
Systematic Review: The Development of Behavioral Laterality Across the First Year of Life in Nonhuman Primates



Abstract
1. Introduction
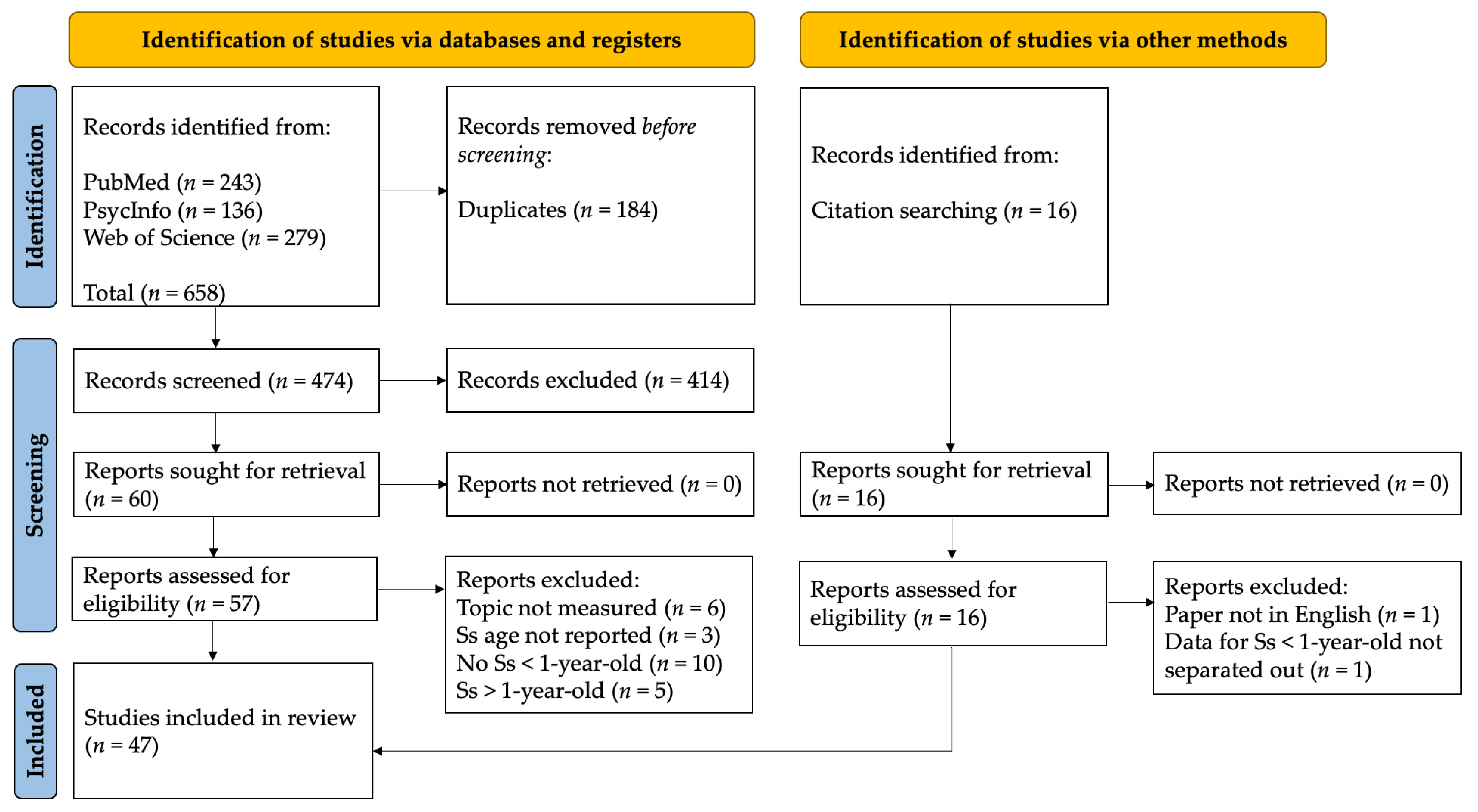
2. Methods
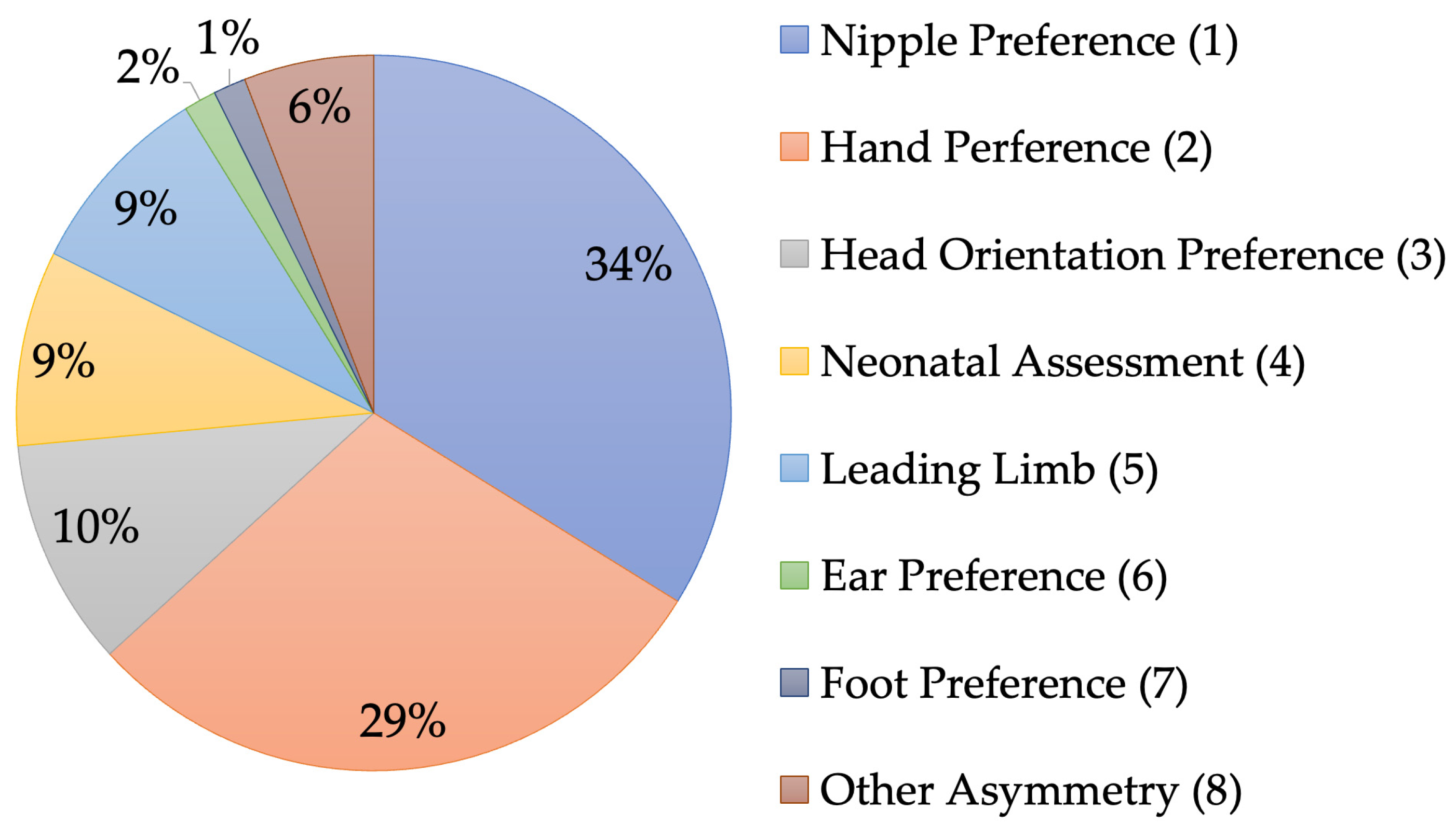
3. Results and Discussion
3.1. Considerations for Measuring Nipple Preference Across the First Year of Life
3.2. Considerations for Measuring Hand Preference across the First Year of Life
3.3. Increasing Developmental Rigor in Primate Behavioral Laterality Studies
3.3.1. Developmental Rigor in Platyrrhine Studies
3.3.2. Developmental Rigor in Catarrhine Studies
3.3.3. Developmental Rigor in Ape Studies
4. Limitations and Future Directions
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Database | Search Strategy |
|---|---|
| PubMed | Search: ((“catarrhini”[mh:noexp] OR “cercopithecidae”[mh] OR “gorilla gorilla”[mh] OR “haplorhini”[mh:noexp] OR “hominidae”[mh:noexp] OR “hylobatidae”[mh] OR “pan paniscus”[mh] OR “pan troglodytes”[mh] OR “platyrrhini”[mh] OR “pongo”[mh] OR “primates”[mh:noexp] OR “strepsirhini”[mh] OR “tarsii”[mh] OR “allenopithecus”[tiab] OR “allocebus”[tiab] OR “alouatta”[tiab] OR “alouattinae”[tiab] OR “angwantibo *”[tiab] OR “anthropoid”[tiab] OR “anthropoidea”[tiab] OR “anthropoids”[tiab] OR “aotes”[tiab] OR “aotidae”[tiab] OR “aotinae”[tiab] OR “aotus”[tiab] OR “ape”[tiab] OR “apes”[tiab] OR “arctocebus”[tiab] OR “ateles”[tiab] OR “atelidae”[tiab] OR “atelinae”[tiab] OR “avahi”[tiab] OR “aye-aye *”[tiab] OR “baboon”[tiab] OR “baboons”[tiab] OR “bonobo”[tiab] OR “bonobos”[tiab] OR “brachyteles”[tiab] OR “bushbabies”[tiab] OR “bushbaby”[tiab] OR “cacajao”[tiab] OR “callibella”[tiab] OR “callicebinae”[tiab] OR “callicebus”[tiab] OR “callimico”[tiab] OR “callithrichid *”[tiab] OR “callithrichinae”[tiab] OR “callithrix”[tiab] OR “callitrichid”[tiab] OR “callitrichidae”[tiab] OR “callitrichide”[tiab] OR “callitrichids”[tiab] OR “callitrichinae”[tiab] OR “capuchin”[tiab] OR “capuchins”[tiab] OR “carlito syrichta”[tiab] OR “catarhine *”[tiab] OR “catarhini”[tiab] OR “catarrhina”[tiab] OR “catarrhine *”[tiab] OR “catarrhini”[tiab] OR “cebid”[tiab] OR “cebidae”[tiab] OR “cebids”[tiab] OR “cebinae”[tiab] OR “ceboidea”[tiab] OR “cebuella”[tiab] OR “cebus”[tiab] OR “cephalopachus”[tiab] OR “cercocebus”[tiab] OR “cercopithecid *”[tiab] OR “cercopithecinae”[tiab] OR “cercopithecine *”[tiab] OR “cercopithecini”[tiab] OR “cercopithecoid”[tiab] OR “cercopithecoidea”[tiab] OR “cercopithecoids”[tiab] OR “cercopithecus”[tiab] OR “cheirogaleidae”[tiab] OR “cheirogaleus”[tiab] OR “cheracebus”[tiab] OR “chimp”[tiab] OR “chimpanzee”[tiab] OR “chimpanzees”[tiab] OR “chimps”[tiab] OR “chiromyiformes”[tiab] OR “chiropotes”[tiab] OR “chlorocebus”[tiab] OR “colobidae”[tiab] OR “colobinae”[tiab] OR “colobine *”[tiab] OR “colobini”[tiab] OR “colobus *”[tiab] OR “cynomolgus”[tiab] OR “daubentonia”[tiab] OR “daubentoniidae”[tiab] OR “douc”[tiab] OR “doucs”[tiab] OR “erythrocebus”[tiab] OR “eulemur”[tiab] OR “euoticus”[tiab] OR “euprimate *”[tiab] OR “galagid *”[tiab] OR “galago”[tiab] OR “galagoides”[tiab] OR “galagonidae”[tiab] OR “galagos”[tiab] OR “gelada”[tiab] OR “geladas”[tiab] OR “gibbon”[tiab] OR “gibbons”[tiab] OR “gorilla”[tiab] OR “gorillas”[tiab] OR “grivet”[tiab] OR “grivets”[tiab] OR “guenon *”[tiab] OR “guereza *”[tiab] OR “hapalemur”[tiab] OR “haplorrhine *”[tiab] OR “haplorhini”[tiab] OR “haplorrhine *”[tiab] OR “haplorrhini”[tiab] OR “hominid *”[tiab] OR “hominin”[tiab] OR “homininae”[tiab] OR “hominine”[tiab] OR “hominines”[tiab] OR “hominini”[tiab] OR “hominins”[tiab] OR “hominoidea”[tiab] OR “hoolock”[tiab] OR “howler *”[tiab] OR “hylobates”[tiab] OR “hylobatidae”[tiab] OR “indri”[tiab] OR “indridae”[tiab] OR “indriid *”[tiab] OR “indris”[tiab] OR “kipunji *”[tiab] OR “lagothrix”[tiab] OR “langur”[tiab] OR “langurs”[tiab] OR “lemur”[tiab] OR “lemurid *”[tiab] OR “lemuriform”[tiab] OR “lemuriformes”[tiab] OR “lemuriforms”[tiab] OR “lemurinae”[tiab] OR “lemuroidea”[tiab] OR “lemurs”[tiab] OR “leontideus”[tiab] OR “leontocebus”[tiab] OR “leontopithecus”[tiab] OR “lepilemur”[tiab] OR “lepilemurid *”[tiab] OR “lesula *”[tiab] OR “lophocebus”[tiab] OR “loriform”[tiab] OR “loriformes”[tiab] OR “lorinae”[tiab] OR “loris”[tiab] OR “lorises”[tiab] OR “lorisid *”[tiab] OR “lorisiform *”[tiab] OR “lorisinae”[tiab] OR “lorisoid *”[tiab] OR “lutung”[tiab] OR “lutungs”[tiab] OR “macaca”[tiab] OR “macaque’s”[tiab] OR “macaque”[tiab] OR “macaques”[tiab] OR “malbrouck *”[tiab] OR “mandrill”[tiab] OR “mandrills”[tiab] OR “mandrillus”[tiab] OR “mangabey *”[tiab] OR “marmoset”[tiab] OR “marmosets”[tiab] OR “mico argentatus”[tiab] OR “mico chrysoleucos”[tiab] OR “mico emiliae”[tiab] OR “mico humilis”[tiab] OR “mico marcai”[tiab] OR “mico melanurus”[tiab] OR “mico rondoni”[tiab] OR “microcebus”[tiab] OR “miopithecus”[tiab] OR “mirza coquereli”[tiab] OR “mirza zaza”[tiab] OR “monkey”[tiab] OR “monkeys”[tiab] OR “muriqui *”[tiab] OR “nasalis larvatus”[tiab] OR “nomascus”[tiab] OR “nycticebus”[tiab] OR “oedipomidas”[tiab] OR “orang utan *”[tiab] OR “orang-utan *”[tiab] OR “orangutan *”[tiab] OR “oreonax”[tiab] OR “otolemur”[tiab] OR “pan paniscus”[tiab] OR “pan troglodytes”[tiab] OR “panin”[tiab] OR “panina”[tiab] OR “panins”[tiab] OR “papio”[tiab] OR “papionini”[tiab] OR “paragalago”[tiab] OR “perodicticinae”[tiab] OR “perodicticus”[tiab] OR “phaner”[tiab] OR “piliocolobus”[tiab] OR “pithecia”[tiab] OR “pithecidae”[tiab] OR “pitheciid *”[tiab] OR “pitheciinae”[tiab] OR “pithecinae”[tiab] OR “platyrhine *”[tiab] OR “platyrhini”[tiab] OR “platyrrhina”[tiab] OR “platyrrhine *”[tiab] OR “platyrrhini”[tiab] OR “plecturocebus”[tiab] OR “pongid *”[tiab] OR “ponginae”[tiab] OR “pongo”[tiab] OR “potto”[tiab] OR “pottos”[tiab] OR “presbytini”[tiab] OR “presbytis”[tiab] OR “primate”[tiab] OR “primates”[tiab] OR “procolobus”[tiab] OR “prolemur”[tiab] OR “propithecus”[tiab] OR “prosimian *”[tiab] OR “prosimii”[tiab] OR “pseudopotto”[tiab] OR “pygathrix”[tiab] OR “rhinopithecus”[tiab] OR “rungwecebus”[tiab] OR “saguinus”[tiab] OR “saimiri”[tiab] OR “saimiriinae”[tiab] OR “sapajus”[tiab] OR “sciurocheirus”[tiab] OR “semnopithecus”[tiab] OR “siamang”[tiab] OR “siamangs”[tiab] OR “sifaka”[tiab] OR “sifakas”[tiab] OR “simians”[tiab] OR “simias”[tiab] OR “simiiform *”[tiab] OR “strepsir *”[tiab] OR “surili *”[tiab] OR “symphalangus”[tiab] OR “talapoin *”[tiab] OR “tamarin”[tiab] OR “tamarins”[tiab] OR “tamarinus”[tiab] OR “tarsier”[tiab] OR “tarsiers”[tiab] OR “tarsiid *”[tiab] OR “tarsiiform *”[tiab] OR “tarsius”[tiab] OR “theropithecus”[tiab] OR “trachypithecus”[tiab] OR “uacari *”[tiab] OR “uakari”[tiab] OR “uakaris”[tiab] OR “varecia”[tiab] OR “vervet *”[tiab]) AND (“laterality” or “handedness” or “hand preference” or “asymmetry”)) AND (“Infant” or “longitudinal” or “developmental” or “neonatal” or “neonate”) |
| PsycINFO | (DE(“baboons” OR “bonobos” OR “chimpanzees” OR “gorillas” OR “lemurs” OR “monkeys” OR “primates (nonhuman)”) OR TX(“allenopithecus” OR “allocebus” OR “alouatta” OR “alouattinae” OR “angwantibo *” OR “anthropoid” OR “anthropoidea” OR “anthropoids” OR “aotes” OR “aotidae” OR “aotinae” OR “aotus” OR “ape” OR “apes” OR “arctocebus” OR “ateles” OR “atelidae” OR “atelinae” OR “avahi” OR “aye-aye *” OR “baboon” OR “baboons” OR “bonobo” OR “bonobos” OR “brachyteles” OR “bushbabies” OR “bushbaby” OR “cacajao” OR “callibella” OR “callicebinae” OR “callicebus” OR “callimico” OR “callithrichid *” OR “callithrichinae” OR “callithrix” OR “callitrichid” OR “callitrichidae” OR “callitrichide” OR “callitrichids” OR “callitrichinae” OR “capuchin” OR “capuchins” OR “carlito syrichta” OR “catarhine *” OR “catarhini” OR “catarrhina” OR “catarrhine *” OR “catarrhini” OR “cebid” OR “cebidae” OR “cebids” OR “cebinae” OR “ceboidea” OR “cebuella” OR “cebus” OR “cephalopachus” OR “cercocebus” OR “cercopithecid *” OR “cercopithecinae” OR “cercopithecine *” OR “cercopithecini” OR “cercopithecoid” OR “cercopithecoidea” OR “cercopithecoids” OR “cercopithecus” OR “cheirogaleidae” OR “cheirogaleus” OR “cheracebus” OR “chimp” OR “chimpanzee” OR “chimpanzees” OR “chimps” OR “chiromyiformes” OR “chiropotes” OR “chlorocebus” OR “colobidae” OR “colobinae” OR “colobine *” OR “colobini” OR “colobus *” OR “cynomolgus” OR “daubentonia” OR “daubentoniidae” OR “douc” OR “doucs” OR “erythrocebus” OR “eulemur” OR “euoticus” OR “euprimate *” OR “galagid *” OR “galago” OR “galagoides” OR “galagonidae” OR “galagos” OR “gelada” OR “geladas” OR “gibbon” OR “gibbons” OR “gorilla” OR “gorillas” OR “grivet” OR “grivets” OR “guenon *” OR “guereza *” OR “hapalemur” OR “haplorhine *” OR “haplorhini” OR “haplorrhine *” OR “haplorrhini” OR “hominid *” OR “hominin” OR “homininae” OR “hominine” OR “hminines” OR “hominini” OR “hominins” OR “hominoidea” OR “hoolock” OR “howler *” OR “hylobates” OR “hylobatidae” OR “indri” OR “indridae” OR “indriid *” OR “indris” OR “kipunji *” OR “lagothrix” OR “langur” OR “langurs” OR “lemur” OR “lemurid *” OR “lemuriform” OR “lemuriformes” OR “lemuriforms” OR “lemurinae” OR “lemuroidea” OR “lemurs” OR “leontideus” OR “leontocebus” OR “leontopithecus” OR “lepilemur” OR “lepilemurid *” OR “lesula *” OR “lophocebus” OR “loriform” OR “loriformes” OR “lorinae” OR “loris” OR “lorises” OR “lorisid *” OR “lorisiform *” OR “lorisinae” OR “lorisoid *” OR “lutung” OR “lutungs” OR “macaca” OR “macaque’s” OR “macaque” OR “macaques” OR “malbrouck *” OR “mandrill” OR “mandrills” OR “mandrillus” OR “mangabey *” OR “marmoset” OR “marmosets” OR “mico argentatus” OR “mico chrysoleucos” OR “mico emiliae” OR “mico humilis” OR “mico marcai” OR “mico melanurus” OR “mico rondoni” OR “microcebus” OR “miopithecus” OR “mirza coquereli” OR “mirza zaza” OR “monkey” OR “monkeys” OR “muriqui *” OR “nasalis larvatus” OR “nomascus” OR “nycticebus” OR “oedipomidas” OR “orang utan *” OR “orang-utan *” OR “orangutan *” OR “oreonax” OR “otolemur” OR “pan paniscus” OR “pan troglodytes” OR “panin” OR “panina” OR “panins” OR “papio” OR “papionini” OR “paragalago” OR “perodicticinae” OR “perodicticus” OR “phaner” OR “piliocolobus” OR “pithecia” OR “pithecidae” OR “pitheciid *” OR “pitheciinae” OR “pithecinae” OR “platyrhine *” OR “platyrhini” OR “platyrrhina” OR “platyrrhine *” OR “platyrrhini” OR “plecturocebus” OR “pongid *” OR “ponginae” OR “pongo” OR “potto” OR “pottos” OR “presbytini” OR “presbytis” OR “primate” OR “primates” OR “procolobus” OR “prolemur” OR “propithecus” OR “prosimian *” OR “prosimii” OR “pseudopotto” OR “pygathrix” OR “rhinopithecus” OR “rungwecebus” OR “saguinus” OR “saimiri” OR “saimiriinae” OR “sapajus” OR “sciurocheirus” OR “semnopithecus” OR “siamang” OR “siamangs” OR “sifaka” OR “sifakas” OR “simians” OR “simias” OR “simiiform *” OR “strepsir *” OR “surili *” OR “symphalangus” OR “talapoin *” OR “tamarin” OR “tamarins” OR “tamarinus” OR “tarsier” OR “tarsiers” OR “tarsiid *” OR “tarsiiform *” OR “tarsius” OR “theropithecus” OR “trachypithecus” OR “uacari *” OR “uakari” OR “uakaris” OR “varecia” OR “vervet *”)) AND (“laterality” or “handedness” or “hand preference” or “asymmetry”) AND (“Infant” or “longitudinal” or “developmental” or “neonatal” or “neonate”) |
| Web of Science | ((TS = (“allenopithecus” OR “allocebus” OR “alouatta” OR “alouattinae” OR “angwantibo *” OR “anthropoid” OR “anthropoidea” OR “anthropoids” OR “aotes” OR “aotidae” OR “aotinae” OR “aotus” OR “ape” OR “apes” OR “arctocebus” OR “ateles” OR “atelidae” OR “atelinae” OR “avahi” OR “aye-aye *” OR “baboon” OR “baboons” OR “bonobo” OR “bonobos” OR “brachyteles” OR “bushbabies” OR “bushbaby” OR “cacajao” OR “callibella” OR “callicebinae” OR “callicebus” OR “callimico” OR “callithrichid *” OR “callithrichinae” OR “callithrix” OR “callitrichid” OR “callitrichidae” OR “callitrichide” OR “callitrichids” OR “callitrichinae” OR “capuchin” OR “capuchins” OR “carlito syrichta” OR “catarhine *” OR “catarhini” OR “catarrhina” OR “catarrhine *” OR “catarrhini” OR “cebid” OR “cebidae” OR “cebids” OR “cebinae” OR “ceboidea” OR “cebuella” OR “cebus” OR “cephalopachus” OR “cercocebus” OR “cercopithecid *” OR “cercopithecinae” OR “cercopithecine *” OR “cercopithecini” OR “cercopithecoid” OR “cercopithecoidea” OR “cercopithecoids” OR “cercopithecus” OR “cheirogaleidae” OR “cheirogaleus” OR “cheracebus” OR “chimp” OR “chimpanzee” OR “chimpanzees” OR “chimps” OR “chiromyiformes” OR “chiropotes” OR “chlorocebus” OR “colobidae” OR “colobinae” OR “colobine *” OR “colobini” OR “colobus *” OR “cynomolgus” OR “daubentonia” OR “daubentoniidae” OR “douc” OR “doucs” OR “erythrocebus” OR “eulemur” OR “euoticus” OR “euprimate *” OR “galagid *” OR “galago” OR “galagoides” OR “galagonidae” OR “galagos” OR “gelada” OR “geladas” OR “gibbon” OR “gibbons” OR “gorilla” OR “gorillas” OR “grivet” OR “grivets” OR “guenon *” OR “guereza *” OR “hapalemur” OR “haplorhine *” OR “haplorhini” OR “haplorrhine *” OR “haplorrhini” OR “hominid *” OR “hominin” OR “homininae” OR “hominine” OR “hominines” OR “hominini” OR “hominins” OR “hominoidea” OR “hoolock” OR “howler *” OR “hylobates” OR “hylobatidae” OR “indri” OR “indridae” OR “indriid *” OR “indris” OR “kipunji *” OR “lagothrix” OR “langur” OR “langurs” OR “lemur” OR “lemurid *” OR “lemuriform” OR “lemuriformes” OR “lemuriforms” OR “lemurinae” OR “lemuroidea” OR “lemurs” OR “leontideus” OR “leontocebus” OR “leontopithecus” OR “lepilemur” OR “lepilemurid *” OR “lesula *” OR “lophocebus” OR “loriform” OR “loriformes” OR “lorinae” OR “loris” OR “lorises” OR “lorisid *” OR “lorisiform *” OR “lorisinae” OR “lorisoid *” OR “lutung” OR “lutungs” OR “macaca” OR “macaque’s” OR “macaque” OR “macaques” OR “malbrouck *” OR “mandrill” OR “mandrills” OR “mandrillus” OR “mangabey *” OR “marmoset” OR “marmosets” OR “mico argentatus” OR “mico chrysoleucos” OR “mico emiliae” OR “mico humilis” OR “mico marcai” OR “mico melanurus” OR “mico rondoni” OR “microcebus” OR “miopithecus” OR “mirza coquereli” OR “mirza zaza” OR “monkey” OR “monkeys” OR “muriqui *” OR “nasalis larvatus” OR “nomascus” OR “nycticebus” OR “oedipomidas” OR “orang utan *” OR “orang-utan *” OR “orangutan *” OR “oreonax” OR “otolemur” OR “pan paniscus” OR “pan troglodytes” OR “panin” OR “panina” OR “panins” OR “papio” OR “papionini” OR “paragalago” OR “perodicticinae” OR “perodicticus” OR “phaner” OR “piliocolobus” OR “pithecia” OR “pithecidae” OR “pitheciid *” OR “pitheciinae” OR “pithecinae” OR “platyrhine *” OR “platyrhini” OR “platyrrhina” OR “platyrrhine *” OR “platyrrhini” OR “plecturocebus” OR “pongid *” OR “ponginae” OR “pongo” OR “potto” OR “pottos” OR “presbytini” OR “presbytis” OR “primate” OR “primates” OR “procolobus” OR “prolemur” OR “propithecus” OR “prosimian *” OR “prosimii” OR “pseudopotto” OR “pygathrix” OR “rhinopithecus” OR “rungwecebus” OR “saguinus” OR “saimiri” OR “saimiriinae” OR “sapajus” OR “sciurocheirus” OR “semnopithecus” OR “siamang” OR “siamangs” OR “sifaka” OR “sifakas” OR “simians” OR “simias” OR “simiiform *” OR “strepsir *” OR “surili *” OR “symphalangus” OR “talapoin *” OR “tamarin” OR “tamarins” OR “tamarinus” OR “tarsier” OR “tarsiers” OR “tarsiid *” OR “tarsiiform *” OR “tarsius” OR “theropithecus” OR “trachypithecus” OR “uacari *” OR “uakari” OR “uakaris” OR “varecia” OR “vervet *”)) AND ALL = (“laterality” or “handedness” or “hand preference” or “asymmetryy”)) AND ALL = (“Infant” or “longitudinal” or “developmental” or “neonatal” or “neonate”) |
| Identification of Studies via Databases and Registers |
|---|
| Topic not measured (n = 6) |
| Ss’ ages not reported (n = 3) |
| No Ss < 1 year old (n = 10) |
| Ss > 1 year old (n = 5) |
| Identification of Studies via Other Methods |
Data for Ss < 1 year old not separated out (n = 1)
|
Paper not in English (n = 1)
|
References
- MacNeilage, P.F.; Studdert-Kennedy, M.G.; Lindblom, B. Primate handedness reconsidered. Behav. Brain Sci. 1987, 10, 247–263. [Google Scholar] [CrossRef]
- Fagot, J.; Vauclair, J. Manual laterality in nonhuman primates: A distinction between handedness and manual specialization. Psychol. Bull. 1991, 109, 76. [Google Scholar] [CrossRef] [PubMed]
- Papademetriou, E.; Sheu, C.F.; Michel, G.F. A meta-analysis of primate hand preferences, particularly for reaching. J. Comp. Psychol. 2005, 119, 33–48. [Google Scholar] [CrossRef] [PubMed]
- McGrew, W.; Marchant, L. On the other hand: Current issues in and meta-analysis of the behavioral laterality of hand function in nonhuman primates. Am. J. Phys. Anthropol. 1997, 104, 201–232. [Google Scholar] [CrossRef]
- Caspar, K.R.; Pallasdies, F.; Mader, L.; Sartorelli, H.; Begall, S. The evolution and biological correlates of hand preferences in anthropoid primates. Elife 2022, 11, e77875. [Google Scholar] [CrossRef]
- Soto, C.; Gázquez, J.M.; Llorente, M. Hand preferences in coordinated bimanual tasks in non-human primates: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2022, 141, 104822. [Google Scholar] [CrossRef]
- Fitch, W.T.; Braccini, S.N. Primate laterality and the biology and evolution of human handedness: A review and synthesis. Ann. N. Y. Acad. Sci. 2013, 1288, 70–85. [Google Scholar] [CrossRef]
- Papadatou-Pastou, M.; Ntolka, E.; Schmitz, J.; Martin, M.; Munafo, M.R.; Ocklenburg, S.; Paracchini, S. Human handedness: A meta-analysis. Psychol. Bull. 2020, 146, 481–524. [Google Scholar] [CrossRef] [PubMed]
- Michel, G.F.; Babik, I.; Sheu, C.F.; Campbell, J.M. Latent classes in the developmental trajectories of infant handedness. Dev. Psychol. 2014, 50, 349–359. [Google Scholar] [CrossRef]
- Campbell, J.M.; Marcinowski, E.C.; Michel, G.F. The development of neuromotor skills and hand preference during infancy. Dev. Psychobiol. 2018, 60, 165–175. [Google Scholar] [CrossRef]
- Michel, G.F. Development of infant handedness. In Conceptions of Development: Lessons from the Laboratory; Lewkowicz, D.J., Lickliter, R., Eds.; Psychology Press: New York, NY, USA, 2002; pp. 165–186. [Google Scholar]
- Michel, G.F. Handedness Development: A Model for Investigating the Development of Hemispheric Specialization and Interhemispheric Coordination. Symmetry 2021, 13, 992. [Google Scholar] [CrossRef]
- Michel, G.F.; Nelson, E.L.; Babik, I.; Campbell, J.M.; Marcinowski, E.C. Multiple trajectories in the developmental psychobiology of human handedness. Adv. Child Dev. Behav. 2013, 45, 227–260. [Google Scholar] [CrossRef]
- Nelson, E.L. Developmental cascades as a framework for primate handedness. Front. Behav. Neurosci. 2022, 16, 1063348. [Google Scholar] [CrossRef] [PubMed]
- Boulinguez-Ambroise, G.; Aychet, J.; Pouydebat, E. Limb Preference in Animals: New Insights into the Evolution of Manual Laterality in Hominids. Symmetry 2022, 14, 96. [Google Scholar] [CrossRef]
- Nelson, E.L. Insights Into Human and Nonhuman Primate Handedness From Measuring Both Hands. Curr. Dir. Psychol. Sci. 2022, 31, 154–161. [Google Scholar] [CrossRef]
- Rogers, L.J. Development of Hand and Paw Preferences and Their Association with Other Patterns of Behaviour and Cognition. Symmetry 2023, 15, 926. [Google Scholar] [CrossRef]
- Nelson, E.; Alvarez, J.; Jimenez, B.; Padron, K. State of the Field. In Primate Cognitive Studies; Schwartz, B.L., Beran, M.J., Eds.; Cambridge University Press: Cambridge, UK, 2022; pp. 88–114. [Google Scholar]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Syst. Rev. 2021, 10, 1–11. [Google Scholar] [CrossRef]
- Moher, D.; Shamseer, L.; Clarke, M.; Ghersi, D.; Liberati, A.; Petticrew, M.; Shekelle, P.; Stewart, L.A. Preferred reporting items for systematic review and meta-analysis protocols (PRISMA-P) 2015 statement. Syst. Rev. 2015, 4, 1–9. [Google Scholar] [CrossRef]
- Shamseer, L.; Moher, D.; Clarke, M.; Ghersi, D.; Liberati, A.; Petticrew, M.; Shekelle, P.; Stewart, L.A. Preferred reporting items for systematic review and meta-analysis protocols (PRISMA-P) 2015: Elaboration and explanation. Bmj 2015, 349, g7647. [Google Scholar] [CrossRef]
- Nelson, E.L.; Karimi, A. Behavioral Laterality across the First Year of Life in Nonhuman Primates [Systematic Review Protocol]; OSF: Galesburg, IL, USA, 2023. [Google Scholar] [CrossRef]
- Cassidy, L.C.; Leenaars, C.H.C.; Rincon, A.V.; Pfefferle, D. Comprehensive search filters for retrieving publications on nonhuman primates for literature reviews (filterNHP). Am. J. Primatol. 2021, 83, e23287. [Google Scholar] [CrossRef]
- The EndNote Team. EndNote; EndNote 20; Clarivate: Philadelphia, PA, USA, 2013. [Google Scholar]
- Wallace, B.C.; Small, K.; Brodley, C.E.; Lau, J.; Trikalinos, T.A. Deploying an interactive machine learning system in an evidence-based practice center: Abstrackr. In Proceedings of the 2nd ACM SIGHIT International Health Informatics Symposium, Miami, FL, USA, 28–30 January 2012; pp. 819–824. [Google Scholar] [CrossRef]
- Hook, M.A.; Rogers, L.J. Development of hand preferences in marmosets (Callithrix jacchus) and effects of aging. J. Comp. Psychol. 2000, 114, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Matoba, M.; Masataka, N.; Tanioka, Y. Cross-generational continuity of hand-use preferences in marmosets. Behaviour 1991, 117, 281–286. [Google Scholar] [CrossRef]
- Rogers, L.J.; Kaplan, G. Teat preference for suckling in common marmosets: Relationship to side of being carried and hand preference. Laterality Asymmetries Body Brain Cogn. 1998, 3, 269–281. [Google Scholar] [CrossRef]
- Guerra, R.F.; da Silveira, N.; Bernardi, N.; Legal, E.J. Hand preference during behavioral tests and spontaneous activity in two species of common marmoset (Callithrix jacchus and Callithrix penicillata). Rev. Bras. Biol. 1997, 57, 563–570. [Google Scholar]
- Westergaard, G.C.; Byrne, G.; Suomi, S.J. Handedness and cortisol in tufted capuchin monkey infants. Dev. Psychobiol. J. Int. Soc. Dev. Psychobiol. 2000, 36, 213–217. [Google Scholar] [CrossRef]
- Westergaard, G.C.; Byrne, G.; Suomi, S.J. Early lateral bias in tufted capuchins (Cebus apella). Dev. Psychobiol. 1998, 32, 45–50. [Google Scholar] [CrossRef]
- Tanaka, I. Parity-related differences in suckling behavior and nipple preference among free-ranging Japanese macaques. Am. J. Primatol. 1997, 42, 331–339. [Google Scholar] [CrossRef]
- Ôta, K.; Makino, Y.; Kimura, M.; Suzuki, J. Lactation in the Japanese monkey (Macaca fuscata): Yield and composition of milk and nipple preference of young. Primates 1991, 32, 35–48. [Google Scholar] [CrossRef]
- Tanaka, I. Change of nipple preference between successive offspring in Japanese macaques. Am. J. Primatol. 1989, 18, 321–325. [Google Scholar] [CrossRef]
- Nakamichi, M. Development of infant twin Japanese monkeys (Macaca fuscata) in a free-ranging group. Primates 1983, 24, 576–583. [Google Scholar] [CrossRef]
- Hiraiwa, M. Maternal and alloparental care in a troop of free-ranging Japanese monkeys. Primates 1981, 22, 309–329. [Google Scholar] [CrossRef]
- Jaffe, B.D.; Evans, T.A.; Howell, S.; Westergaard, G.C.; Snoy, P.J.; Higley, J.D. Left versus right nipple preference in free-ranging infant rhesus macaques (Macaca mulatta). Dev. Psychobiol. 2006, 48, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Tomaszycki, M.; Cline, C.; Griffin, B.; Maestripieri, D.; Hopkins, W.D. Maternal cradling and infant nipple preferences in rhesus monkeys (Macaca mulatta). Dev. Psychobiol. 1998, 32, 305–312. [Google Scholar] [CrossRef]
- Lindburg, D.G. The rhesus monkey in North India: An ecological and behavioral study. In Primate Behavior: Developments in Field and Laboratory Research; Rosenblum, L., Ed.; Academic Press: New York, NY, USA, 1971; Volume 2, pp. 1–106. [Google Scholar]
- Deets, A.C.; Harlow, H.F. Nipple preferences in nursing singleton-and twin-reared rhesus monkey infants. Dev. Psychol. 1970, 2, 159. [Google Scholar] [CrossRef]
- Spencer-Booth, Y. The behaviour of twin rhesus monkeys and comparisons with the behaviour of single infants. Primates 1968, 9, 75–84. [Google Scholar] [CrossRef]
- Hinde, R.; Bowell, T.; Spencer-Booth, Y. Behaviour of socially living rhesus monkeys in their first six months. Proc. Zool. Soc. Lond. 1964, 143, 609–649. [Google Scholar]
- Nelson, E.L.; Konidaris, G.D.; Berthier, N.E.; Braun, M.C.; Novak, M.F.; Suomi, S.J.; Novak, M.A. Kinematics of reaching and implications for handedness in rhesus monkey infants. Dev. Psychobiol. 2012, 54, 460–467. [Google Scholar] [CrossRef]
- Westergaard, G.; Champoux, M.; Suomi, S. Plasma cortisol is associated with handedness in infant rhesus monkeys. Dev. Psychobiol. J. Int. Soc. Dev. Psychobiol. 2001, 38, 116–122. [Google Scholar] [CrossRef]
- Westergaard, G.C.; Champoux, M.; Suomi, S.J. Hand preference in infant rhesus macaques (Macaca mulatta). Child. Dev. 1997, 68, 387–393. [Google Scholar] [CrossRef]
- Hauser, M.D.; Andersson, K. Left hemisphere dominance for processing vocalizations in adult, but not infant, rhesus monkeys: Field experiments. Proc. Natl. Acad. Sci. USA 1994, 91, 3946–3948. [Google Scholar] [CrossRef] [PubMed]
- Hauser, M.D.; Akre, K. Asymmetries in the timing of facial and vocal expressions by rhesus monkeys: Implications for hemispheric specialization. Anim. Behav. 2001, 61, 391–400. [Google Scholar] [CrossRef]
- Nelson, E.L.; Emery, M.S.; Babcock, S.M.; Novak, M.F.; Suomi, S.J.; Novak, M.A. Head orientation and handedness trajectory in rhesus monkey infants (Macaca mulatta). Dev. Psychobiol. 2011, 53, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Garber, P.A.; Tian, J.; Lu, J. Neonatal nipple preference and maternal cradling laterality in wild Taihangshan macaques (Macaca mulatta tcheliensis). Am. J. Primatol. 2020, 82, e23197. [Google Scholar] [CrossRef] [PubMed]
- Erwin, J.; Anderson, B.; Bunger, D. Nursing behavior of infant pigtail monkeys (Macaca nemestrina): Preferences for nipples. Percept. Mot. Ski. 1975, 40, 592–594. [Google Scholar] [CrossRef]
- Regaiolli, B.; Spiezio, C.; Hopkins, W.D. Asymmetries in mother-infant behaviour in Barbary macaques (Macaca sylvanus). PeerJ 2018, 6, e4736. [Google Scholar] [CrossRef] [PubMed]
- Boulinguez-Ambroise, G.; Pouydebat, E.; Disarbois, E.; Meguerditchian, A. Maternal cradling bias in baboons: The first environmental factor affecting early infant handedness development? Dev. Sci. 2022, 25, e13179. [Google Scholar] [CrossRef]
- Damerose, E.; Hopkins, W.D. Scan and focal sampling: Reliability in the laterality for maternal cradling and infant nipple preferences in olive baboons, Papio anubis. Anim. Behav. 2002, 63, 511–518. [Google Scholar] [CrossRef][Green Version]
- Westergaard, G.C. Hand preference in the use of tools by infant baboons (Papio cynocephalus anubis). Percept. Mot. Ski. 1993, 76, 447–450. [Google Scholar] [CrossRef]
- Fagot, J. Ontogeny of object manipulation and manual lateralization in the Guinea baboon: Preliminary observations. In Primate Laterality Current Behavioral Evidence of Primate Asymmetries; Springer: Berlin, Germany, 1993; pp. 235–250. [Google Scholar]
- Winkler, P.; Wrogemann, D.; Prestel, H. Twins in free-ranging Hanuman langurs (Presbytis entellus). Primates 1989, 30, 255–259. [Google Scholar] [CrossRef]
- Horwich, R.H. Development of behaviors in a male spectacled langur (Presbytis obscurus). Primates 1974, 15, 151–178. [Google Scholar] [CrossRef]
- Zhao, D.; Gao, X.; Li, B.; Watanabe, K. First wild evidence of neonate nipple preference and maternal cradling laterality in Old World monkeys: A preliminary study from Rhinopithecus roxellana. Behav. Process. 2008, 77, 364–368. [Google Scholar] [CrossRef]
- Hopkins, W.D.; De Lathouwers, M. Left nipple preferences in infant Pan paniscus and P. troglodytes. Int. J. Primatol. 2006, 27, 1653–1662. [Google Scholar] [CrossRef][Green Version]
- Dienske, H.; Hopkins, B.; Reid, A.K. Lateralisation of infant holding in chimpanzees: New data do not confirm previous findings. Behaviour 1995, 132, 801–809. [Google Scholar] [CrossRef]
- Nishida, T. Left nipple suckling preference in wild chimpanzees. Ethol. Sociobiol. 1993, 14, 45–51. [Google Scholar] [CrossRef]
- Hopkins, W.D.; Bard, K.A. Asymmetries in spontaneous head orientation in infant chimpanzees (Pan troglodytes). Behav. Neurosci. 1995, 109, 808–812. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hopkins, W.D.; Bard, K.A. Hemispheric specialization in infant chimpanzees (Pan troglodytes): Evidence for a relation with gender and arousal. Dev. Psychobiol. J. Int. Soc. Dev. Psychobiol. 1993, 26, 219–235. [Google Scholar] [CrossRef]
- Bard, K.A.; Hopkins, W.D.; Fort, C.L. Lateral bias in infant chimpanzees (Pan troglodytes). J. Comp. Psychol. 1990, 104, 309. [Google Scholar] [CrossRef]
- Hopkins, W.D.; Bard, K.A.; Griner, K.M. Locomotor adaptation and leading limb asymmetries in neonatal chimpanzees (Pan troglodytes). Int. J. Primatol. 1997, 18, 105–114. [Google Scholar] [CrossRef]
- Brésard, B.; Bresson, F. Handedness in Pongo pygmaeus and Pan troglodytes. J. Hum. Evol. 1983, 12, 659–666. [Google Scholar] [CrossRef]
- Fagot, J.; Bard, K.A. Asymmetric grasping response in neonate chimpanzees (Pan troglodytes). Infant Behav. Dev. 1995, 18, 253–255. [Google Scholar] [CrossRef]
- Hopkins, W.D.; Bard, K.A. A longitudinal study of hand preference in chimpanzees (Pan troglodytes). Dev. Psychobiol. 2000, 36, 292–300. [Google Scholar] [CrossRef]
- Chorazyna, H. Shifts in laterality in a baby chimpanzee. Neuropsychologia 1976, 14, 381–384. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.D.; Parr, L.A. Lateralized behavior and lymphocyte counts in chimpanzees (Pan troglodytes): A cross-sectional and longitudinal assessment. Dev. Neuropsychol. 1998, 14, 519–533. [Google Scholar] [CrossRef]
- Bard, K.A. Neonatal neurobehavioral correlates of lateral bias and affect in infant chimpanzees (Pan troglodytes). Dev. Neuropsychol. 1998, 14, 471–494. [Google Scholar] [CrossRef]
- Cunningham, D.; Forsythe, C.; Ward, J.P. A report of behavioral lateralization in an infant orang-utan (Pongo pygmaeus). Primates 1989, 30, 249–253. [Google Scholar] [CrossRef]
- Needham, A.W.; Nelson, E.L. How babies use their hands to learn about objects: Exploration, reach-to-grasp, manipulation, and tool use. WIREs Cogn. Sci. 2023, e1661. [Google Scholar] [CrossRef] [PubMed]
- Fagard, J.; Pezé, A. Age Changes in Interlimb Coupling and the Development of Bimanual Coordination. J. Mot. Behav. 1997, 29, 199–208. [Google Scholar] [CrossRef]
- Michel, G.F. Right-handedness: A consequence of infant supine head-orientation preference? Science 1981, 212, 685–687. [Google Scholar] [CrossRef]
- Michel, G.F.; Harkins, D.A. Postural and lateral asymmetries in the ontogeny of handedness during infancy. Dev. Psychobiol. 1986, 19, 247–258. [Google Scholar] [CrossRef]
- Konishi, Y.; Kuriyama, M.; Mikawa, H.; Suzuki, J. Effect of body position on later postural and functional lateralities of preterm infants. Dev. Med. Child Neurol. 1987, 29, 751–756. [Google Scholar] [CrossRef]
- Corbetta, D.; Thelen, E. Lateral biases and fluctuations in infants’ spontaneous arm movements and reaching. Dev. Psychobiol. 1999, 34, 237–255. [Google Scholar] [CrossRef]
- Boulinguez-Ambroise, G.; Pouydebat, E.; Disarbois, É.; Meguerditchian, A. Human-like maternal left-cradling bias in monkeys is altered by social pressure. Sci. Rep. 2020, 10, 11036. [Google Scholar] [CrossRef] [PubMed]
- Bailoo, J.D.; Quinius, B.; Schapiro, S.J.; Hopkins, W.D.; Bennett, A.J. Freestanding bipedal posture and coordinated bimanual manipulation significantly influence lateralized hand use in rhesus monkeys (Macaca mulatta) and chimpanzees (Pan troglodytes). J. Comp. Psychol. 2019, 133, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Dettmer, A.M.; Ruggiero, A.M.; Novak, M.A.; Meyer, J.S.; Suomi, S.J. Surrogate mobility and orientation affect the early neurobehavioral development of infant rhesus macaques (Macaca mulatta). Dev. Psychobiol. 2008, 50, 418–422. [Google Scholar] [CrossRef] [PubMed]
- van der Meer, A.L. Keeping the arm in the limelight: Advanced visual control of arm movements in neonates. Eur. J. Paediatr. Neurol. 1997, 1, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Gunderson, V.M.; Sackett, G.P. Development of pattern recognition in infant pigtailed macaques (Macaca nemestrina). Dev. Psychol. 1984, 20, 418–426. [Google Scholar] [CrossRef]
- Hopkins, W.D. Neuroanatomical asymmetries in nonhuman primates in the homologs to Broca's and Wernicke’s areas: A mini-review. Emerg. Top. Life Sci. 2022, 6, 271–284. [Google Scholar] [CrossRef]
- Manning, J.; Heaton, R.; Chamberlain, A. Left-side cradling: Similarities and differences between apes and humans. J. Hum. Evol. 1994, 26, 77–83. [Google Scholar] [CrossRef]
- Trehub, S.E.; Corter, C.M.; Shosenberg, N. Neonatal reflexes: A search for lateral asymmetries. In Manual Specialization and the Developing Brain; Young, G., Segalowitz, S.J., Corter, C.M., Trehub, S.E., Eds.; Academic Press: San Diego, CA, USA, 1983; pp. 257–274. [Google Scholar]
- Michel, G.F.; Goodwin, R. Intrauterine birth position predicts newborn supine head position preferences. Infant Behav. Dev. 1979, 2, 29–38. [Google Scholar] [CrossRef]
- Hepper, P.G.; Wells, D.L.; Lynch, C. Prenatal thumb sucking is related to postnatal handedness. Neuropsychologia 2005, 43, 313–315. [Google Scholar] [CrossRef]
- Previc, F.H. A general theory concerning the prenatal origins of cerebral lateralization in humans. Psychol. Rev. 1991, 98, 299–334. [Google Scholar] [CrossRef]
- Roberts, V.H.; Castro, J.N.; Wessel, B.M.; Conrad, D.F.; Lewis, A.D.; Lo, J.O. Rhesus macaque fetal and placental growth demographics: A resource for laboratory animal researchers. Am. J. Primatol. 2023, e23526. [Google Scholar] [CrossRef] [PubMed]
- ManyPrimates; Altschul, D.M.; Beran, M.J.; Bohn, M.; Caspar, K.R.; Fichtel, C.; Försterling, M.; Grebe, N.; Hernandez-Aguilar, R.; Kwok, S.C.; et al. Collaborative open science as a way to reproducibility and new insights in primate cognition research. Jpn. Psychol. Rev. 2019, 62, 205–220. [Google Scholar] [CrossRef]
- Becker, Y.; Sein, J.; Velly, L.; Giacomino, L.; Renaud, L.; Lacoste, R.; Anton, J.-L.; Nazarian, B.; Berne, C.; Meguerditchian, A. Early Left-Planum Temporale Asymmetry in newborn monkeys (Papio anubis): A longitudinal structural MRI study at two stages of development. NeuroImage 2021, 227, 117575. [Google Scholar] [CrossRef] [PubMed]
- Becker, Y.; Phelipon, R.; Sein, J.; Velly, L.; Renaud, L.; Meguerditchian, A. Planum temporale grey matter volume asymmetries in newborn monkeys (Papio anubis). Brain Struct. Funct. 2022, 227, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Vingerhoets, G.; Verhelst, H.; Gerrits, R.; Badcock, N.; Bishop, D.V.; Carey, D.; Flindall, J.; Grimshaw, G.; Harris, L.J.; Hausmann, M. Laterality indices consensus initiative (LICI): A Delphi expert survey report on recommendations to record, assess, and report asymmetry in human behavioural and brain research. Laterality 2023, 28, 122–191. [Google Scholar] [CrossRef]
- Van Lawick-Goodall, J. Some aspects of mother-infant relationships in a group of wild chimpanzees. In The Origins of Human Social Relations; Academic Press: Oxford, UK, 1971; p. 297. [Google Scholar]
- Kubota, K. Learning of a hiding task and a delayed response task in infant rhesus monkeys. Neurosci. Res. 1994, 18, 301–313. [Google Scholar] [CrossRef]
- Corp, N.; Byrne, R.W. The ontogeny of manual skill in wild chimpanzees: Evidence from feeding on the fruit of Saba florida. Behaviour 2002, 139, 137–168. [Google Scholar] [CrossRef]
- Rossano, F. Sequence organization and timing of bonobo mother-infant interactions. Interact. Stud. Soc. Behav. Commun. Biol. Artif. Syst. 2013, 14, 160–189. [Google Scholar] [CrossRef]
- Sakamoto, K.; Sawada, K.; Fukunishi, K.; Noritaka, I.; Sakata-Haga, H.; Yoshihiro, F. Postnatal change in sulcal length asymmetry in cerebrum of cynomolgus monkeys (Macaca fascicularis). Anat. Rec. 2014, 297, 200–207. [Google Scholar] [CrossRef]
- Scott, J.A.; Grayson, D.; Fletcher, E.; Lee, A.; Bauman, M.D.; Schumann, C.M.; Buonocore, M.H.; Amaral, D.G. Longitudinal analysis of the developing rhesus monkey brain using magnetic resonance imaging: Birth to adulthood. Brain Struct. Funct. 2016, 221, 2847–2871. [Google Scholar] [CrossRef]
- Lehman, R.A. Distribution and changes in strength of hand preference of cynomolgus monkeys. Brain Behav. Evol. 1980, 17, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Brooker, R.J.; Lehman, R.A.; Heimbuch, R.C.; Kidd, K.K. Hand usage in a colony of bonnet monkeys, Macaca radiata. Behav. Genet. 1981, 11, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.D.; Wesley, M.J.; Russell, J.L.; Schapiro, S.J. Parental and perinatal factors influencing the development of handedness in captive chimpanzees. Dev. Psychobiol. 2006, 48, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.D.; Bard, K.A.; Jones, A.; Bales, S.L. Chimpanzee Hand Preference in Throwing and Infant Cradling: Implications for the Origin of Human Handedness. Curr. Anthropol. 1993, 34, 786–790. [Google Scholar] [CrossRef]
- Manning, J.T.; Denman, J. Lateral cradling preferences in humans (Homo sapiens): Similarities within families. J. Comp. Psychol. 1994, 108, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.D.; Leavens, D.A. Hand use and gestural communication in chimpanzees (Pan troglodytes). J. Comp. Psychol. 1998, 112, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Carriba, S.; Loeches, A.; Morcillo, A.; Hopkins, W.D. Asymmetry in facial expression of emotions by chimpanzees. Neuropsychologia 2002, 40, 1523–1533. [Google Scholar] [CrossRef]
- Fletcher, A.W.; Weghorst, J.A. Laterality of hand function in naturalistically housed chimpanzees (Pan troglodytes). Laterality 2005, 10, 219–242. [Google Scholar] [CrossRef]
- Lyn, H.; Pierre, P.; Bennett, A.J.; Fears, S.; Woods, R.; Hopkins, W.D. Planum temporale grey matter asymmetries in chimpanzees (Pan troglodytes), vervet (Chlorocebus aethiops sabaeus), rhesus (Macaca mulatta) and bonnet (Macaca radiata) monkeys. Neuropsychologia 2011, 49, 2004–2012. [Google Scholar] [CrossRef]
- Bourjade, M.; Meunier, H.; Blois-Heulin, C.; Vauclair, J. Baboons’ hand preference resists to spatial factors for a communicative gesture but not for a simple manipulative action. Dev. Psychobiol. 2013, 55, 651–661. [Google Scholar] [CrossRef]
- Hopkins, W.D.; Reamer, L.; Mareno, M.C.; Schapiro, S.J. Genetic basis in motor skill and hand preference for tool use in chimpanzees (Pan troglodytes). Proc. Biol. Sci. 2015, 282, 20141223. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.D.; Meguerditchian, A.; Coulon, O.; Misiura, M.; Pope, S.; Mareno, M.C.; Schapiro, S.J. Motor skill for tool-use is associated with asymmetries in Broca’s area and the motor hand area of the precentral gyrus in chimpanzees (Pan troglodytes). Behav. Brain Res. 2017, 318, 71–81. [Google Scholar] [CrossRef]
- Rogers, L.J. Differential Ageing of the Brain Hemispheres: Evidence from a Longitudinal Study of Hand Preferences in Common Marmosets. Symmetry 2021, 13, 2349. [Google Scholar] [CrossRef]
- Drea, C.M.; Wallen, K.; Akinbami, M.A.; Mann, D.R. Neonatal testosterone and handedness in yearling rhesus monkeys (Macaca mulatta). Physiol. Behav. 1995, 58, 1257–1262. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.D. Hand preferences for simple reaching in juvenile chimpanzees (Pan troglodytes): Continuity in development. Dev. Psychol. 1995, 31, 619–625. [Google Scholar] [CrossRef]
- Westergaard, G.C.; Suomi, S.J. Lateral bias in capuchin monkeys (Cebus apella): Concordance between parents and offspring. Dev. Psychobiol. 1997, 31, 143–147. [Google Scholar] [CrossRef]
- Morino, L.; Uchikoshi, M.; Bercovitch, F.; Hopkins, W.D.; Matsuzawa, T. Tube task hand preference in captive hylobatids. Primates 2017, 58, 403–412. [Google Scholar] [CrossRef]
- Fu, W.-W.; Ren, Y.; Wang, C.-L.; Wang, X.-W.; Li, B.-G. Hand Preference in Rhinopithecus roxellana Infants: Is It Influenced by Familial Inheritance? Symmetry 2020, 12, 1905. [Google Scholar] [CrossRef]
- Vauclair, J.; Fagot, J. Spontaneous hand usage and handedness in a troop of baboons. Cortex 1987, 23, 265–274. [Google Scholar] [CrossRef]
- Gao, X.; Guo, S.; Hu, Y.; Li, B.; Qi, X. Maternal cradling laterality and neonate nipple preference in the Sichuan snub-nosed monkey (Rhinopithecus roxellana) in Qinling Mountains. Acta Theriol. Sinica 2010, 30, 133–138. [Google Scholar]


| Source | Species | N | Setting | Behavior(s) | Developmental Quality | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Q1 | Q2 | Q3 | Q4 | Q5 | Total | |||||
| Platyrrhine Monkeys | ||||||||||
| Hook and Rogers [26] | C. j. jacchus | 15 * | Captive | 2 | 1 | 1 | 1 | 1 | 1 | 5 |
| Matoba, et al. [27] | C. j. jacchus | 23 | Captive | 2 | 1 | 0 | 0 | 0 | 1 | 2 |
| Rogers and Kaplan [28] | C. j. jacchus | 15 * | Captive | 1, 2 | 1 | 1 | 1 | 1 | 1 | 5 |
| Guerra, et al. [29] | C. pencillata | 2 | Captive | 2 | 0 | 1 | 1 | 0 | 1 | 3 |
| Westergaard, et al. [30] | C. apella | 14 | Captive | 2 | 1 | 1 | 1 | 1 | 1 | 5 |
| Westergaard, et al. [31] | C. apella | 16 | Captive | 2, 3 | 1 | 1 | 1 | 1 | 1 | 5 |
| Catarrhine Monkeys | ||||||||||
| Tanaka [32] | M. fuscata | 63 | Wild | 1 | 0 | 1 | 1 | 1 | 1 | 4 |
| Ôta, et al. [33] | M. fuscata | 3 | Captive | 1 | 1 | 1 | 1 | 0 | 0 | 3 |
| Tanaka [34] | M. fuscata | 40 | Wild | 1 | 1 | 1 | 1 | 0 | 1 | 4 |
| Nakamichi [35] | M. fuscata | 2 * | Free ranging | 1 | 0 | 1 | 1 | 0 | 0 | 2 |
| Hiraiwa [36] | M. fuscata | 16 | Wild | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Jaffe, et al. [37] | M. mulatta | 64 | Free ranging | 1 | 1 | 1 | 1 | 1 | 1 | 5 |
| Tomaszycki, et al. [38] | M. mulatta | 41 | Captive | 1 | 1 | 1 | 1 | 1 | 1 | 5 |
| Lindburg [39] | M. mulatta | NR | Wild | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Deets and Harlow [40] | M. mulatta | 12 * | Captive | 1 | 0 | 1 | 1 | 1 | 0 | 3 |
| Spencer-Booth [41] | M. mulatta | 2 * | Captive | 1 | 0 | 1 | 1 | 0 | 0 | 2 |
| Hinde, et al. [42] | M. mulatta | 3 | Captive | 1 | 0 | 1 | 1 | 0 | 0 | 2 |
| Nelson, et al. [43] | M. mulatta | 12 | Captive | 2 | 1 | 0 | 0 | 0 | 0 | 1 |
| Westergaard, et al. [44] | M. mulatta | 18 | Captive | 2 | 1 | 0 | 0 | 0 | 1 | 2 |
| Westergaard, et al. [45] | M. mulatta | 20 | Captive | 2 | 1 | 0 | 0 | 1 | 1 | 3 |
| Hauser and Andersson [46] | M. mulatta | 37 | Free ranging | 6 | 1 | 0 | 0 | 0 | 1 | 2 |
| Hauser and Akre [47] | M. mulatta | 55 | Free ranging | 8 | 1 | 0 | 0 | 1 | 1 | 3 |
| Nelson, et al. [48] | M. mulatta | 16 | Captive | 2, 3, 4 | 1 | 1 | 1 | 1 | 1 | 5 |
| Guo, et al. [49] | M. m. tcheliensis | 9 | Wild | 1 | 1 | 1 | 1 | 1 | 1 | 5 |
| Erwin, et al. [50] | M. nemestrina | 56 | Captive | 1 | 1 | 0 | 0 | 0 | 1 | 2 |
| Regaiolli, et al. [51] | M. sylvanus | 7 | Captive | 1, 2, 8 | 1 | 0 | 0 | 0 | 1 | 2 |
| Boulinguez-Ambroise, et al. [52] | P. anubis | 41 | Captive | 2 | 1 | 1 | 1 | 1 | 1 | 5 |
| Damerose and Hopkins [53] | P. anubis | 10 | Captive | 1, 3 | 0 | 0 | 1 | 0 | 0 | 1 |
| Westergaard [54] | P. c. anubis | 5 | Captive | 2 | 0 | 0 | 0 | 0 | 1 | 1 |
| Fagot [55] | P. papio | 4 | Captive | 1, 2 | 1 | 1 | 1 | 1 | 1 | 5 |
| Winkler, et al. [56] | P. entellus | 4 * | Wild | 1 | 0 | 1 | 1 | 0 | 1 | 3 |
| Horwich [57] | P. obscurus | 1 | Captive | 1 | 0 | 1 | 1 | 0 | 0 | 2 |
| Zhao, et al. [58] | R. roxellana | 10 | Wild | 1 | 1 | 0 | 0 | 0 | 1 | 2 |
| Apes | ||||||||||
| Hopkins and De Lathouwers [59] | P. paniscus | 3 | Captive | 1 | 1 | 1 | 1 | 1 | 1 | 5 |
| Hopkins and De Lathouwers [59] | P. troglodytes | 3 | Captive | 1 | 1 | 1 | 1 | 1 | 1 | 5 |
| Dienske, et al. [60] | P. troglodytes | 5 | Captive | 1 | 0 | 0 | 0 | 0 | 1 | 1 |
| Nishida [61] | P. troglodytes | 10 | Wild | 1 | 0 | 1 | 1 | 0 | 0 | 2 |
| Hopkins and Bard [62] | P. troglodytes | 43 | Captive | 3 | 1 | 0 | 0 | 0 | 1 | 2 |
| Hopkins and Bard [63] | P. troglodytes | 31 | Captive | 4 | 1 | 0 | 1 | 1 | 1 | 4 |
| Bard, et al. [64] | P. troglodytes | 12 | Captive | 4 | 1 | 1 | 1 | 1 | 1 | 5 |
| Hopkins, et al. [65] | P. troglodytes | 13 | Captive | 5 | 1 | 0 | 0 | 0 | 1 | 2 |
| Brésard and Bresson [66] | P. troglodytes | 1 | Captive | 2, 5 | 0 | 1 | 1 | 0 | 1 | 3 |
| Fagot and Bard [67] | P. troglodytes | 13 | Captive | 2, 7 | 1 | 0 | 0 | 0 | 1 | 2 |
| Hopkins and Bard [68] | P. troglodytes | 53 | Captive | 2, 3, 4 | 1 | 0 | 1 | 0 | 1 | 3 |
| Chorazyna [69] | P. troglodytes | 1 | Captive | 2, 5, 8 | 1 | 1 | 1 | 0 | 1 | 4 |
| Hopkins and Parr [70] | P. troglodytes | 42 | Captive | 3, 4, 5 | 1 | 0 | 1 | 1 | 1 | 4 |
| Bard [71] | P. troglodytes | 37 | Captive | 2, 3, 4, 5 | 1 | 0 | 1 | 1 | 1 | 4 |
| Cunningham, et al. [72] | P. pygmaeus | 1 | Captive | 2, 5 | 1 | 1 | 1 | 0 | 1 | 4 |
| Species | Developmental Outcomes | Source | |
|---|---|---|---|
| C. j. jacchus | −/+ | Preferences developed in week 1, with stable patterns through month 2 | Rogers and Kaplan [28] * |
| ~ | No links to hand preference measured at 18 months | Rogers and Kaplan [28] * | |
| M. fuscata | −/+ | No preference in the first week of life; established by week 2 | Tanaka [34] †; Tanaka [32] |
| −/+ | No preference in the first two weeks of life; established in weeks 2–4 | Hiraiwa [36] | |
| −/+ | Preferences established no later than week 8 (earliest timing not given) | Ôta, et al. [33] | |
| = | Preference by each twin for opposite nipples (no timing given) | Nakamichi [35] | |
| ~ | Not tested in this species by any author(s) | ||
| M. mulatta | −/+ | Preferences established between 48 h and 2 weeks of life, increased in strength until 3 months, and then remained stable | Jaffe, et al. [37] |
| +/− | Preferences observed in weeks 1, 2, and 3, but not 4, 5, or 6 | Tomaszycki, et al. [38] *,† | |
| + | Preferences observed at weeks 2–4 matched preferences at weeks 12–14 | Deets and Harlow [40] | |
| −/+ | Preferences not established until week 5 and stable until weeks 39/42 | Spencer-Booth [41] | |
| −/+ | Preferences established after week 6 and stable in weeks 11/26 | Hinde et al. [42] | |
| = | Preferences reported (no timing given) | Lindburg [39] | |
| ~ | Not tested in this species by any author(s) | ||
| M. m. tcheliensis | + | Preferences observed in weeks 1–2; no change through weeks 11–12 | Guo, et al. [49] † |
| ~ | Not tested in this species by any author(s) | ||
| M. nemestrina | = | Preferences observed after 1 month of age (cross-sectional) | Erwin, et al. [50] |
| ~ | Not tested in this species by any author(s) | ||
| M. sylvanus | = | Preferences observed on days 20–30 (single timepoint) | Regaiolli, et al. [51] † |
| ~ | Not tested in this species by any author(s) | ||
| P. anubis | = | Preferences observed in weeks 0–6 (earliest timing not given) | Damerose and Hopkins [53] |
| ~ | Not tested in this species by any author(s) | ||
| P. papio | −/+ | Preferences observed in weeks 0–2 for some individuals; all individuals had a significant and stable preference after week 2 through weeks 8–12 | Fagot [55] |
| ~ | Not tested in this species by any author(s) | ||
| P. entellus | −/+ | Preference by each twin for opposite nipples observed at weeks 1–12 (earliest timing not given); preferences strengthened weeks 13–48 | Winkler, et al. [56] |
| ~ | Not tested in this species by any author(s) | ||
| P. obscurus | −/+ | Preference established after day 80; stable through day 360 (case study) | Horwich [57] |
| ~ | Not tested in this species by any author(s) | ||
| R. roxellana | = | Preferences observed in the first week of life (single timepoint) | Zhao, et al. [58] † |
| ~ | Not tested in this species by any author(s) | ||
| P. paniscus | + | Consistent preference in longitudinal design (earliest timing not given) | Hopkins and De Lathouwers [59] † |
| ~ | Not tested in this species by any author(s) | ||
| P. troglodytes | −/+ | Mix of consistent/inconsistent preferences at 2–3 timepoints (age varied) | Nishida [61] |
| + | Consistent preference in longitudinal design (earliest timing not given) | Hopkins and De Lathouwers [59] † | |
| = | Preferences in individuals < 6 months but not > 1 year (cross-sectional) | Dienske, et al. [60] † | |
| ~ | Not tested in this species by any author(s) | ||
| Species | Developmental Outcomes | Source | |
|---|---|---|---|
| C. j. jacchus | = | Preferences observed 4–10 months (cross-sectional) | Matoba, et al. [27] * |
| − | Preferences increased in strength between 1–2 and 10–12 months | Hook and Rogers [26] | |
| −/+ | No preference at 0–2 months; stable preferences by 10–12 months | Rogers and Kaplan [28] | |
| ~ | Nipple preference did not predict hand preference at any age | Rogers and Kaplan [28] | |
| C. pencillata | − | Preferences fluctuated by task and time | Guerra, et al. [29] |
| ~ | Not tested in this species by any author(s) | ||
| C. apella | + | Positive correlation between preferences at 6 and 12 months | Westergaard, et al. [30] |
| + | Positive correlation between preferences at 23–24 and 47–48 weeks | Westergaard, et al. [31] | |
| ~ | Negative correlation between head orientation at 1–2 weeks and hand preference at 23–24 weeks, but no link to hand preference at 47–48 weeks | Westergaard, et al. [31] | |
| ~ | Stress cortisol at 6 months predicted hand preference at 12 months | Westergaard, et al. [30] | |
| M. mulatta | = | No effect of preference on reach-to-grasp kinematics at 4.5 months | Nelson, et al. [43] |
| = | No effect of age on direction or strength of preferences (cross-sectional) | Westergaard, et al. [45] | |
| − | No links between preferences measured at 1 month and 6–9 months | Nelson, et al. [48] | |
| ~ | Head orientation did not predict hand preference at 1 or 6–9 months | Nelson, et al. [48] | |
| ~ | Stress cortisol at 1 and 3 months predicted hand preference at 4–11 months | Westergaard, et al. [44] | |
| M. sylvanus | = | No hand preference at 20–30 days of life (single timepoint) | Regaiolli, et al. [51] † |
| ~ | Not tested in this species by any author(s) | ||
| P. anubis | = | Preferences observed as early as 0–4 months | Boulinguez-Ambroise, et al. [52] * |
| ~ | Hand preference in right-cradled infants changed significantly over time, whereas left- or ambiguously cradled infants’ preferences were stable | Boulinguez-Ambroise, et al. [52] * | |
| P. c. anubis | = | Preferences observed that varied by task at 6–7 months (single timepoint) | Westergaard [54] |
| ~ | Not tested in this species by any author(s) | ||
| P. papio | - | Preferences first observed at 0–2 weeks or 2–4 weeks but fluctuated | Fagot [55] † |
| ~ | Not tested in this species by any author(s) | ||
| P. troglodytes | = | No preference in spontaneous observations (case study; timing not given) | Brésard and Bresson [66] |
| = | Asymmetry in grasping strength at 14 days (mean age; single timepoint) | Fagot and Bard [67] | |
| −/+ | Preferences observed at 15 weeks; shifted through 27 months (case study) | Chorazyna [69] | |
| ~ | Neonatal hand use did not predict juvenile hand preference at 2–4 years | Hopkins and Parr [70] | |
| ~ | State regulation at 2 days predicted hand preference at 9–10 months | Bard [71] | |
| ~ | Head orientation at 3 months predicted hand preference at 9–10 months | Bard [71] | |
| ~ | Head orientation predicted hand preference at 3–5 years of age | Hopkins and Bard [68] | |
| P. pygmaeus | −/+ | Preferences observed at study onset (week 44), followed by periods of consistency and fluctuation depending on the action assessed (case study) | Cunningham, et al. [72] † |
| ~ | Not tested in this species by any author(s) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nelson, E.L.; Karimi, A. Systematic Review: The Development of Behavioral Laterality Across the First Year of Life in Nonhuman Primates. Symmetry 2023, 15, 1335. https://doi.org/10.3390/sym15071335
Nelson EL, Karimi A. Systematic Review: The Development of Behavioral Laterality Across the First Year of Life in Nonhuman Primates. Symmetry. 2023; 15(7):1335. https://doi.org/10.3390/sym15071335
Chicago/Turabian StyleNelson, Eliza L., and Atefeh Karimi. 2023. "Systematic Review: The Development of Behavioral Laterality Across the First Year of Life in Nonhuman Primates" Symmetry 15, no. 7: 1335. https://doi.org/10.3390/sym15071335
APA StyleNelson, E. L., & Karimi, A. (2023). Systematic Review: The Development of Behavioral Laterality Across the First Year of Life in Nonhuman Primates. Symmetry, 15(7), 1335. https://doi.org/10.3390/sym15071335