

Taphonomic Analysis of the Sinotubulites from the Shibantan Member of the Dengying Formation in Yangtze Gorges Area (China)

Abstract
1. Introduction
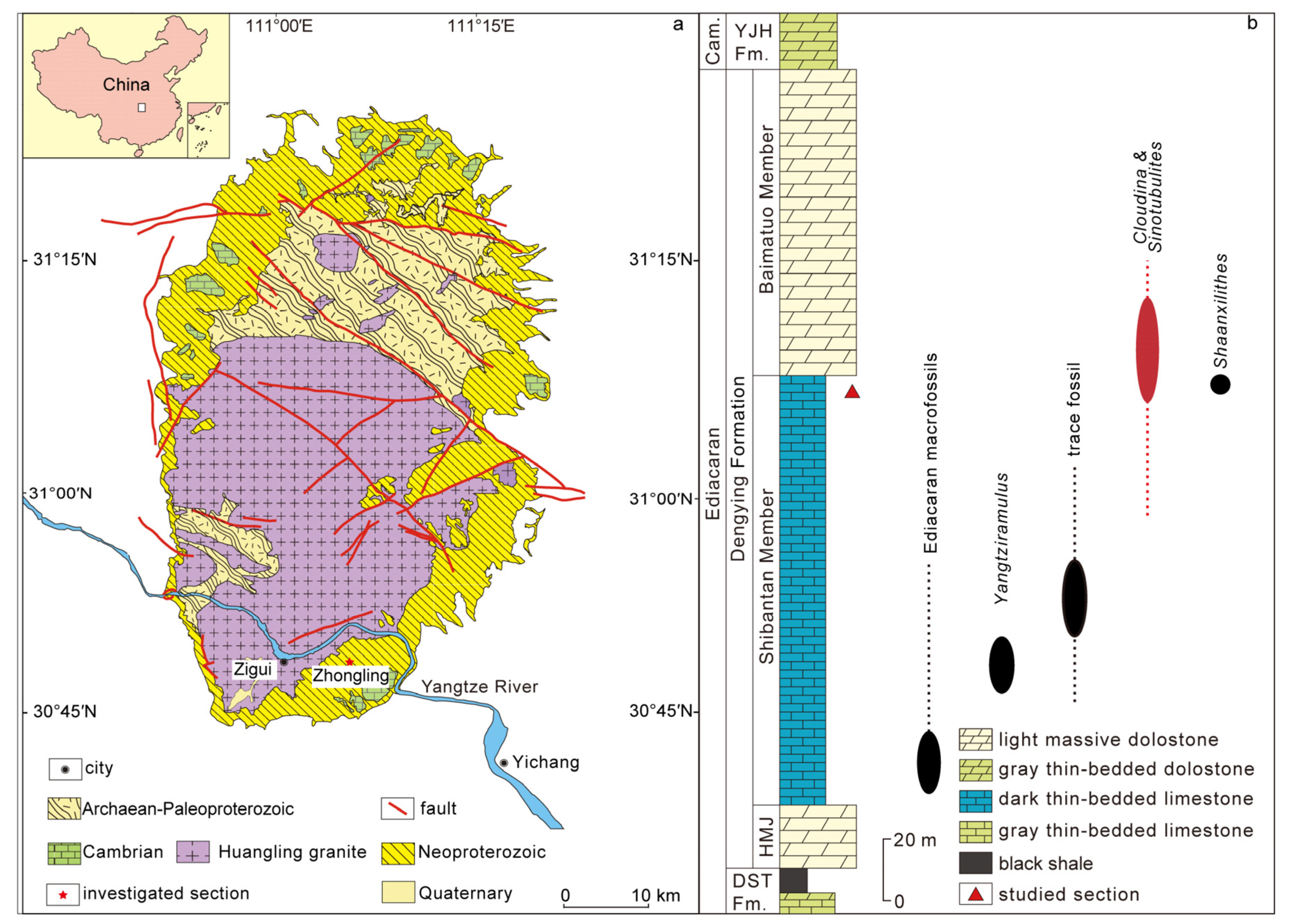
2. Geological Background and Research Methods
Methods
3. Results
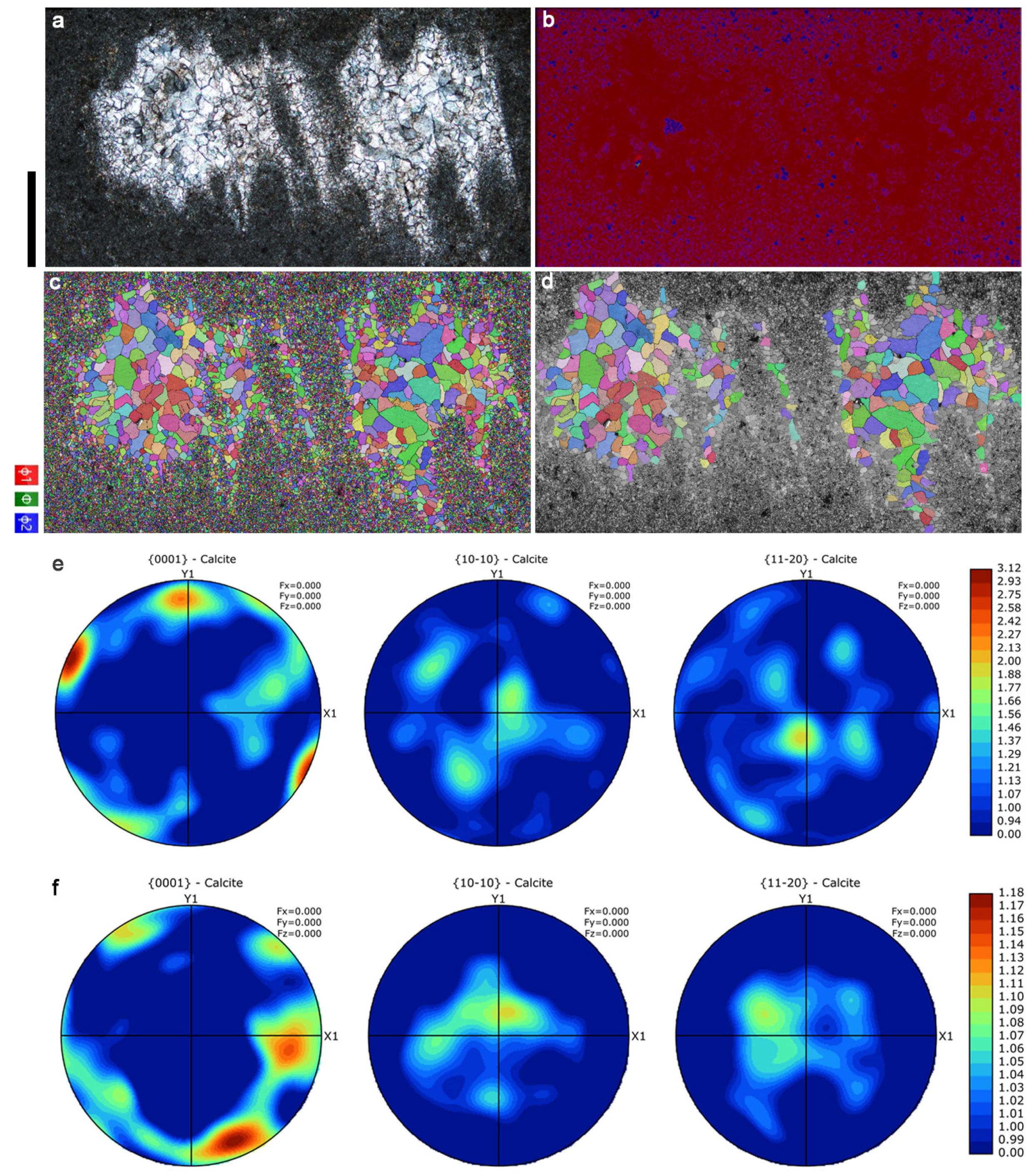
3.1. EBSD Analysis
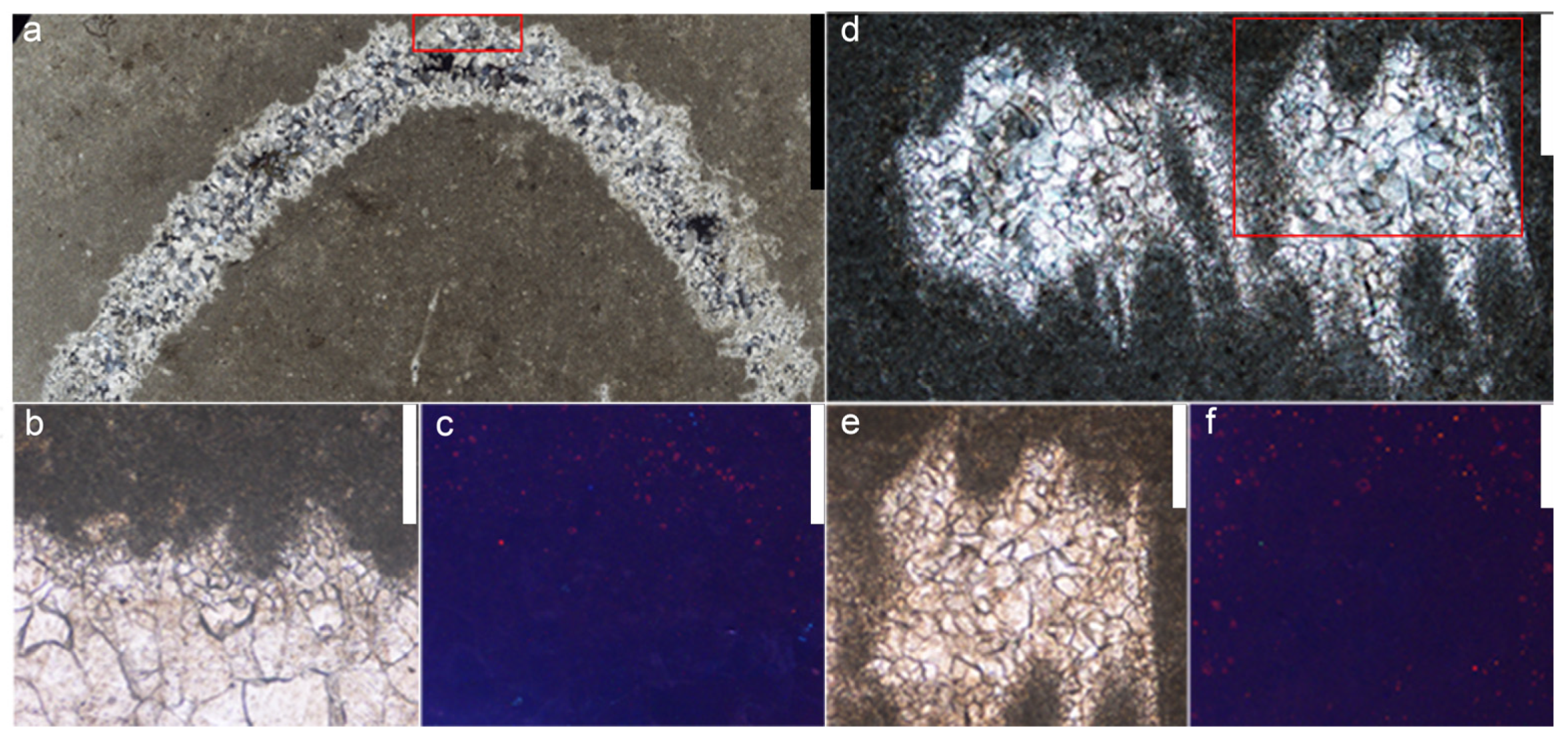
3.2. Cathodoluminescence Analysis
4. Discussion
4.1. Taphonomic Implications
4.2. Stratigraphic and Palaeogeographic Implications
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dohrmann, M.; Wörheide, G. Dating early animal evolution using phylogenomic data. Sci. Rep. 2017, 7, 3599. [Google Scholar] [CrossRef] [PubMed]
- Dos Reis, M.; Thawornwattana, Y.; Angelis, K.; Telford, M.J.; Donoghue, P.C.J.; Yang, Z. Uncertainty in the Timing of Origin of Animals and the Limits of Precision in Molecular Timescales. Curr. Biol. 2015, 25, 2939–2950. [Google Scholar] [CrossRef] [PubMed]
- Jensen, S. The Proterozoic and Earliest Cambrian Trace Fossil Record; Patterns, Problems and Perspectives. Integr. Comp. Biol. 2003, 43, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Matthews, J.J.; Liu, A.G.; Yang, C.; McIlroy, D.; Levell, B.; Condon, D.J. A Chronostratigraphic Framework for the Rise of the Ediacaran Macrobiota: New Constraints from Mistaken Point Ecological Reserve, Newfoundland. GSA Bull. 2020, 133, 612–624. [Google Scholar] [CrossRef]
- Narbonne, G.M. THE EDIACARA BIOTA: Neoproterozoic Origin of Animals and Their Ecosystems. Annu. Rev. Earth Planet. Sci. 2005, 33, 421–442. [Google Scholar] [CrossRef]
- Fedonkin, M.A.; Gehling, J.G.; Grey, K.; Narbonne, G.M.; Vickers-Rich, P. The Rise of Animals: Evolution and Diversification of the Kingdom Animalia; Johns Hopkins University Press: Baltimore, MD, USA, 2007; pp. 1–326. [Google Scholar]
- Xiao, S.; Laflamme, M. On the eve of animal radiation: Phylogeny, ecology and evolution of the Ediacara biota. Trends Ecol. Evol. 2009, 24, 31–40. [Google Scholar] [CrossRef]
- Droser, M.L.; Tarhan, L.G.; Gehling, J.G. The Rise of Animals in a Changing Environment: Global Ecological Innovation in the Late Ediacaran. Annu. Rev. Earth Planet. Sci. 2017, 45, 593–617. [Google Scholar] [CrossRef]
- Morris, S.C. The Fossil Record and the Early Evolution of the Metazoa. Nature 1993, 361, 219–225. [Google Scholar] [CrossRef]
- Germs, G. New shelly fossils from Nama Group, South West Africa. Am. J. Sci. 1972, 272, 752–761. [Google Scholar] [CrossRef]
- Hua, H.; Chen, Z.; Yuan, X.; Zhang, L.; Xiao, S. Skeletogenesis and asexual reproduction in the earliest biomineralizing animal Cloudina. Geology 2005, 33, 277–280. [Google Scholar] [CrossRef]
- Hua, H.; Cai, Y.P.; Min, X.; Chai, S.; Dai, Q.K. “Tubular animal radiation” at the terminal Ediacaran stage. J. Northwest Univ. 2020, 50, 141–174, (In Chinese with English Abstract). [Google Scholar]
- Yang, B.; Steiner, M.; Zhu, M.; Li, G.; Liu, J.; Liu, P. Transitional Ediacaran–Cambrian small skeletal fossil assemblages from South China and Kazakhstan: Implications for chronostratigraphy and metazoan evolution. Precambrian Res. 2016, 285, 202–215. [Google Scholar] [CrossRef]
- Chen, M.; Chen, Y.; Qian, Y. Some tubular fossils from Sinian–lower Cambrian boundary sequences, Yangtze Gorge. Bull. Tianjin Inst. Geol. Miner. Resour. 1981, 3, 117–124. (In Chinese) [Google Scholar]
- Yang, B.; Shang, X.; Steiner, M.; Liu, P. Ediacaran tubular fossils from the Shennongjia area, Hubei Province, and their stratigraphic significance. J. Stratigr. 2020, 44, 448–454, (In Chinese with English Abstract). [Google Scholar]
- Zhang, L.; Dong, J.; Tian, S.; Ding, L. The Gaojiashan Biota. In Late Aurignacian–Early Precambrian biota of the northern margin of the Yangzi Plateau; Ding, L., Zhang, L., Li, Y., Dong, J., Eds.; Science and Technology Literature Press: Beijing, China, 1992; p. 156. (In Chinese) [Google Scholar]
- Adorno, R.R.; Walde, D.H.G.; Erdtmann, B.D.; Denezine, M.; Cortijo, I.; Carmo, D.A.D.; Giorgioni, M.; Ramos, M.E.A.F.; Fazio, G. First occurrence of Cloudina carinata Cortijo et al., 2010 in South America, Tamengo Formation, Corμmbá Group, upper Ediacaran of Midwestern. Estud. Geol. 2019, 75, e095. [Google Scholar] [CrossRef]
- Walde, D.H.G.; do Carmo, D.A.; Guimarães, E.M.; Vieira, L.C.; Erdtmann, B.-D.; Sanchez, E.A.M.; Adorno, R.R.; Tobias, T.C. New aspects of Neoproterozoic—Cambrian transition in the Corumbá region (state of Mato Grosso do Sul, Brazil). Ann. Paléontol. 2015, 101, 213–224. [Google Scholar] [CrossRef]
- Walde, D.H.-G.; Weber, B.; Erdtmann, B.-D.; Steiner, M. Taphonomy of Corμmbella werneri from the Ediacaran of Brazil: Sinotubulitid tube or conulariid test? Alcheringa 2019, 43, 335–350. [Google Scholar] [CrossRef]
- Warren, L.V.; Quaglio, F.; Simões, M.G.; Gaucher, C.; Riccomini, C.; Poiré, D.G.; Freitas, B.T.; Boggiani, P.C.; Sial, A.N. Cloudina-Corμmbella-Namacalathus association from the Itapucμmi Group, Paraguay: Increasing ecosystem complexity and tiering at the end of the Ediacaran. Precambrian Res. 2017, 298, 79–87. [Google Scholar] [CrossRef]
- Yang, B.; Warren, L.V.; Steiner, M.; Smith, E.F.; Liu, P. Taxonomic revision of Ediacaran tubular fossils: Cloudina, Sinotubulites and Conotubus. J. Paleontol. 2022, 96, 256–273. [Google Scholar] [CrossRef]
- Hagadorn, J.W.; Waggoner, B. Ediacaran fossils from the southwestern Great Basin, United States. J. Paleontol. 2000, 74, 349–359. [Google Scholar] [CrossRef]
- Selly, T.; Schiffbauer, J.D.; Jacquet, S.M.; Smith, E.F.; Nelson, L.L.; Andreasen, B.D.; Huntley, J.W.; Strange, M.A.; O’Neil, G.R.; Thater, C.A. A new cloudinid fossil assemblage from the terminal Ediacaran of Nevada, USA. J. Syst. Palaeontol. 2020, 18, 357–379. [Google Scholar] [CrossRef]
- Cortijo, I.; Martí Mus, M.; Jensen, S.; Palacios, T. Late Ediacaran skeletal body fossil assemblage from the Navalpino anticline, central Spain. Precambrian Res. 2015, 267, 186–195. [Google Scholar] [CrossRef]
- Yang Ben Steiner, M.; Schiffbauer, J.D.; Selly, T.; Wu, X.; Zhang, C.; Liu, P. Ultrastructure of Ediacaran cloudinids suggests diverse taphonomic histories and affinities with non-biomineralized annelids. Sci. Rep. 2020, 10, 535. [Google Scholar]
- Zhuravlev, A.Y.; Liñán, E.; Vintaned, J.A.G.; Debrenne, F.; Fedorov, A.B. New Finds of Skeletal Fossils in the Terminal Neoproterozoic of the Siberian Platform and Spain. Acta Palaeontol. Pol. 2012, 57, 205–224. [Google Scholar] [CrossRef]
- Grant, S. Shell structure and distribution of Cloudina, a potential index fossil for the terminal Proterozoic. Am. J. Sci. 1990, 290-A, 261–294. [Google Scholar]
- Vinn, O.; Zaton, M. Inconsistencies in proposed annelid affinities of early biomineralized organism Cloudina (Ediacaran): Structural and ontogenetic evidences. Carnets Géol. 2012, 3, 39–47. [Google Scholar] [CrossRef]
- Beurlen, K.; Sommer, F.W. Observações estratigráficas e paleontológicas sôbre o calcário Corumbá. Bol. Geol. Mineral.—DNPM 1957, 168, 1–47. [Google Scholar]
- Chen, M.; Wang, Y. Note on the tubular metazoan fossils from the Dengying Formation of the upper Sinian System in the Gorge District of the Yangtze River. Chin. Sci. Bull. 1977, 22, 219–221. (In Chinese) [Google Scholar]
- Hua, H.; Zhang, L.; Zhang, Z.; Wang, J. New fossil evidences from latest Neoproterozoic Gaojiashan biota, South Shaanxi. J. Paleontol. 2000, 39, 381–390. (In Chinese) [Google Scholar]
- Cai, Y.; Xiao, S.; Hua, H.; Yuan, X. New material of the biomineralizing tubular fossil Sinotubulites from the late Ediacaran Dengying Formation, South China. Precambrian Res. 2015, 261, 12–24. [Google Scholar] [CrossRef]
- Liang, D.; Cai, Y.; Nolan, M.; Xiao, S. The terminal Ediacaran tubular fossil Cloudina in the Yangtze Gorges area of South China. Precambrian Res. 2020, 351, 105931. [Google Scholar] [CrossRef]
- Signor, P.W.; McMenamin, M.A.S.; Gevirtzman, D.A.; Mount, J.F. Two new pretrilobite faunas from western North America. Nature 1983, 303, 415–418. [Google Scholar] [CrossRef]
- Signor, P.W.; Mount, J.F.; Onken, B.R. A pre-trilobite shelly fauna from the White–Inyo region of eastern California and western Nevada. J. Paleontol. 1987, 61, 425–438. [Google Scholar] [CrossRef]
- McMenamin, M.A.S. Basal Cambrian small shelly fossils from the La Ciénega Formation, northwestern Sonora, Mexico. J. Paleontol. 1985, 59, 1414–1425. [Google Scholar]
- Sour-Tovar, F.; Hagadorn, J.W.; Huitrón-Rubio, T. Ediacaran and Cambrian index fossils from Sonora, Mexico. Palaeontology 2007, 50, 169–175. [Google Scholar] [CrossRef]
- Hahn, G.; Pflug, H.D. Die Cloudinidae n. fam., KallRöhren aus dem Vendiμm und Unter-Kambriμm. Senckenberg. Lethaea 1985, 65, 413–431. [Google Scholar]
- Chen, Z.; Sun, W. Late Simian (tubular) metazoan fossils: Cloudina and Sinotubulites from southern Shaanxi. Microb. Palaeontol. Lett. 2001, 18, 180–193, 199–202. [Google Scholar]
- Doddapaneni, S.; Kumar, S.; Sharma, S.; Shankar, G.; Shettar, M.; Kumar, N.; Aroor, G.; Ahmad, S.M. Advancements in EBSD Techniques: A Comprehensive Review on Characterization of Composites and Metals, Sample Preparation, and Operational Parameters. J. Compos. Sci. 2025, 9, 132. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, Q.; Wang, X. Electron backscatter diffraction (EBSD) study of elongatoolithid eggs from China with microstructural and parataxonomic implications. Paleobiology 2024, 50, 330–345. [Google Scholar] [CrossRef]
- Cusack, M. Biomineral electron backscatter diffraction for palaeontology. Palaeontology 2016, 59, 171–179. [Google Scholar] [CrossRef]
- Grenier, C.; Berent, K.; Rodríguez-Navarro, A.; Vinn, O.; Checa, A.G. Microstructures, crystallography and growth patterns of serpulid tubeworms (Class Polychaeta). Mar. Biol. 2024, 171, 231. [Google Scholar] [CrossRef]
- An, Z.H.; Ye, Q.; Tong, J.N.; Tian, L.; Zhao, X. Stratigraphic Sequence of the Ediacaran Dengying Formation around the Huangling Anticline, Western Hubei Province: Implications for Regional Stratigraphic Correlation. Earth Sci. 2024, 49, 2992–3009, (In Chinese with English Abstract). [Google Scholar]
- Skinner, H.C.W.; Jahren, A.H. Biomineralization. Treatise Geochem. 2003, 8, 682. [Google Scholar]
- Levi-Kalisman, Y.; Falini, G.; Addadi, L.; Weiner, S. Structure of the nacreous organic matrix of a bivalve mollusk shell examined in the hydrated state using cryo-TEM. J. Struct. Biol. 2001, 135, 8–17. [Google Scholar] [CrossRef]
- Lowenstam, H.A.; Weiner, S. On Biomineralization; Oxford University Press: New York, NY, USA, 1989; ISBN 978-0-19-504977-0. [Google Scholar]
- Marin, F.; Luquet, G.; Marie, B.; Medakovic, D. Marin2007. In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2007; Volume 80, pp. 209–276. ISBN 978-0-12-373914-8. [Google Scholar]
- Nudelman, F.; Sommerdijk, N.A.J.M. Biomineralization as an Inspiration for Materials Chemistry. Angew. Chem. Int. Ed. 2012, 51, 6582–6596. [Google Scholar] [CrossRef]
- Zhao, C.; Zeng, X.; Li, X.; Zhen, J.; Zhou, P.; Gui, K.; Zhang, H. A discussion on the sedimentary environment of Shibantan Member of Dengying Formation in the eastern Yangtze Gorges area. Geol. China 2013, 40, 1129–1139. (In Chinese) [Google Scholar]
- Chen, Z.; Zhou, C.; Xiao, S.; Wang, W.; Guan, C.; Hua, H.; Yuan, X. New Ediacara fossils preserved in marine limestone and their ecological implications. Sci. Rep. 2014, 4, 4180. [Google Scholar] [CrossRef]
- Chen, Z.; Zhou, C.; Yuan, X.; Xiao, S. Death march of a segmented and trilobate bilaterian elucidates early animal evolution. Nature 2019, 573, 412–415. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Occurrence | Max. Length (mm) | Max. Diameter (mm) | References |
|---|---|---|---|
| Dengying Fm., Yichang, Hubei Prov., China | 221.24 | 14.7 | This study |
| Dengying Fm., Yichang, Hubei Prov., China | 18 | 5 | Chen et al., 1981 [14] |
| Dengying Fm., Shennongjia, Hubei Prov., China | 2.8 | 1 | Yang et al., 2020 [15] |
| Dengying Fm., southern Shaanxi Prov., China | 20 | 4 | Chen et al., 2001 [39] |
| Deep Spring Fm., Mount Dunfee, Nevada, USA | 7.6 | 3.3 | Signor et al., 1987 [35] |
| La Ciénega Fm., Sonora, Mexico | 22 | 2.9 | Sour-Tovar et al., 2006 [37] |
| Villarta Limestone of lbor Group, Bada-joz Prov., Spain | 20 | 6 | Cortijo et al., 2015 [24] |
| Corumbá Group, Mato Grosso do Sul State, Brazil | 8 | 3.1 | Hahn and Pflug, 1985 [38] |
| Mooifontein Member, Nama Group, South Namibia | 2.5 | 0.5 | Yang et al., 2022 [21] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Yang, B.; An, Z.; Zhao, Z. Taphonomic Analysis of the Sinotubulites from the Shibantan Member of the Dengying Formation in Yangtze Gorges Area (China). Minerals 2025, 15, 570. https://doi.org/10.3390/min15060570
Wang X, Yang B, An Z, Zhao Z. Taphonomic Analysis of the Sinotubulites from the Shibantan Member of the Dengying Formation in Yangtze Gorges Area (China). Minerals. 2025; 15(6):570. https://doi.org/10.3390/min15060570
Chicago/Turabian StyleWang, Xinjie, Ben Yang, Zhihui An, and Zhongbao Zhao. 2025. "Taphonomic Analysis of the Sinotubulites from the Shibantan Member of the Dengying Formation in Yangtze Gorges Area (China)" Minerals 15, no. 6: 570. https://doi.org/10.3390/min15060570
APA StyleWang, X., Yang, B., An, Z., & Zhao, Z. (2025). Taphonomic Analysis of the Sinotubulites from the Shibantan Member of the Dengying Formation in Yangtze Gorges Area (China). Minerals, 15(6), 570. https://doi.org/10.3390/min15060570