani, S.; Vlassov, A.; Benner, S.; Coombs, A.; Olasagasti, F.; Deamer, D. Lipid-assisted Synthesis of RNA-like Polymers from Mononucleotides. Orig. Life Evol. Biosph. 2008, 38, 57–74. [Google Scholar] [CrossRef]
- Wurl, O.; Wurl, E.; Miller, L.; Johnson, K.; Vagle, S. Formation and global distribution of sea-surface microlayers. Biogeosciences 2011, 8, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Walde, P. Surfactant Assemblies and their Various Possible Roles for the Origin(S) of Life. Orig. Life Evol. Biosph. 2006, 36, 109–150. [Google Scholar] [CrossRef]
- Engel, A.; Sperling, M.; Sun, C.; Grosse, J.; Friedrichs, G. Organic Matter in the Surface Microlayer: Insights From a Wind Wave Channel Experiment. Front. Mar. Sci. 2018, 5, 182. [Google Scholar] [CrossRef]
- Ćosović, B. Surface-Active Properties of the Sea Surface Microlayerand Consequences for Pollution in the Mediterranean Sea. In The Mediterranean Sea; Saliot, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 269–296. [Google Scholar] [CrossRef]
- Södergren, A. Role of aquatic surface microlayer in the dynamics of nutrients and organic compounds in lakes, with implications for their ecotones. Hydrobiologia 1993, 251, 217–225. [Google Scholar] [CrossRef]
- Yanagawa, H.; Ogawa, Y.; Kojima, K.; Ito, M. Construction of protocellular structures under simulated primitive earth conditions. Orig. Life Evol. Biosph. 1988, 18, 179–207. [Google Scholar] [CrossRef]
- Clark, B.C.; Kolb, V.M. Comet Pond II: Synergistic Intersection of Concentrated Extraterrestrial Materials and Planetary Environments to Form Procreative Darwinian Ponds. Life 2018, 8, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawless, J.G.; Yuen, G.U. Quantification of monocarboxylic acids in the Murchison carbonaceous meteorite. Nature 1979, 282, 396–398. [Google Scholar] [CrossRef]
- Liu, Z.; Wu, L.F.; Xu, J.; Bonfio, C.; Russell, D.A.; Sutherland, J.D. Harnessing chemical energy for the activation and joining of prebiotic building blocks. Nat. Chem. 2020, 12, 1023–1028. [Google Scholar] [CrossRef] [PubMed]
- Jordan, S.F.; Rammu, H.; Zheludev, I.N.; Hartley, A.M.; Maréchal, A.; Lane, N. Promotion of protocell self-assembly from mixed amphiphiles at the origin of life. Nat. Ecol. Evol. 2019, 3, 1705–1714. [Google Scholar] [CrossRef]
- Damer, B.; Deamer, D. Coupled Phases and Combinatorial Selection in Fluctuating Hydrothermal Pools: A Scenario to Guide Experimental Approaches to the Origin of Cellular Life. Life 2015, 5, 872–887. [Google Scholar] [CrossRef] [Green Version]
- Damer, B. A Field Trip to the Archaean in Search of Darwin’s Warm Little Pond. Life 2016, 6, 21. [Google Scholar] [CrossRef] [Green Version]
- Damer, B.; Deamer, D. The Hot Spring Hypothesis for an Origin of Life. Astrobiology 2020, 20, 429–452. [Google Scholar] [CrossRef] [Green Version]
- Deamer, D. Where Did Life Begin? Testing Ideas in Prebiotic Analogue Conditions. Life 2021, 11, 134. [Google Scholar] [CrossRef]
- Trevors, J.T.; Pollack, G.H. Hypothesis: The origin of life in a hydrogel environment. Prog. Biophys. Mol. Biol. 2005, 89, 1–8. [Google Scholar] [CrossRef]
- Allwood, A.C.; Walter, M.R.; Burch, I.W.; Kamber, B.S. 3.43 billion-year-old stromatolite reef from the Pilbara Craton of Western Australia: Ecosystem-scale insights to early life on Earth. Precambrian Res. 2007, 158, 198–227. [Google Scholar] [CrossRef]
- Jefferson, K. What drives bacteria to produce a biofilm? FEMS Microbiol. Lett. 2004, 236, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, H.S. The RNA world hypothesis: The worst theory of the early evolution of life (except for all the others). Biol. Direct 2012, 7, 23. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, A.W. Phosphorus in prebiotic chemistry. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1743–1749. [Google Scholar] [CrossRef] [Green Version]
- Graaf, R.M.D.; Schwartz, A.W. Reduction and Activation of Phosphate on the Primitive Earth. Orig. Life Evol. Biosph. 2000, 30, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Patel, B.H.; Percivalle, C.; Ritson, D.J.; Duffy, C.D.; Sutherland, J.D. Common origins of RNA, protein and lipid precursors in a cyanosulfidic protometabolism. Nat. Chem. 2015, 7, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Maurette, M.; Brack, A.; Kurat, G.; Perreau, M.; Engrand, C. Were micrometeorites a source of prebiotic molecules on the early Earth? Adv. Space Res. 1995, 15, 113–126. [Google Scholar] [CrossRef]
- Rimmer, P.B.; Thompson, S.J.; Xu, J.; Russell, D.A.; Green, N.J.; Ritson, D.J.; Sutherland, J.D.; Queloz, D.P. Timescales for Prebiotic Photochemistry Under Realistic Surface Ultraviolet Conditions. Astrobiology 2021. [Google Scholar] [CrossRef]
- Cleaves, H.J.; Miller, S.L. Oceanic protection of prebiotic organic compounds from UV radiation. Proc. Natl. Acad. Sci. USA 1998, 95, 7260–7263. [Google Scholar] [CrossRef] [Green Version]
- Fleischmann, E.M. The measurement and penetration of ultraviolet radiation into tropical marine water. Limnol. Oceanogr. 1989, 34, 1623–1629. [Google Scholar] [CrossRef]
- Pilla, R.M.; Couture, R. Attenuation of photosynthetically active radiation and ultraviolet radiation in response to changing dissolved organic carbon in browning lakes: Modeling and parametrization. Limnol. Oceanogr. 2021, 66, 2278–2289. [Google Scholar] [CrossRef]
- Woese, C.R.; Fox, G.E. The concept of cellular evolution. J. Mol. Evol. 1977, 10, 1–6. [Google Scholar] [CrossRef]
- Wotton, R.S.; Preston, T.M. Surface Films: Areas of Water Bodies That Are Often Overlooked. BioScience 2005, 55, 137–145. [Google Scholar] [CrossRef]
- Hulshof, J.; Ponnamperuma, C. Prebiotic Condensation Reactions in an aqueous medium: A review of condensing agents. Orig. Life 1976, 7, 197–224. [Google Scholar] [CrossRef]
- Ross, D.S.; Deamer, D. Dry/Wet Cycling and the Thermodynamics and Kinetics of Prebiotic Polymer Synthesis. Life 2016, 6, 28. [Google Scholar] [CrossRef] [Green Version]
- Butcher, S.E.; Pyle, A.M. The Molecular Interactions That Stabilize RNA Tertiary Structure: RNA Motifs, Patterns, and Networks. Accounts Chem. Res. 2011, 44, 1302–1311. [Google Scholar] [CrossRef] [PubMed]
- Jaeger, J.A.; Turner, D.H.; Zuker, M. Improved predictions of secondary structures for RNA. Proc. Natl. Acad. Sci. USA 1989, 86, 7706–7710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freier, S.M.; Kierzek, R.; Jaeger, J.A.; Sugimoto, N.; Caruthers, M.H.; Neilson, T.; Turner, D.H. Improved free-energy parameters for predictions of RNA duplex stability. Proc. Natl. Acad. Sci. USA 1986, 83, 9373–9377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Disalvo, E.; Pinto, O.; Martini, M.; Bouchet, A.; Hollmann, A.; Frías, M. Functional role of water in membranes updated: A tribute to Träuble. Biochim. Biophys. Acta (BBA)-Biomembr. 2015, 1848, 1552–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Disalvo, E.; Martini, M.; Bouchet, A.; Hollmann, A.; Frías, M. Structural and thermodynamic properties of water–membrane interphases: Significance for peptide/membrane interactions. Adv. Colloid Interface Sci. 2014, 211, 17–33. [Google Scholar] [CrossRef]
- Disalvo, E.; Lairion, F.; Martini, F.; Tymczyszyn, E.; Frías, M.; Almaleck, H.; Gordillo, G. Structural and functional properties of hydration and confined water in membrane interfaces. Biochim. Biophys. Acta (BBA)-Biomembr. 2008, 1778, 2655–2670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damodaran, S. Water activity at interfaces and its role in regulation of interfacial enzymes: A hypothesis. Colloids Surf. B Biointerfaces 1998, 11, 231–237. [Google Scholar] [CrossRef]
- Rao, C.S.; Damodaran, S. Surface pressure dependence of phospholipase A2 activity in lipid monolayers is linked to interfacial water activity. Colloids Surf. B Biointerfaces 2004, 34, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Rao, C.S.; Damodaran, S. Is Interfacial Activation of Lipases in Lipid Monolayers Related to Thermodynamic Activity of Interfacial Water? Langmuir 2002, 18, 6294–6306. [Google Scholar] [CrossRef]
- Schiraldi, A.; Fessas, D.; Signorelli, M. Water Activity in Biological Systems—A Review. Pol. J. Food Nutr. Sci. 2012, 62, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Benkovic, S.J.; Hammes-Schiffer, S. A Perspective on Enzyme Catalysis. Science 2003, 301, 1196–1202. [Google Scholar] [CrossRef] [Green Version]
- Boekhoven, J.; Hendriksen, W.E.; Koper, G.J.M.; Eelkema, R.; Esch, J.H.v. Transient assembly of active materials fueled by a chemical reaction. Science 2015, 349, 1075–1079. [Google Scholar] [CrossRef]
- Hanson, R.W. The role of ATP in metabolism. Biochem. Educ. 1989, 17, 86–92. [Google Scholar] [CrossRef]
- Nam, I.; Lee, J.K.; Nam, H.G.; Zare, R.N. Abiotic production of sugar phosphates and uridine ribonucleoside in aqueous microdroplets. Proc. Natl. Acad. Sci. USA 2017, 114, 12396–12400. [Google Scholar] [CrossRef] [Green Version]
- Pasek, M.A. Thermodynamics of Prebiotic Phosphorylation. Chem. Rev. 2020, 11, 4690–4706. [Google Scholar] [CrossRef]
- Yousef, A.; Coy, B.M. Diffusion of surfactants to an interface: Effect of a barrier of oriented water. J. Colloid Interface Sci. 1983, 94, 497–501. [Google Scholar] [CrossRef]
- Cornell, C.E.; Black, R.A.; Xue, M.; Litz, H.E.; Ramsay, A.; Gordon, M.; Mileant, A.; Cohen, Z.R.; Williams, J.A.; Lee, K.K.; et al. Prebiotic amino acids bind to and stabilize prebiotic fatty acid membranes. Proc. Natl. Acad. Sci. USA 2019, 116, 17239–17244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, M.; Black, R.A.; Cohen, Z.R.; Roehrich, A.; Drobny, G.P.; Keller, S.L. Binding of Dipeptides to Fatty Acid Membranes Explains Their Colocalization in Protocells but Does Not Select for Them Relative to Unjoined Amino Acids. J. Phys. Chem. B 2021, 125, 7933–7939. [Google Scholar] [CrossRef] [PubMed]
- Powner, M.W.; Gerland, B.; Sutherland, J.D. Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions. Nature 2009, 459, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Deamer, D. Electrochemical energy for living systems. Curr. Opin. Electrochem. 2021, 29, 100742. [Google Scholar] [CrossRef]
- Summers, D.P.; Rodoni, D. Vesicle Encapsulation of a Nonbiological Photochemical System Capable of Reducing NAD+ to NADH. Langmuir 2015, 31, 10633–10637. [Google Scholar] [CrossRef] [PubMed]
- Deamer, D. Polycyclicaromatic Hydrocarbons: Primitive Pigment Systems in th e Prebiotic Environment. Adv. Space Res. 1992, 12, 183–189. [Google Scholar] [CrossRef]
- Robertson, R.M.; Laib, S.; Smith, D.E. Diffusion of isolated DNA molecules: Dependence on length and topology. Proc. Natl. Acad. Sci. USA 2006, 103, 7310–7314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walton, C.R.; Shorttle, O. Scum of the Earth: A Hypothesis for Prebiotic Multi-Compartmentalised Environments. Life 2021, 11, 976. https://doi.org/10.3390/life11090976
Walton CR, Shorttle O. Scum of the Earth: A Hypothesis for Prebiotic Multi-Compartmentalised Environments. Life. 2021; 11(9):976. https://doi.org/10.3390/life11090976
Chicago/Turabian StyleWalton, Craig Robert, and Oliver Shorttle. 2021. "Scum of the Earth: A Hypothesis for Prebiotic Multi-Compartmentalised Environments" Life 11, no. 9: 976. https://doi.org/10.3390/life11090976
APA StyleWalton, C. R., & Shorttle, O. (2021). Scum of the Earth: A Hypothesis for Prebiotic Multi-Compartmentalised Environments. Life, 11(9), 976. https://doi.org/10.3390/life11090976