Independent Evolution of Sex Chromosomes in Eublepharid Geckos, A Lineage with Environmental and Genotypic Sex Determination




{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
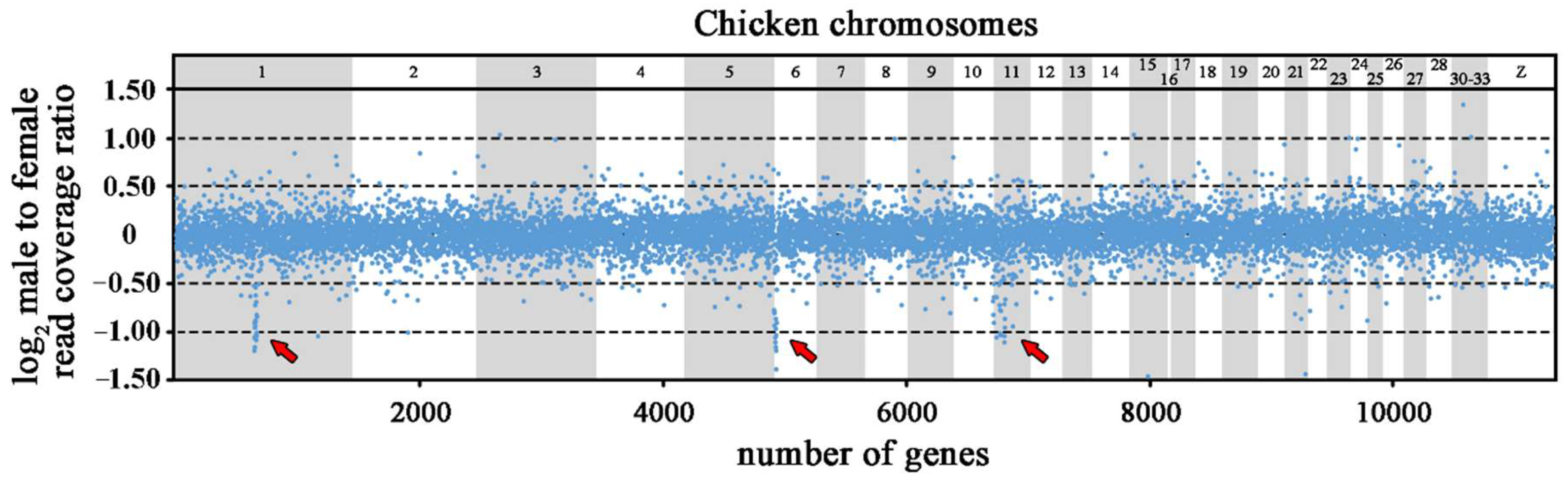
2.1. DNA-seq and Coverage Analysis
2.2. Validation of X-Specific Genes by qPCR in C. elegans and Test of Sex Chromosome Homology in Other Geckos
3. Discussion
4. Materials and Methods
4.1. Samples and Species Verification
4.2. DNA-seq and Coverage Analysis
4.3. Validation of X-Specific Genes by qPCR in C. elegans and Test of Homology to Other Geckos
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Capel, B. Sex Determination in Vertebrates, 1st ed.; Elsevier Science: London, UK, 2019; pp. 1–395. [Google Scholar]
- Straková, B.; Rovatsos, M.; Kubička, L.; Kratochvíl, L. Evolution of sex determination in amniotes: Did stress and sequential hermaphroditism produce environmental determination? BioEssays 2020, 42, 2000050. [Google Scholar] [CrossRef] [PubMed]
- Spangenberg, V.; Arakelyan, M.; Cioffi, M.B.; Liehr, T.; Al-Rikabi, A.; Martynova, E.; Danielyan, F.; Stepanyan, I.; Galoyan, E.; Kolomiets, O. Cytogenetic mechanisms of unisexuality in rock lizards. Sci. Rep. 2020, 10, 8697. [Google Scholar] [CrossRef] [PubMed]
- Kratochvíl, L.; Vukić, J.; Červenka, J.; Kubička, L.; Johnson Pokorná, M.; Kukačková, D.; Rovatsos, M.; Piálek, L. Mixed-sex offspring produced via cryptic parthenogenesis in a lizard. Mol. Ecol. 2020, 29, 4118–4127. [Google Scholar] [CrossRef] [PubMed]
- Pokorná, M.; Kratochvíl, L. Phylogeny of sex-determining mechanisms in squamate reptiles: Are sex chromosomes an evolutionary trap? Zool. J. Linn. Soc. 2009, 156, 168–183. [Google Scholar] [CrossRef]
- Johnson Pokorná, M.; Kratochvíl, L. What was the ancestral sex-determining mechanism in amniote vertebrates? Biol. Rev. 2016, 91, 1–12. [Google Scholar] [CrossRef]
- Gamble, T.; Coryell, J.; Ezaz, T.; Lynch, J.; Scantlebury, D.P.; Zarkower, D. Restriction site-associated DNA sequencing (RAD-seq) reveals an extraordinary number of transitions among gecko sex-determining systems. Mol. Biol. Evol. 2015, 32, 1296–1309. [Google Scholar] [CrossRef]
- Waters, S.A.; Marshall Graves, J.A. Monotreme sex chromosomes - implications for the evolution of amniote sex chromosomes. Reprod. Fertil. Dev. 2009, 21, 943–951. [Google Scholar] [CrossRef]
- Zechner, U.; Hameister, H. Sex chromosomes in vertebrates: XX/XY against ZZ/ZW. Sex. Dev. 2011, 5, 266–271. [Google Scholar] [CrossRef]
- O’Meally, D.; Ezaz, T.; Georges, A.; Sarre, S.D.; Marshall Graves, J.A. Are some chromosomes particularly good at sex? Insights from amniotes. Chromosome Res. 2012, 20, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Livernois, A.M.; Graves, J.A.; Waters, P.D. The origin and evolution of vertebrate sex chromosomes and dosage compensation. Heredity 2012, 108, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Ezaz, T.; Srikulnath, K.; Graves, J.A. Origin of amniote sex chromosomes: An ancestral super-sex chromosome, or common requirements? J. Hered. 2017, 108, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Laopichienpong, N.; Kraichak, E.; Singchat, W.; Sillapaprayoon, S.; Muangmai, N.; Suntrarachun, S.; Baicharoen, S.; Peyachoknagul, S.; Chanhome, L.; Ezaz, T.; et al. Genome-wide SNP analysis of Siamese cobra (Naja kaouthia) reveals the molecular basis of transitions between Z and W sex chromosomes and supports the presence of an ancestral super-sex chromosome in amniotes. Genomics 2020. [Google Scholar] [CrossRef] [PubMed]
- Singchat, W.; Ahmad, S.F.; Sillapaprayoon, S.; Muangmai, N.; Duengkae, P.; Peyachoknagul, S.; O’Connor, R.E.; Griffin, D.K.; Srikulnath, K. Partial amniote sex chromosomal linkage homologies shared on snake W sex chromosomes support the ancestral super-sex chromosome evolution in amniotes. Front. Genet. 2020, 11, 948. [Google Scholar] [CrossRef] [PubMed]
- Marshall Graves, J.A.; Peichel, C.L. Are homologies in vertebrate sex determination due to shared ancestry or to limited options? Genome Biol. 2010, 11, 205. [Google Scholar] [CrossRef] [PubMed]
- Rovatsos, M.; Farkačová, K.; Altmanová, M.; Johnson Pokorná, M.; Kratochvíl, L. The rise and fall of differentiated sex chromosomes in geckos. Mol. Ecol. 2019, 28, 3042–3052. [Google Scholar] [CrossRef]
- Pyron, R.A.; Burbrink, F.T.; Wiens, J.J. A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes. BMC Evol. Biol. 2013, 13, 93. [Google Scholar] [CrossRef]
- Zheng, Y.; Wiens, J.J. Combining phylogenomic and supermatrix approaches, and a time-calibrated phylogeny for squamate reptiles (lizards and snakes) based on 52 genes and 4162 species. Mol. Phylogenet. Evol. 2016, 94, 537–547. [Google Scholar] [CrossRef]
- Kostmann, A.; Kratochvíl, L.; Rovatsos, M. Poorly differentiated XX/XY sex chromosomes are widely shared across skink radiation. BioRxiv 2020. [Google Scholar] [CrossRef]
- Gamble, T. A review of sex determining mechanisms in geckos (Gekkota: Squamata). Sex. Dev. 2010, 4, 88–103. [Google Scholar] [CrossRef]
- Pokorná, M.; Rens, W.; Rovatsos, M.; Kratochvíl, L. A ZZ/ZW sex chromosome system in the thick-tailed gecko (Underwoodisaurus milii; Squamata: Gekkota: Carphodactylidae), a member of the ancient gecko lineage. Cytogenet. Genome Res. 2014, 142, 190–196. [Google Scholar] [CrossRef]
- Rovatsos, M.; Gamble, T.; Nielsen, S.V.; Georges, A.; Ezaz, T.; Kratochvíl, L. Do male and female heterogamety really differ in expression regulation? Lack of global dosage balance in pygopodid geckos. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, in press. [Google Scholar] [CrossRef]
- Nielsen, S.V.; Daza, J.D.; Pinto, B.J.; Gamble, T. ZZ/ZW sex chromosomes in the endemic Puerto Rican leaf-toed gecko (Phyllodactylus wirshingi). Cytogenet. Genome Res. 2019, 157, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Keating, S.E.; Griffing, A.H.; Nielsen, S.V.; Scantlebury, D.P.; Gamble, T. Conserved ZZ/ZW sex chromosomes in Caribbean croaking geckos (Aristelliger: Sphaerodactylidae). J. Evol. Biol. 2020, 33, 1316–1326. [Google Scholar] [CrossRef] [PubMed]
- Uetz, P.; Freed, P.; Hošek, J. The Reptile Database. Available online: http://www.reptile-database.org (accessed on 3 November 2020).
- Kluge, A.G. Cladistic Relationships in the Gekkonoidea (Squamata, Sauria); Museum of Zoology, University of Michigan: Ann Arbor, MI, USA, 1987; Volume 173, p. 153. [Google Scholar]
- Grismer, L.L. Phylogeny, taxonomy, classification, and biogeography of eublepharid geckos. In Phylogenetic Relationships of the Lizard Families; Estes, R., Pregill, G., Eds.; Stanford University Press: Stanford, CA, USA, 1988; pp. 369–469. [Google Scholar]
- Gamble, T.; Bauer, A.M.; Colli, G.R.; Greenbaum, E.; Jackman, T.R.; Vitt, L.J.; Simons, A.M. Coming to America: Multiple origins of New World geckos. J. Evol. Biol. 2011, 24, 231–244. [Google Scholar] [CrossRef]
- Pyron, R.A.; Burbrink, F.T. Early origin of viviparity and multiple reversions to oviparity in squamate reptiles. Ecol. Lett. 2014, 17, 13–21. [Google Scholar] [CrossRef]
- Tonini, J.F.R.; Beard, K.H.; Ferreira, R.B.; Jetz, W.; Pyron, R.A. Fully-sampled phylogenies of squamates reveal evolutionary patterns in threat status. Biol. Conserv. 2016, 204, 23–31. [Google Scholar] [CrossRef]
- Burbrink, F.T.; Grazziotin, F.G.; Pyron, R.A.; Cundall, D.; Donnellan, S.; Irish, F.; Keogh, J.S.; Kraus, F.; Murphy, R.W.; Noonan, B.; et al. Interrogating genomic scale data for Squamata (lizards, snakes, and amphisbaenians) shows no support for key traditional morphological relationships. Syst. Biol. 2020, 69, 502–520. [Google Scholar] [CrossRef]
- Singhal, S.; Colston, T.J.; Grundler, M.R.; Smith, A.S.; Costa, G.C.; Colli, G.R.; Moritz, C.; Pyron, R.A.; Rabosky, D.L. Congruence and conflict in the higher-level phylogenetics of squamate rReptiles: An expanded phylogenomic perspective. Syst. Biol. 2020. [Google Scholar] [CrossRef]
- Thorogood, J.; Whimster, I.W. The maintenance and breeding of the leopard gecko Eublepharis macularius as a laboratory animal. Int. Zoo Yearb. 1979, 19, 74–78. [Google Scholar] [CrossRef]
- Wagner, E. Gecko husbandry and reproduction. In Reproductive Biology and Diseases of Captive Reptiles; Murphy, J.B., Collins, J.T., Eds.; Society for the Study of Amphibians and Reptiles: Lawrence, KS, USA, 1980; pp. 115–117. [Google Scholar]
- Wagner, E. Temperature-dependent sex determination in a gekko lizard. Q. Rev. Biol. 1980, 55, 21. [Google Scholar]
- Bull, J.J. Temperature-dependent sex determination in reptiles: Validity of sex diagnosis in hatchling lizards. Can. J. Zool. 1987, 65, 1421–1424. [Google Scholar] [CrossRef]
- Anderson, A. Captive husbandry and reproduction of the African fat-tailed gecko, Hemitheconyx caudicinctus. Dactylus 1993, 2, 12–16. [Google Scholar]
- Viets, B.E.; Tousignant, A.; Ewert, M.A.; Nelson, C.E.; Crews, D. Temperature-dependent sex determination in the leopard gecko, Eublepharis macularius. J. Exp. Zool. 1993, 265, 679–683. [Google Scholar] [CrossRef] [PubMed]
- Viets, B.E.; Ewert, M.A.; Talent, L.G.; Nelson, C.E. Sex-determining mechanisms in squamate reptiles. J. Exp. Zool. 1994, 270, 45–56. [Google Scholar] [CrossRef]
- Bragg, W.K.; Fawcett, J.D.; Bragg, T.B.; Viets, B.E. Nest-site selection in two eublepharid gecko species with temperature-dependent sex determination and one with genotypic sex determination: Biol. J. Linn. Soc. 2000, 69, 319–332. [Google Scholar] [CrossRef]
- Kratochvíl, L.; Kubička, L.; Landová, E. Does the mechanism of sex determination constrain the potential for sex manipulation? A test in geckos with contrasting sex-determining systems. Naturwissenschaften 2008, 95, 209–215. [Google Scholar] [CrossRef]
- Pokorná, M.; Rábová, M.; Ráb, P.; Ferguson-Smith, M.A.; Rens, W.; Kratochvíl, L. Differentiation of sex chromosomes and karyotypic evolution in the eye-lid geckos (Squamata: Gekkota: Eublepharidae), a group with different modes of sex determination. Chromosome Res. 2010, 18, 809–820. [Google Scholar] [CrossRef]
- Kubička, L.; Golinski, A.; John-Alder, H.; Kratochvíl, L. Ontogeny of pronounced female-biased sexual size dimorphism in the Malaysian cat gecko (Aeluroscalabotes felinus: Squamata: Eublepharidae): A test of the role of testosterone in growth regulation. Gen. Comp. Endocrinol. 2013, 188, 183–188. [Google Scholar] [CrossRef]
- Golinski, A.; Kubička, L.; John-Alder, H.; Kratochvíl, L. Androgenic control of male-typical behavior, morphology and sex recognition is independent of the mode of sex determination: A case study on Lichtenfelder’s gecko (Eublepharidae: Goniurosaurus lichtenfelderi). Horm. Behav. 2015, 72, 49–59. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, Q.; Wang, Y.; Luo, L.; Yang, J.; Yang, L.; Liu, M.; Li, Y.; Qian, T.; Zheng, Y.; et al. Gekko japonicus genome reveals evolution of adhesive toe pads and tail regeneration. Nat. Commun. 2015, 6, 10033. [Google Scholar] [CrossRef] [PubMed]
- Cortez, D.; Marin, R.; Toledo-Flores, D.; Froidevaux, L.; Liechti, A.; Waters, P.D.; Grützner, F.; Kaessmann, H. Origins and functional evolution of Y chromosomes across mammals. Nature 2014, 508, 488–493. [Google Scholar] [CrossRef] [PubMed]
- Carré, G.A.; Siggers, P.; Xipolita, M.; Brindle, P.; Lutz, B.; Wells, S.; Greenfield, A. Loss of p300 and CBP disrupts histone acetylation at the mouse Sry promoter and causes XY gonadal sex reversal. Hum. Mol. Genet. 2018, 27, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Colvin, J.S.; Green, R.P.; Schmahl, J.; Capel, B.; Ornitz, D.M. Male-to-female sex reversal in mice lacking fibroblast growth factor 9. Cell 2001, 104, 875–889. [Google Scholar] [CrossRef]
- Bagheri-Fam, S.; Sim, H.; Bernard, P.; Jayakody, I.; Taketo, M.M.; Scherer, G.; Harley, V.R. Loss of Fgfr2 leads to partial XY sex reversal. Dev. Biol. 2008, 314, 71–83. [Google Scholar] [CrossRef][Green Version]
- Polanco, J.C.; Wilhelm, D.; Davidson, T.L.; Knight, D.; Koopman, P. Sox10 gain-of-function causes XX sex reversal in mice: Implications for human 22q-linked disorders of sex development. Hum. Mol. Genet. 2009, 19, 506–516. [Google Scholar] [CrossRef]
- Falah, N.; Posey, J.E.; Thorson, W.; Benke, P.; Tekin, M.; Tarshish, B.; Lupski, J.R.; Harel, T. 22q11.2q13 duplication including SOX10 causes sex-reversal and peripheral demyelinating neuropathy, central dysmyelinating leukodystrophy, Waardenburg syndrome, and Hirschsprung disease. Am. J. Med. Genet. 2017, 173, 1066–1070. [Google Scholar] [CrossRef]
- Seufer, H.; Kaverkin, Y.; Kirschner, A. The Eyelash Geckos: Care, Breeding and Natural History; Kirschner & Seufer: Karlsruhe, Germany, 2005. [Google Scholar]
- Kawai, A.; Ishijima, J.; Nishida, C.; Kosaka, A.; Ota, H.; Kohno, S.; Matsuda, Y. The ZW sex chromosomes of Gekko hokouensis (Gekkonidae, Squamata) represent highly conserved homology with those of avian species. Chromosoma 2009, 118, 43–51. [Google Scholar] [CrossRef]
- Koubová, M.; Johnson Pokorná, M.; Rovatsos, M.; Farkačová, K.; Altmanová, M.; Kratochvíl, L. Sex determination in Madagascar geckos of the genus Paroedura (Squamata: Gekkonidae): Are differentiated sex chromosomes indeed so evolutionary stable? Chromosome Res. 2014, 22, 441–452. [Google Scholar] [CrossRef]
- Nagy, Z.T.; Sonet, G.; Glaw, F.; Vences, M. First large-scale DNA barcoding assessment of reptiles in the biodiversity hotspot of Madagascar, based on newly designed COI primers. PLoS ONE. 2012, 7, e34506. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A quality control tool for high throughput sequence data. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc2010 (accessed on 3 November 2020).
- Vicoso, B.; Emerson, J.J.; Zektser, Y.; Mahajan, S.; Bachtrog, D. Comparative sex chromosome genomics in snakes: Differentiation, evolutionary strata, and lack of global dosage compensation. PLoS Biol. 2013, 11, e1001643. [Google Scholar] [CrossRef] [PubMed]
- Picard, M.A.L.; Cosseau, C.; Ferré, S.; Quack, T.; Grevelding, C.G.; Couté, Y.; Vicoso, B. Evolution of gene dosage on the Z-chromosome of schistosome parasites. eLife 2018, 7, 35684. [Google Scholar] [CrossRef]
- Alföldi, J.; Di Palma, F.; Grabherr, M.; Williams, C.; Kong, L.; Mauceli, E.; Russell, P.J.; Lowe, C.B.; Glor, R.E.; Jaffe, J.D.; et al. The genome of the green anole lizard and a comparative analysis with birds and mammals. Nature 2011, 477, 587–591. [Google Scholar] [CrossRef]
- Andrade, P.; Pinho, C.; De Lanuza, G.P.; Afonso, S.; Brejcha, J.; Rubin, C.-J.; Wallerman, O.; Pereira, P.; Sabatino, S.J.; Bellati, A.; et al. Regulatory changes in pterin and carotenoid genes underlie balanced color polymorphisms in the wall lizard. Proc. Natl. Acad. Sci. USA 2019, 116, 5633–5642. [Google Scholar] [CrossRef]
- International Chicken Genome Sequencing Consortium. Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature 2004, 432, 695–716. [Google Scholar] [CrossRef]
- Rovatsos, M.; Altmanová, M.; Pokorná, M.; Kratochvíl, L. Conserved sex chromosomes across adaptively radiated Anolis lizards. Evolution 2014, 68, 2079–2085. [Google Scholar] [CrossRef]
- Rovatsos, M.; Altmanová, M.; Johnson Pokorná, M.; Kratochvíl, L. Novel X-linked genes revealed by quantitative polymerase chain reaction in the green anole, Anolis carolinensis. G3 2014, 4, 2107–2113. [Google Scholar] [CrossRef]
- Rovatsos, M.; Rehák, I.; Velenský, P.; Kratochvíl, L. Shared ancient sex chromosomes in varanids, beaded lizards, and alligator lizards. Mol. Biol. Evol. 2019, 36, 1113–1120. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.G.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3 — new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pensabene, E.; Kratochvíl, L.; Rovatsos, M. Independent Evolution of Sex Chromosomes in Eublepharid Geckos, A Lineage with Environmental and Genotypic Sex Determination. Life 2020, 10, 342. https://doi.org/10.3390/life10120342
Pensabene E, Kratochvíl L, Rovatsos M. Independent Evolution of Sex Chromosomes in Eublepharid Geckos, A Lineage with Environmental and Genotypic Sex Determination. Life. 2020; 10(12):342. https://doi.org/10.3390/life10120342
Chicago/Turabian StylePensabene, Eleonora, Lukáš Kratochvíl, and Michail Rovatsos. 2020. "Independent Evolution of Sex Chromosomes in Eublepharid Geckos, A Lineage with Environmental and Genotypic Sex Determination" Life 10, no. 12: 342. https://doi.org/10.3390/life10120342
APA StylePensabene, E., Kratochvíl, L., & Rovatsos, M. (2020). Independent Evolution of Sex Chromosomes in Eublepharid Geckos, A Lineage with Environmental and Genotypic Sex Determination. Life, 10(12), 342. https://doi.org/10.3390/life10120342