
Parameter Estimation in the Mathematical Model of Bacterial Colony Patterns in Symmetry Domain





Abstract
1. Introduction
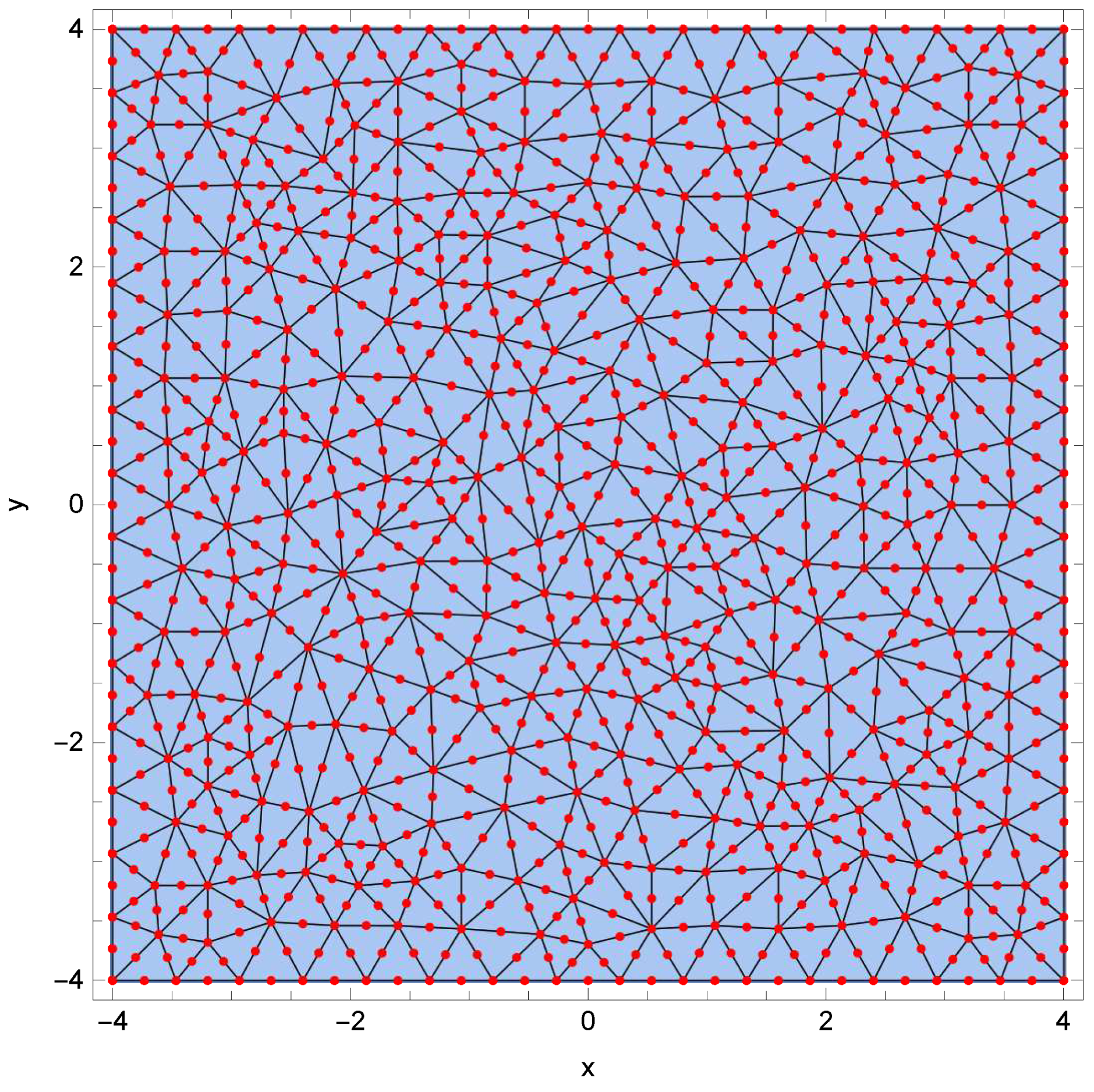
2. Bacterial Colonies Model
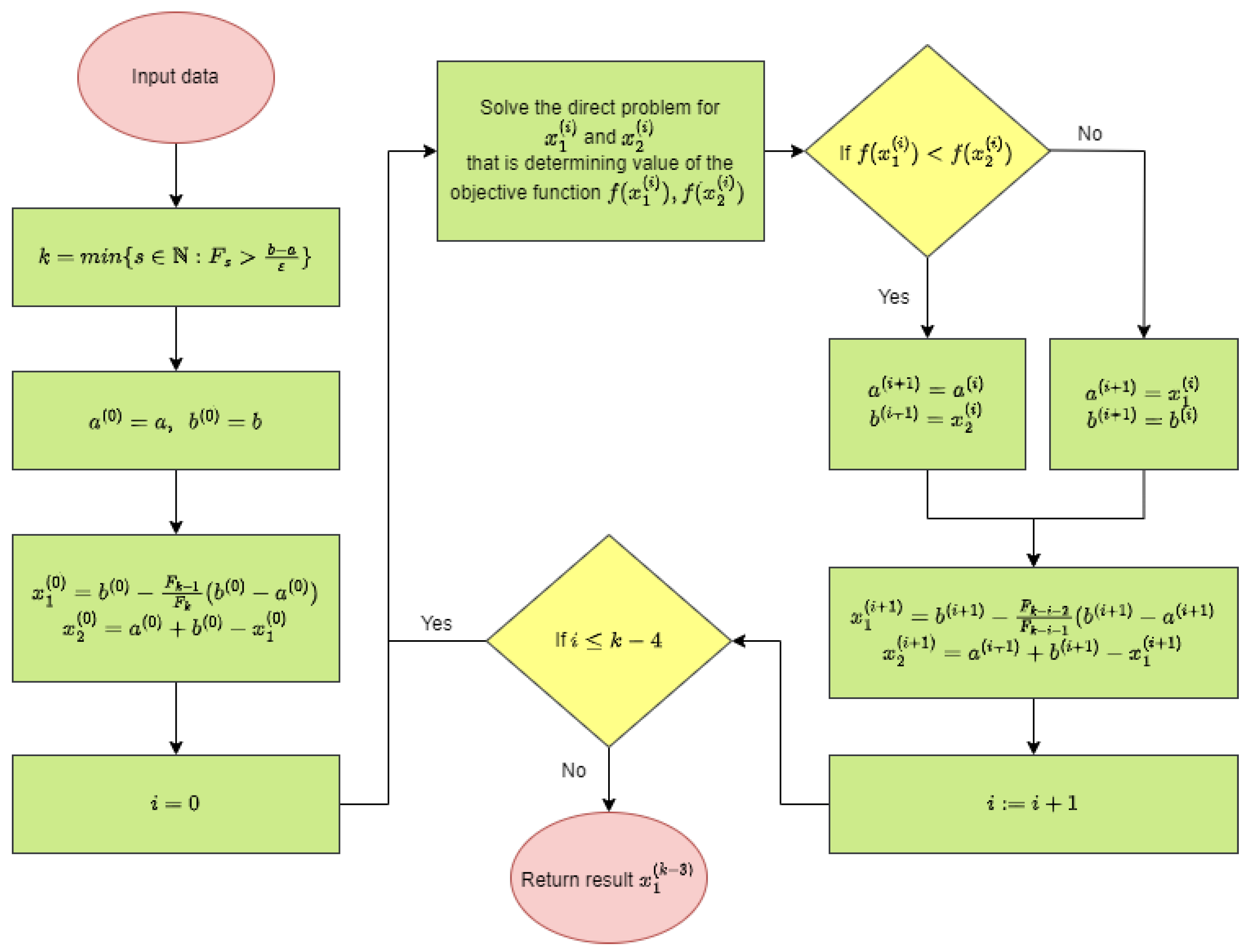
3. Inverse Problem and Its Solution
- 1.
- Input: objective function f (unimodal function),
- interval ,
- required absolute precision .
- 2.
- Determine the smallest number for which , where is the Fibonacci number.
- 3.
- Set and .
- 4.
- Determine and .
- 5.
- For :
- 5a.
- If , then and ,otherwise and .
- 5b.
- Determine
- 6.
- Result .
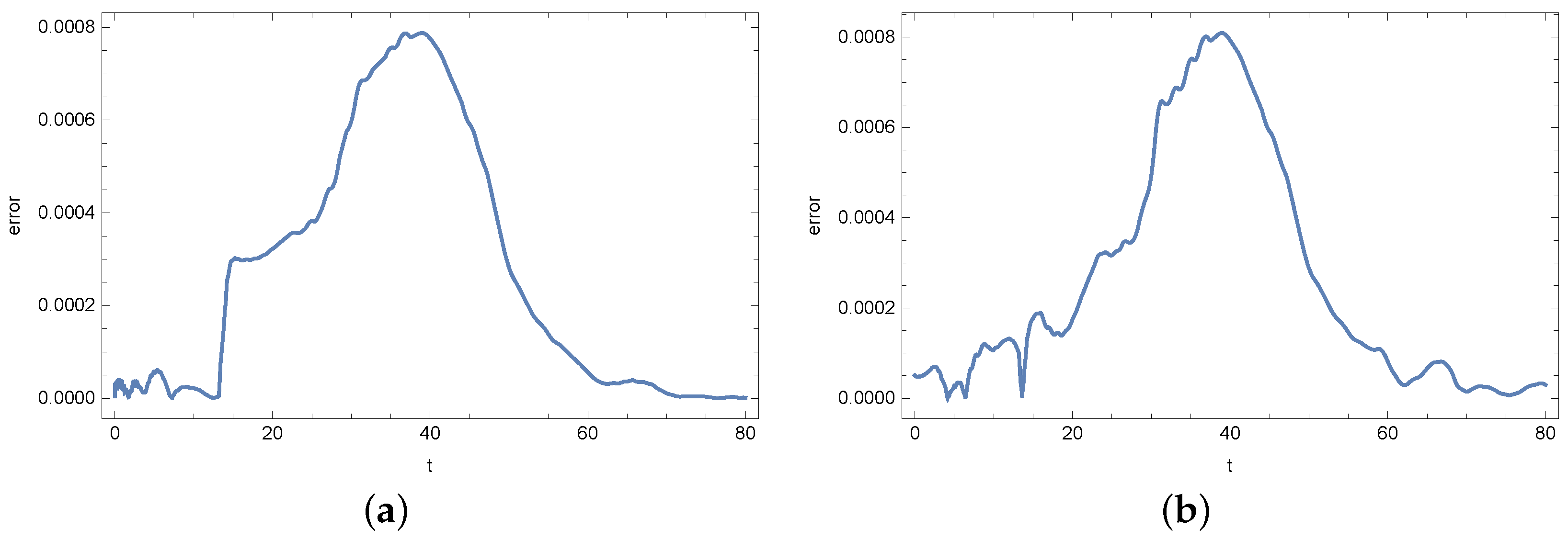
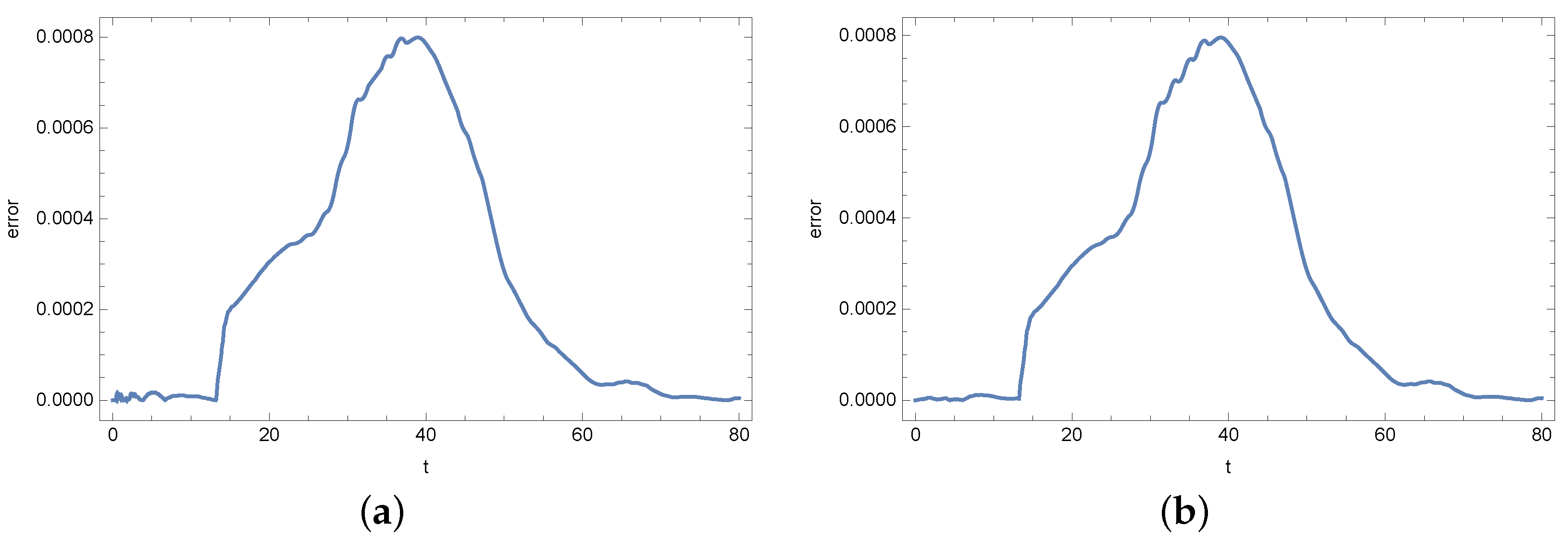
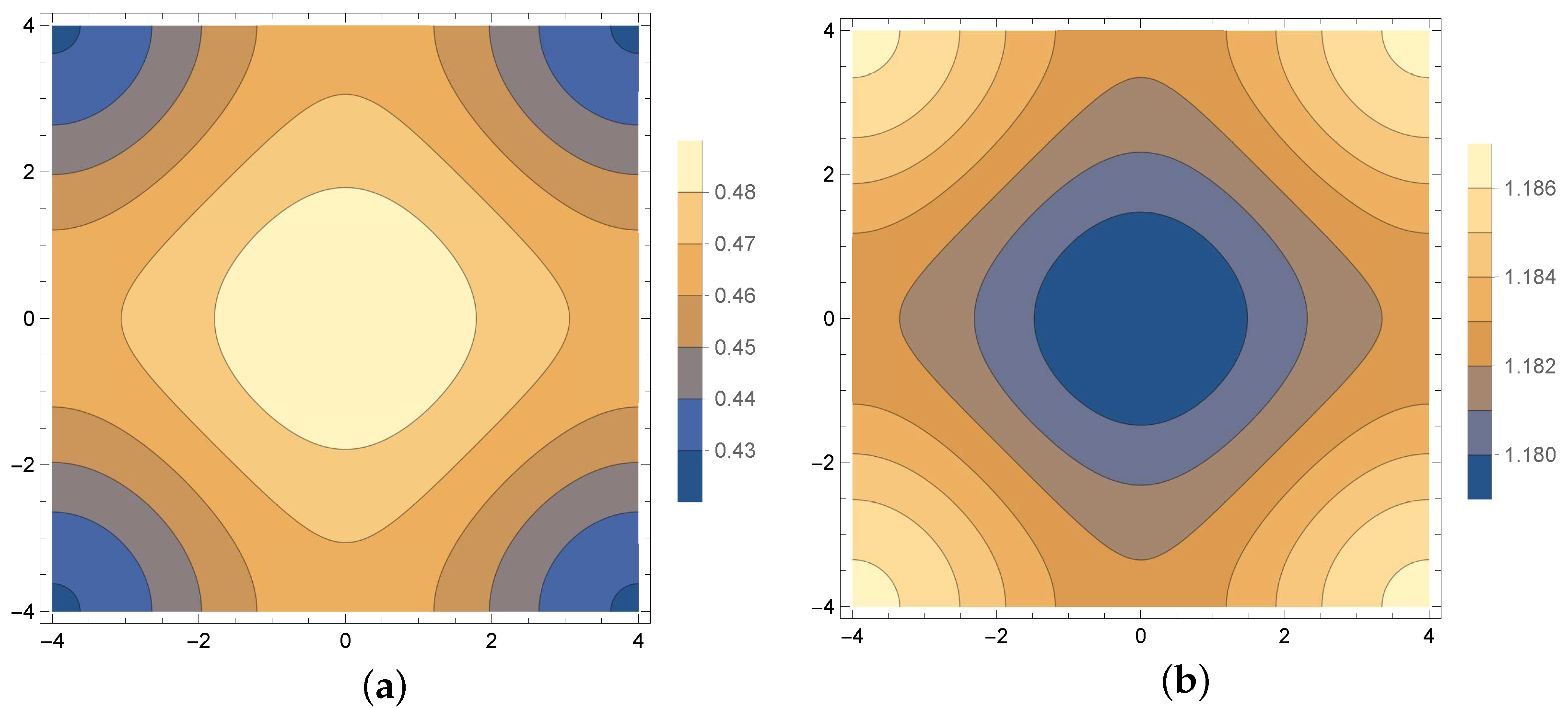
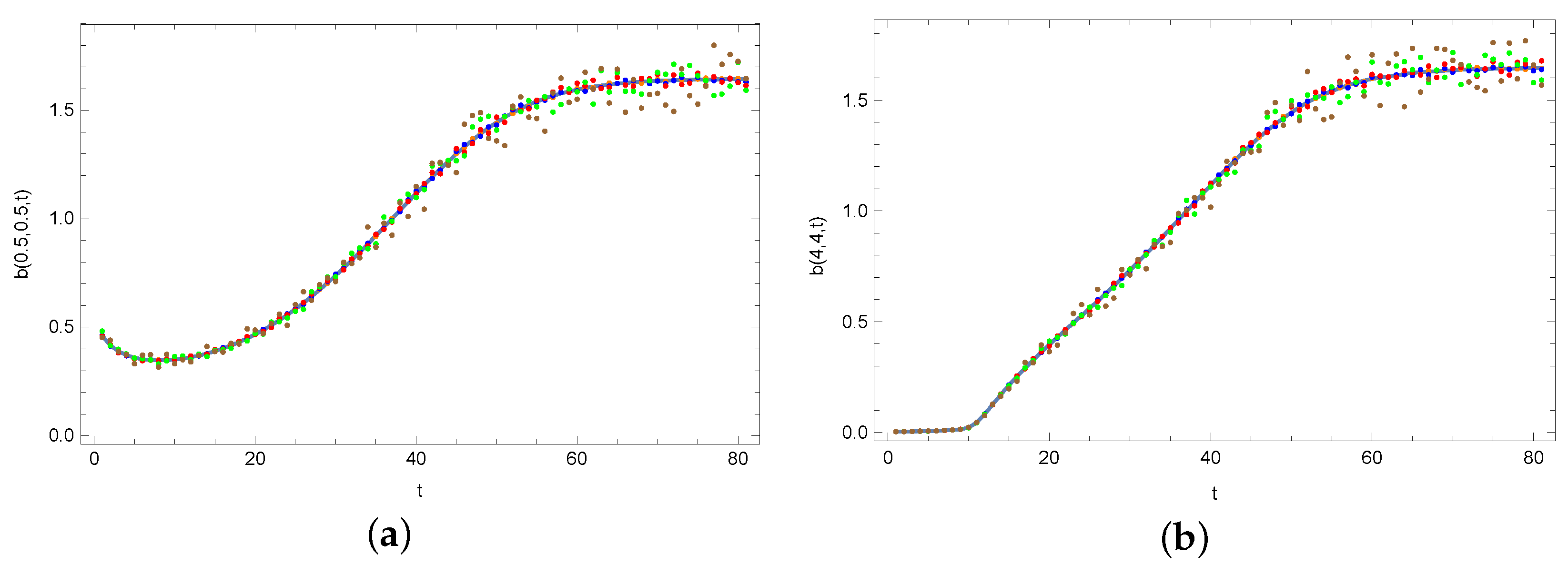
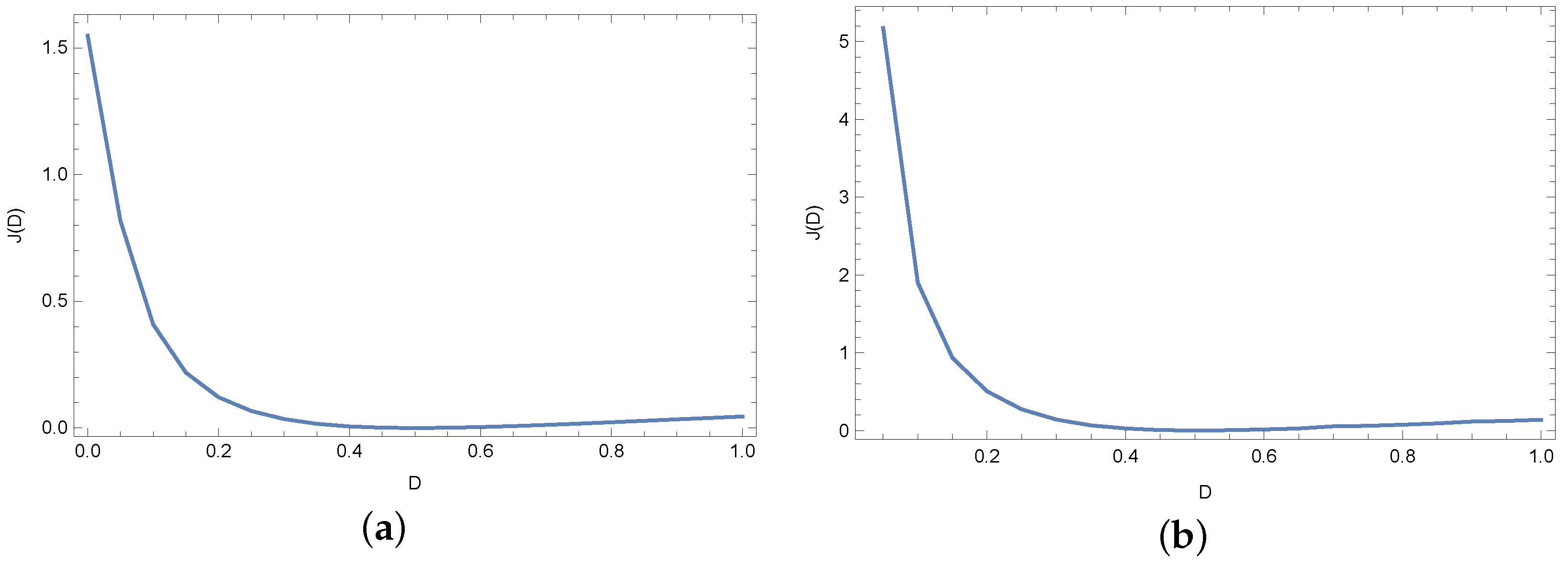
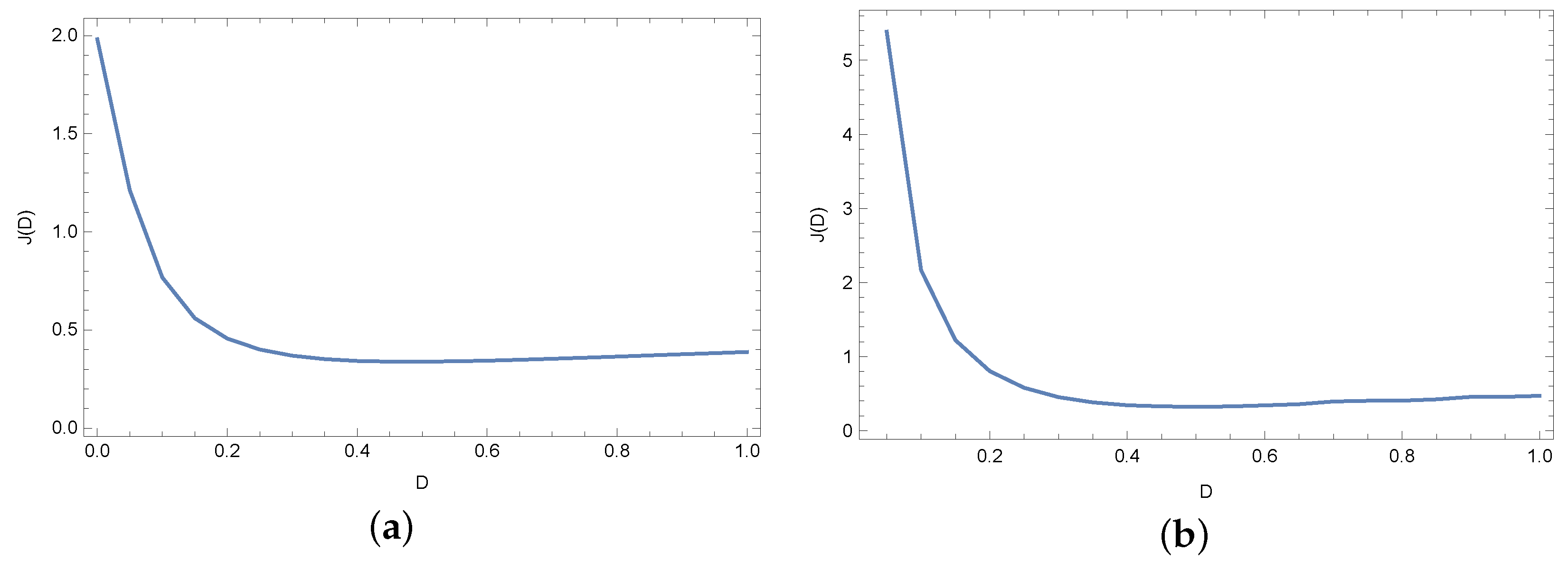
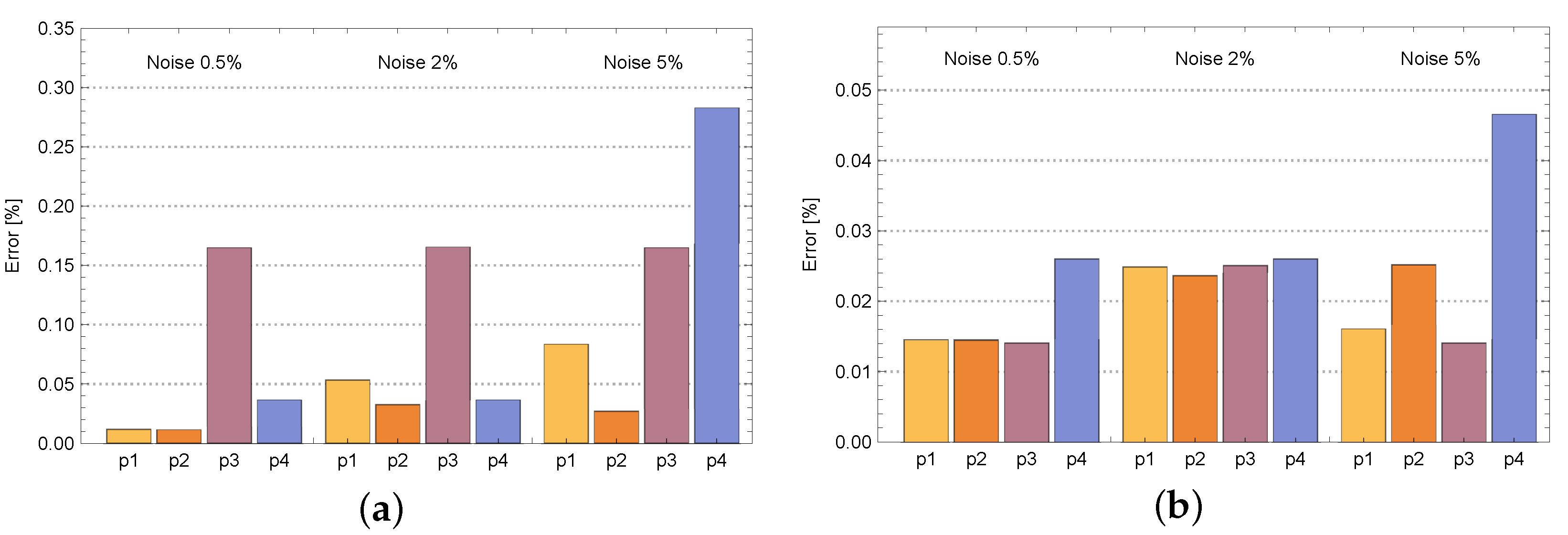
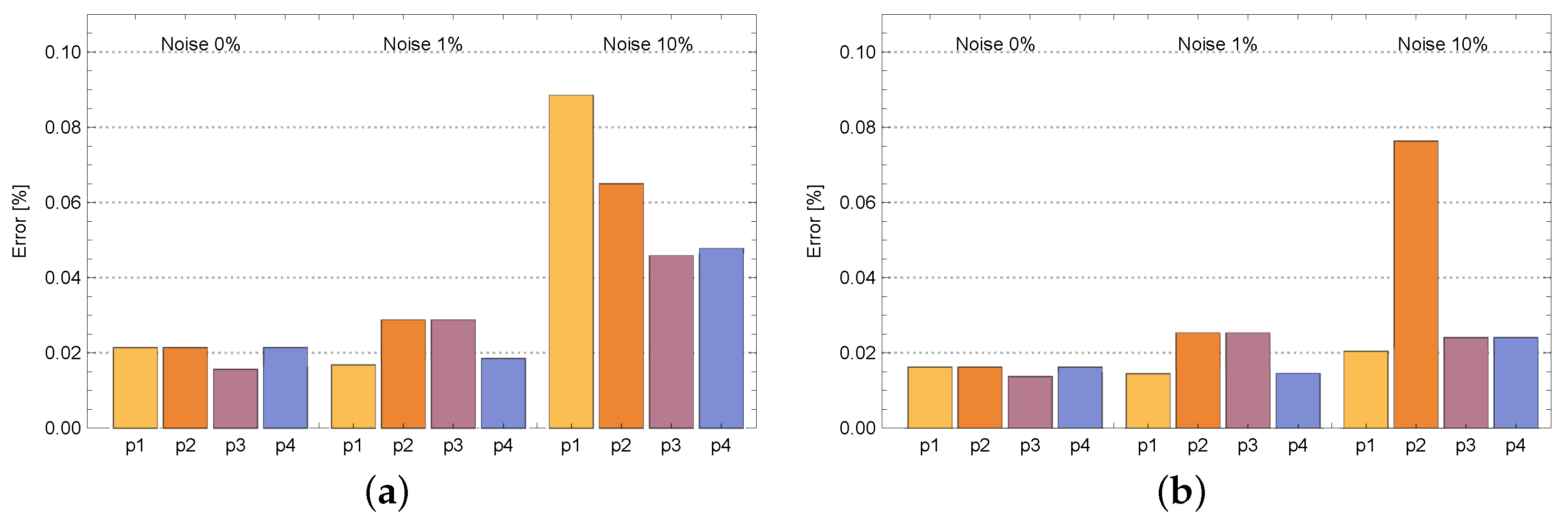
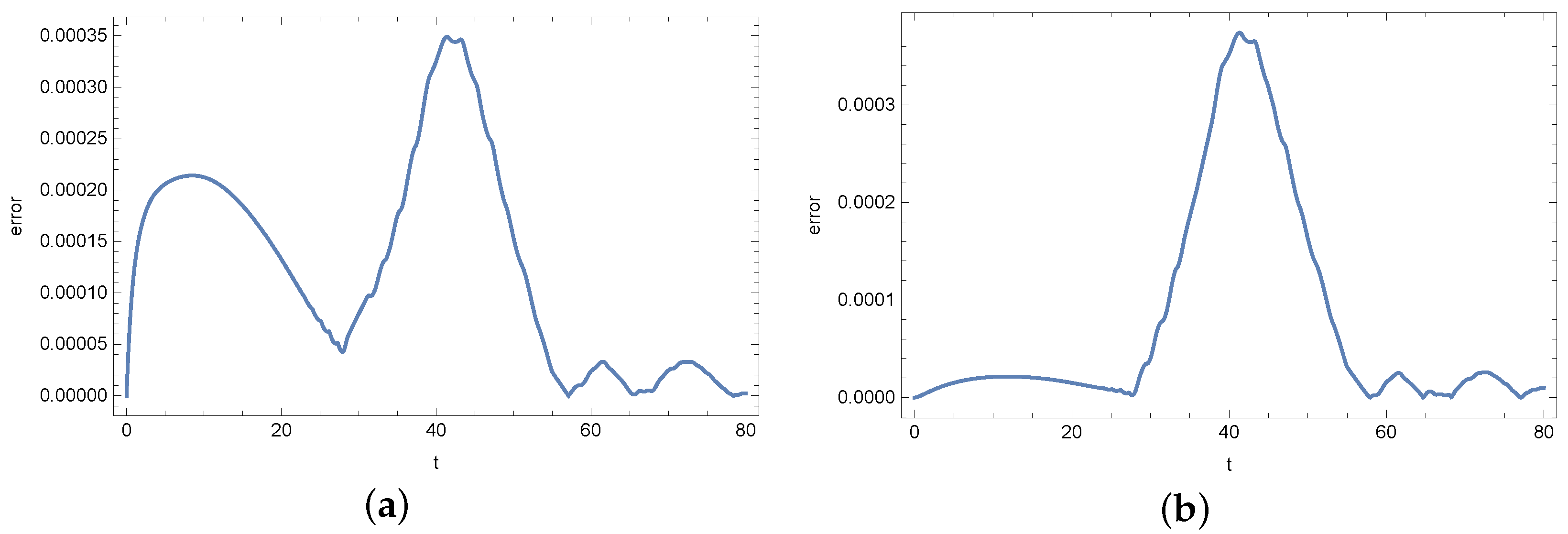
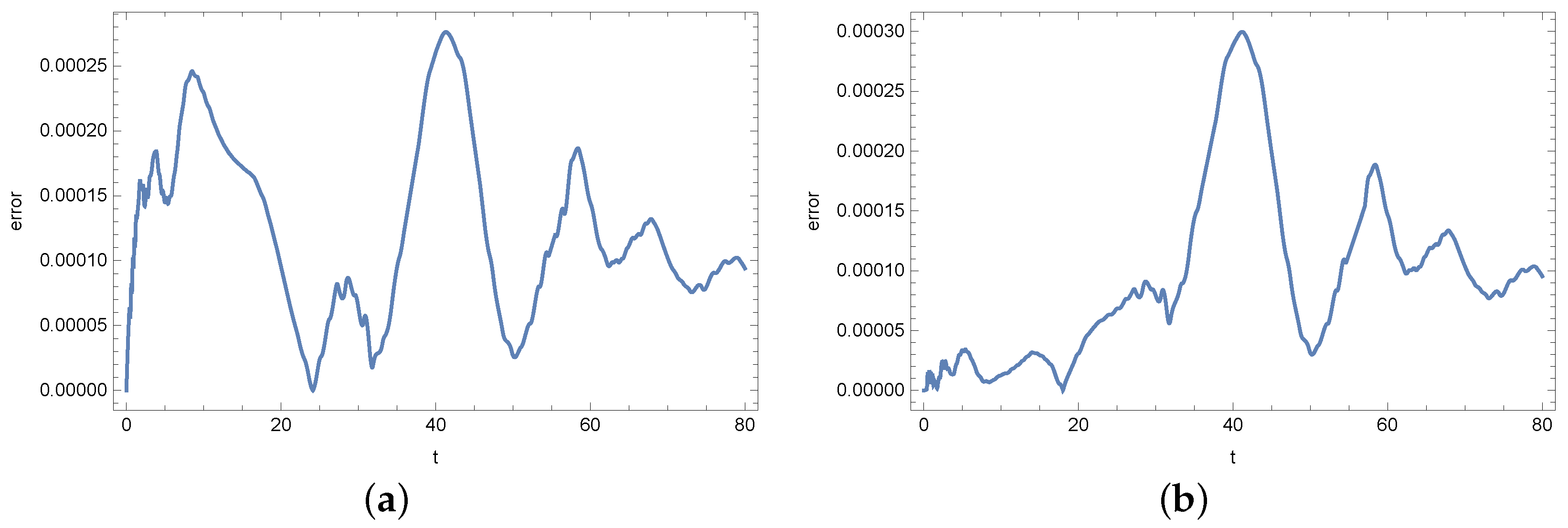
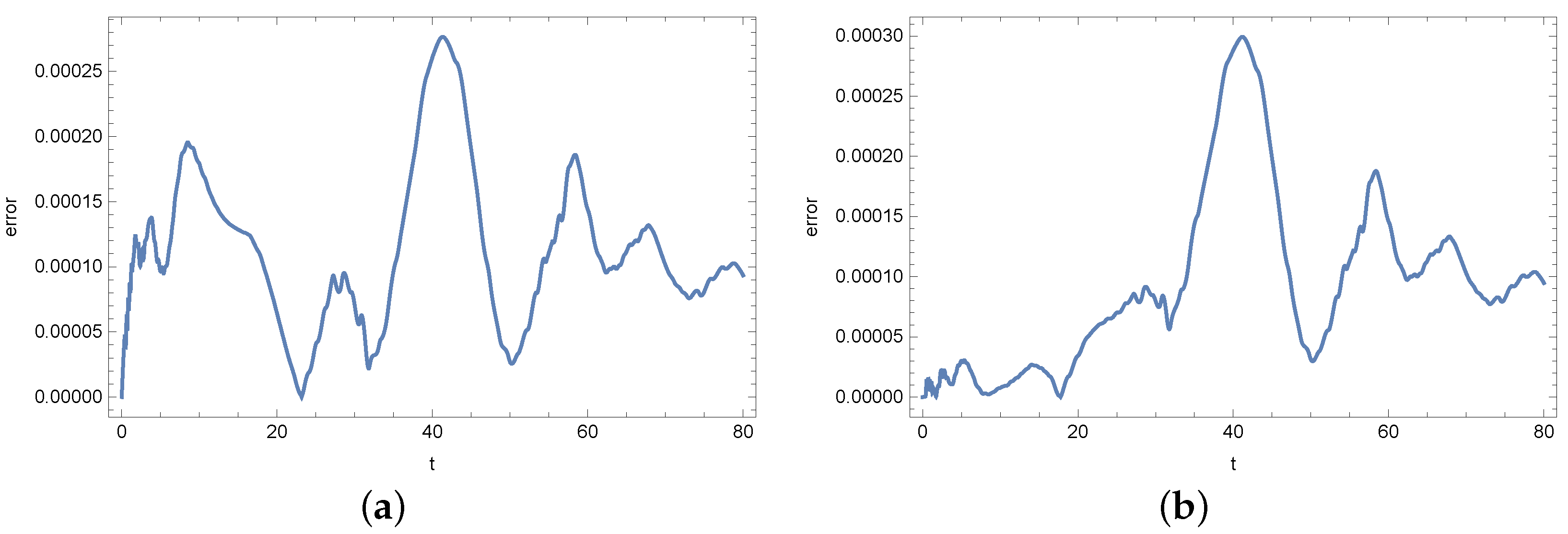
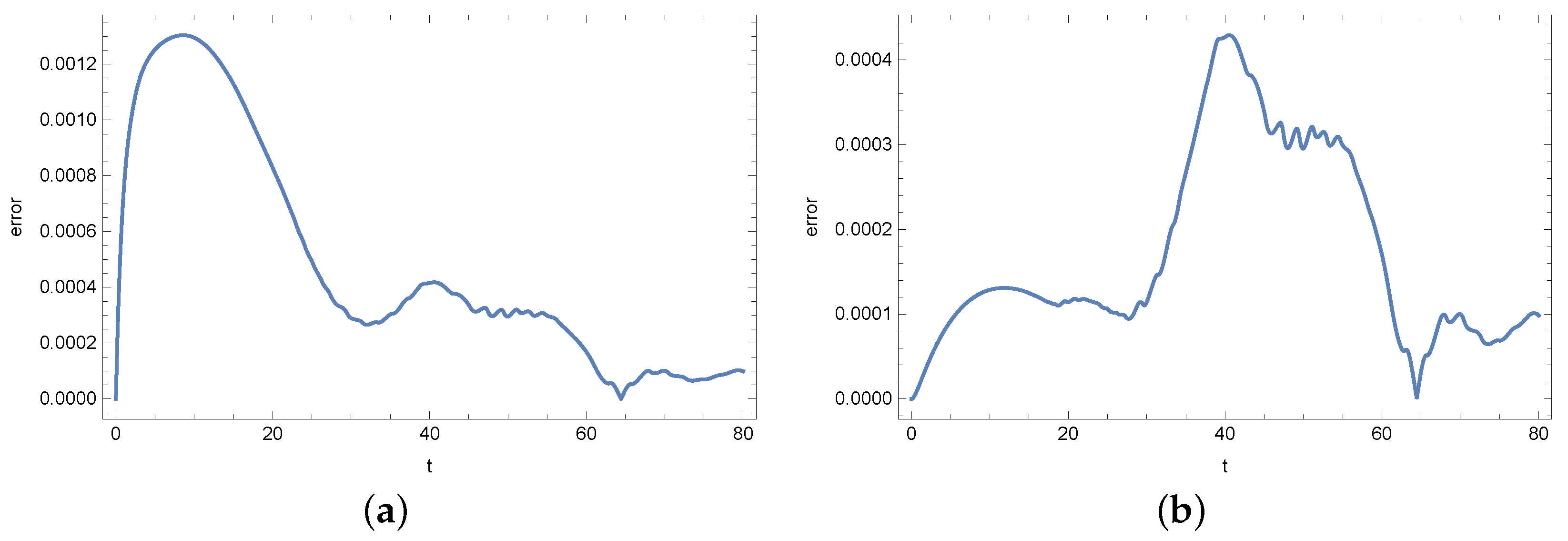
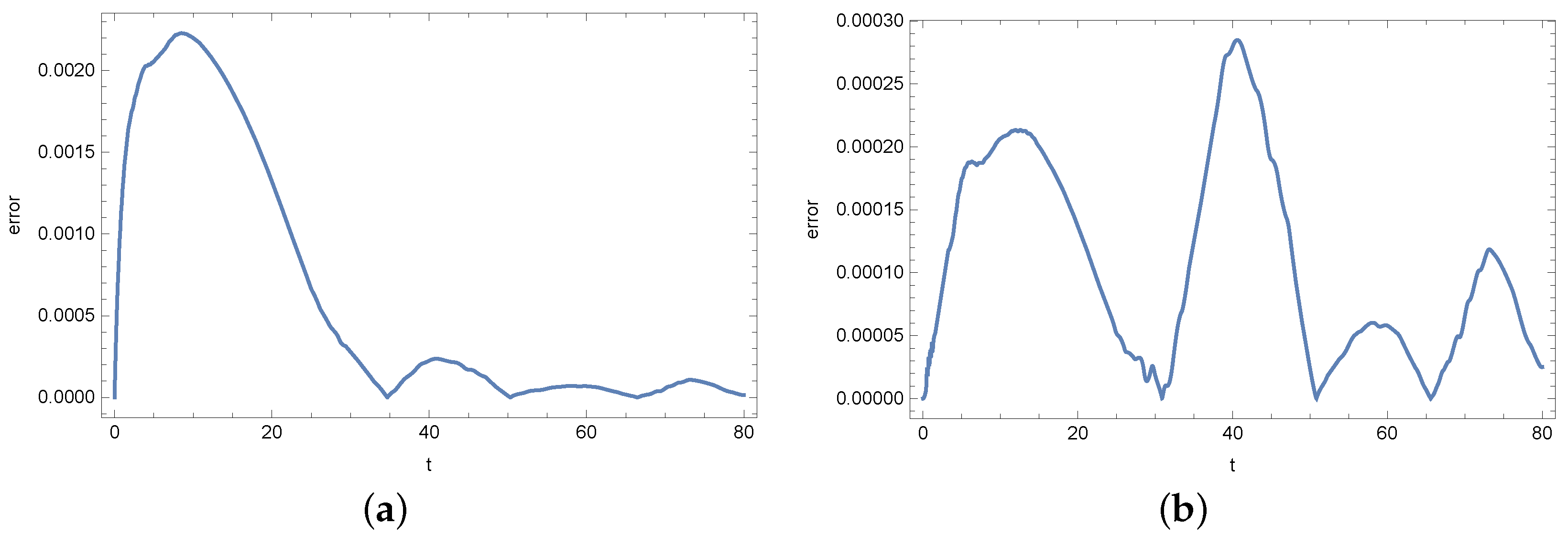
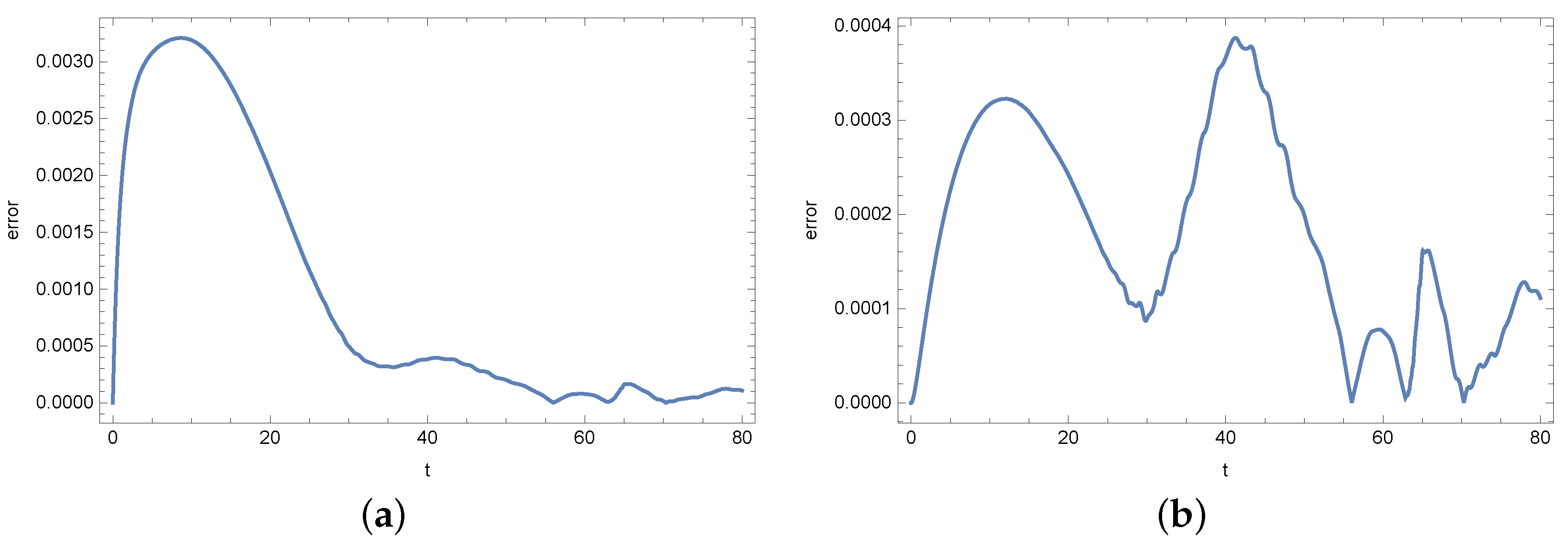
4. Results
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xiong, L.; Cao, Y.; Cooper, R.; Rappel, W.-J.; Hasty, J.; Tsimring, L. Flower-like patterns in multi-species bacterial colonies. eLIFE 2020, 9, e48885. [Google Scholar] [CrossRef]
- Rhodeland, B.; Hoeger, K.; Ursell, T. Bacterial surface motility is modulated by colony-scale flow and granular jamming. J. R. Soc. Interface 2020, 17, 20200147. [Google Scholar] [CrossRef] [PubMed]
- Matoz-Fernandez, D.; Arnaouteli, S.; Porter, M.; MacPhee, C.E.; Stanley-Wall, N.R.; Davidson, F.A. Comment on ”Rivalry in Bacillus subtilis colonies: Enemy or family?”. Soft Matter 2020, 16, 3344–3346. [Google Scholar] [CrossRef] [PubMed]
- Brzozowski, R.S.; Tomlinson, B.R.; Sacco, M.D.; Chen, J.J.; Ali, A.N.; Chen, Y.; Shaw, L.N.; Eswaraa, P.J. Interdependent YpsA- and YfhS-Mediated Cell Division and Cell Size Phenotypes in Bacillus subtilis. mSphere 2020, 5, e00655-20. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Valdes, J.A.; Zhou, L.; de Vries, M.P.; Kuipers, O.P. Impact of spatial proximity on territoriality among human skin bacteria. NPJ Biofilms Microbiomes 2020, 6, 30. [Google Scholar] [CrossRef] [PubMed]
- Sui, Z.-W.; Wang, B.; Liu, S.-Y.; Wang, J.; Fu, B.-Q.; Zhuo, T.-Y.; Wang, Y. Study on Measurement Method of Bacillus subtilis var. niger Spore. Jiliang Xuebao/Acta Metrol. Sin. 2020, 41, 1171–1176. [Google Scholar]
- Earl, C.; Arnaouteli, S.; Bamford, N.C.; Porter, M.; Sukhodub, T.; MacPhee, C.E.; Stanley-Wall, N.R. The majority of the matrix protein TapA is dispensable for Bacillus subtilis colony biofilm architecture. Mol. Microbiol. 2020. [Google Scholar] [CrossRef]
- Schwarcz, D.; Levine, H.; Ben-Jacob, E.; Ariel, G. Uniform modeling of bacterial colony patterns with varying nutrient and substrate. Phys. D 2016, 318–319, 91–99. [Google Scholar] [CrossRef]
- Miyata, S.; Sasaki, T. Asymptotic analysis of a chemotactic model of bacteria colonies. Math. Biosci. 2006, 201, 184–194. [Google Scholar] [CrossRef]
- Golding, I.; Kozlovsky, Y.; Cohen, I.; Ben-Jacob, E. Studies of bacterial branching growth using reaction-diffusion models for colonial development. Phys. A 1998, 260, 510–554. [Google Scholar] [CrossRef]
- Shimada, H.; Ikeda, T.; Wakita, J.; Itoh, H.; Kurosu, S.; Hiramatsu, F.; Nakatsuchi, M.; Yamazaki, Y.; Matsuyama, T.; Matsushita, M. Dependence of local cell density on concentric ring colony formation by bacterial species bacillus subtilis. J. Phys. Soc. Jpn. 2003, 73, 1082–1089. [Google Scholar] [CrossRef]
- Brociek, R.; Słota, D.; Król, M.; Matula, G.; Kwaśny, W. Comparison of mathematical models with fractional derivative for the heat conduction inverse problem based on the measurements of temperature in porous aluminum. Int. J. Heat Mass Transf. 2019, 143, 118440. [Google Scholar] [CrossRef]
- Brociek, R.; Słota, D.; Król, M.; Matula, G.; Kwaśny, W. Modeling of heat distribution in porous aluminum using fractional differential equation. Fractal Fract. 2017, 1, 17. [Google Scholar] [CrossRef]
- Shang, Z.; Liao, Z.; Sarasua, J.A.; Billingham, J.; Axinte, D. On modelling of laser assisted machining: Forward and inverse problems for heat placement control. Int. J. Mach. Tools Manuf. 2019, 138, 36–50. [Google Scholar] [CrossRef]
- Kefai, A.; Yildiz, M. Modeling of sensor placement strategy for shape sensing and structural health monitoring of a wing-shaped sandwich panel using inverse finite element method. Sensors 2017, 17, 2775. [Google Scholar]
- Liang, D.; Cheng, J.; Ke, Z.; Ying, L. Deep magnetic resonance image reconstruction: Inverse problems meet neural networks. IEEE Signal Process. Mag. 2020, 37, 141–151. [Google Scholar] [CrossRef]
- Brociek, R.; Hetmaniok, E.; Słota, D. Reconstruction of aerothermal heating for the thermal protection system of a reusable launch vehicle. Appl. Therm. Eng. 2023, 219, 119405. [Google Scholar] [CrossRef]
- Brociek, R.; Hetmaniok, E.; Napoli, C.; Capizzi, G.; Słota, D. Estimation of aerothermal heating for a thermal protection system with temperature dependent material properties. Int. J. Therm. Sci. 2023, 188, 108229. [Google Scholar] [CrossRef]
- Smyl, D.; Liu, D. Less is often more: Applied inverse problems using hp-forward models. J. Comput. Phys. 2019, 399, 108949. [Google Scholar] [CrossRef]
- Kaipio, J.; Somersalo, E. Statistical and Computational Inverse Problems; Springer: New York, NY, USA, 2005. [Google Scholar]
- Kaipio, J.; Somersalo, E. Statistical inverse problems: Discretization, model reduction and inverse crimes. J. Comput. Appl. Math. 2007, 198, 493–504. [Google Scholar] [CrossRef]
- Clermont, G.; Zenker, S. The inverse problem in mathematical biology. Math. Biosci. 2015, 260, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Abdulla, U.G.; Poteau, R. Identification of parameters in systems biology. Math. Biosci. 2018, 305, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Capasso, V.; Kunze, H.E.; La Torre, D.; Vrscay, E.R. Solving inverse problems for biological models using the collage method for differential equations. J. Math. Biol. 2013, 67, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Kabanikhin, S.I.; Krivorotko, O.I. Optimization methods for solving inverse immunology and epidemiology problems. Comput. Math. Math. Phys. 2020, 60, 580–589. [Google Scholar] [CrossRef]
- Doumic, M.; Maia, P.; Zubelli, J.P. On the calibration of a size-structured population model from experimental data. Acta Biotheor. 2010, 58, 405–413. [Google Scholar] [CrossRef]
- Barnsley, M.F. Fractals Everywhere; AP Professional: Boston, MA, USA, 2012. [Google Scholar]
- Bao, W.; Yang, B.; Chen, B. 2-hydr_Ensemble: Lysine 2-hydroxyisobutyrylation identification with ensemble method. Chemom. Intell. Lab. Syst. 2021, 215, 104351. [Google Scholar] [CrossRef]
- Yang, B.; Bao, W.; Wang, J. Active disease-related compound identification based on capsule network. Briefings Bioinform. 2022, 23, bbab462. [Google Scholar] [CrossRef]
- Bao, W.; Cui, Q.; Chen, B.; Yang, B. Phage_UniR_LGBM: Phage Virion Proteins Classification with UniRep Features and LightGBM Model. Comput. Math. Methods Med. 2022, 2022, 9470683. [Google Scholar] [CrossRef]
- Rida, S.Z.; El-Sayed, A.M.A.; Arafa, A.A.M. Effect of bacterial memory dependent growth by using fractional derivatives reaction-diffusion chemotactic model. J. Stat. Phys. 2010, 140, 797–811. [Google Scholar] [CrossRef]
- Kawasaki, K.; Mochizuki, A.; Matsushita, M.; Umeda, T.; Shigesada, N. Modeling spatio-temporal patterns created by Bacillus subtilis. J. Teor. Biol. 1997, 188, 177–185. [Google Scholar] [CrossRef]
- Gill, P.E.; Murray, W.; Wright, M.H. Practical Optimization; Academic Press: London, UK, 1981. [Google Scholar]
- Mishra, S.K.; Ram, B. Introduction to Unconstrained Optimization with R; Springer Nature: Singapore, 2019. [Google Scholar]
- Wolfram, S. An Elementary Introduction to the Wolfram Language; Wolfram Media: Champaign, IL, USA, 2017. [Google Scholar]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
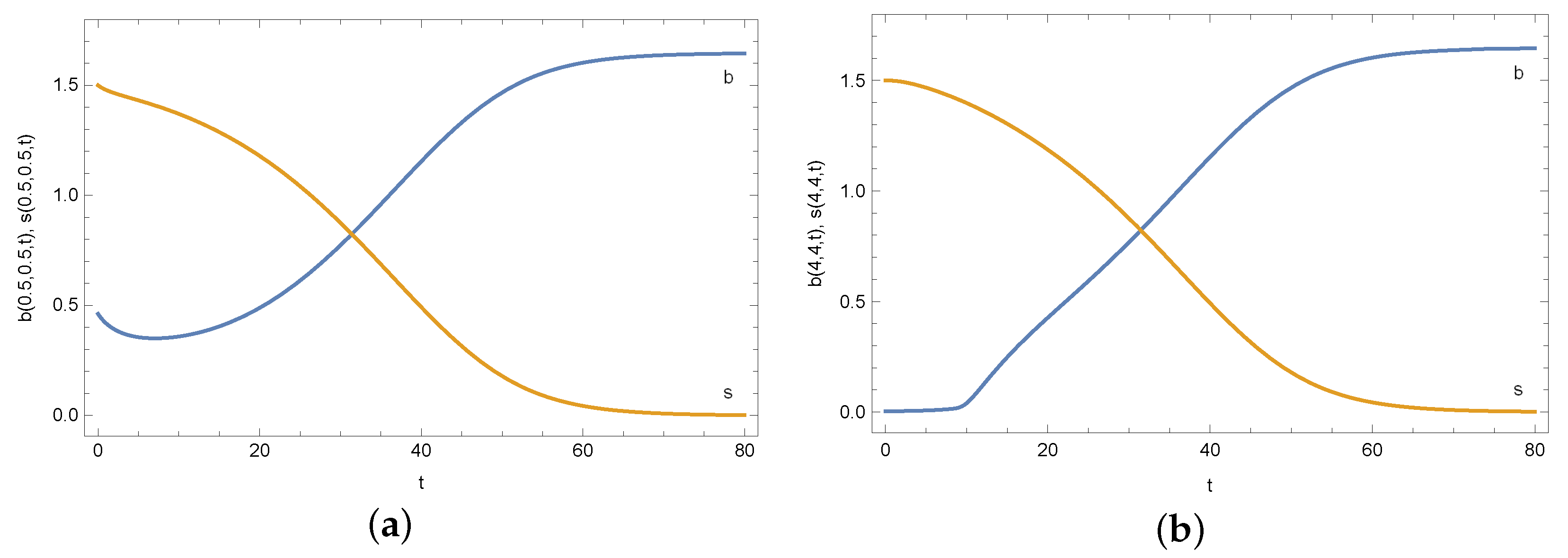{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| [%] | ||||
|---|---|---|---|---|
| 0% | ||||
| % | ||||
| 1% | ||||
| 2% | ||||
| 5% | ||||
| 10% |
| [%] | ||||
|---|---|---|---|---|
| 0% | ||||
| % | ||||
| 1% | ||||
| 2% | ||||
| 5% | ||||
| 10% |
| [%] | [%] | |||
|---|---|---|---|---|
| 0% | ||||
| % | ||||
| 1% | ||||
| 2% | ||||
| 5% | ||||
| 10% |
| [%] | [%] | |||
|---|---|---|---|---|
| 0% | ||||
| % | ||||
| 1% | ||||
| 2% | ||||
| 5% | ||||
| 10% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brociek, R.; Wajda, A.; Capizzi, G.; Słota, D. Parameter Estimation in the Mathematical Model of Bacterial Colony Patterns in Symmetry Domain. Symmetry 2023, 15, 782. https://doi.org/10.3390/sym15040782
Brociek R, Wajda A, Capizzi G, Słota D. Parameter Estimation in the Mathematical Model of Bacterial Colony Patterns in Symmetry Domain. Symmetry. 2023; 15(4):782. https://doi.org/10.3390/sym15040782
Chicago/Turabian StyleBrociek, Rafał, Agata Wajda, Giacomo Capizzi, and Damian Słota. 2023. "Parameter Estimation in the Mathematical Model of Bacterial Colony Patterns in Symmetry Domain" Symmetry 15, no. 4: 782. https://doi.org/10.3390/sym15040782
APA StyleBrociek, R., Wajda, A., Capizzi, G., & Słota, D. (2023). Parameter Estimation in the Mathematical Model of Bacterial Colony Patterns in Symmetry Domain. Symmetry, 15(4), 782. https://doi.org/10.3390/sym15040782