Zebrafish Melanophores Suggest Novel Functions of Cell Chirality in Tissue Formation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Main
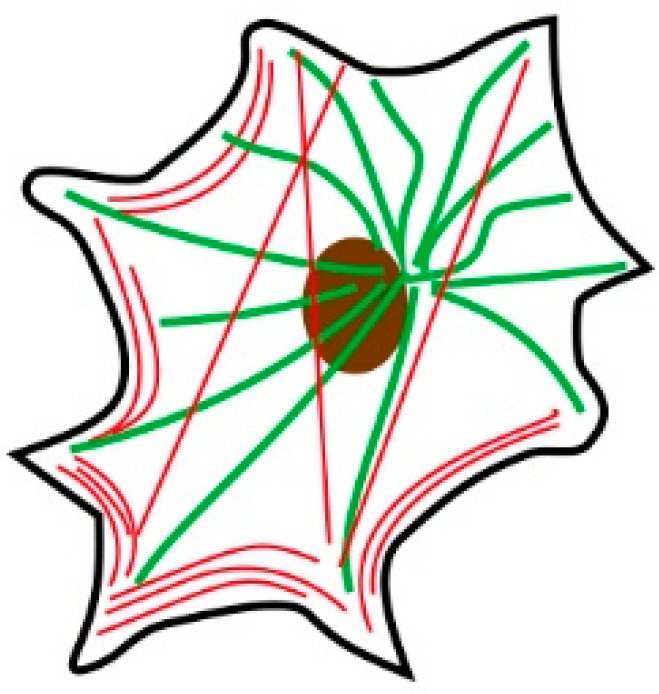
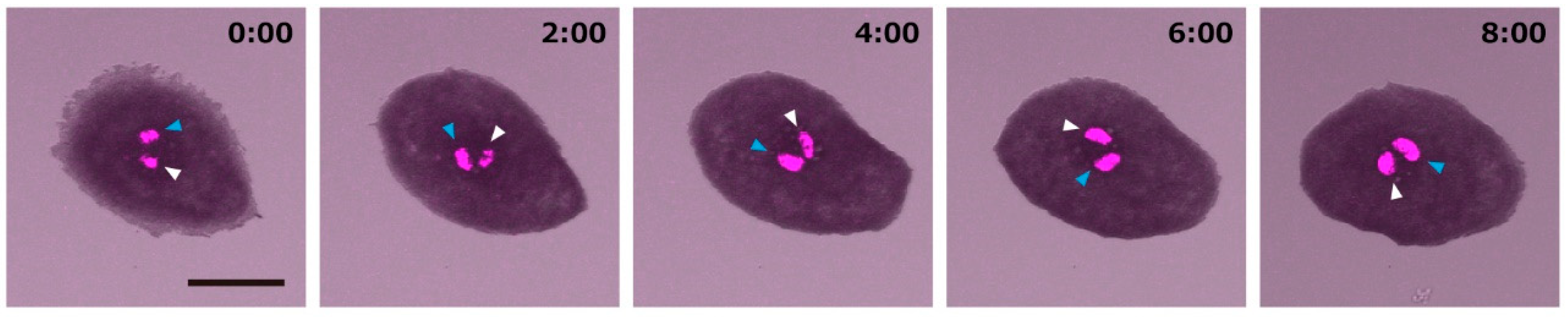
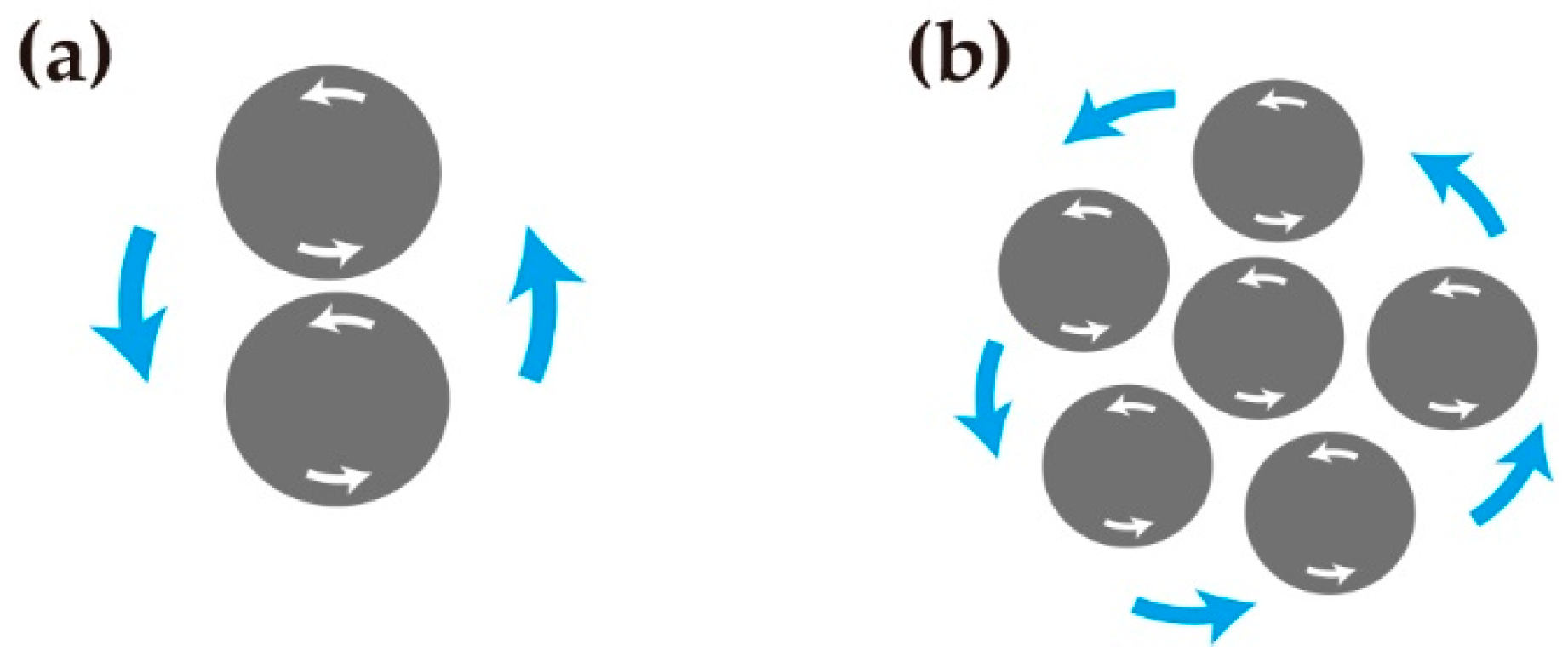
2.1. Zebrafish Melanophores Have Inherent Cellular Chirality
2.2. Rotation of Melanophores Regulated by Actin Fibers and Microtubules
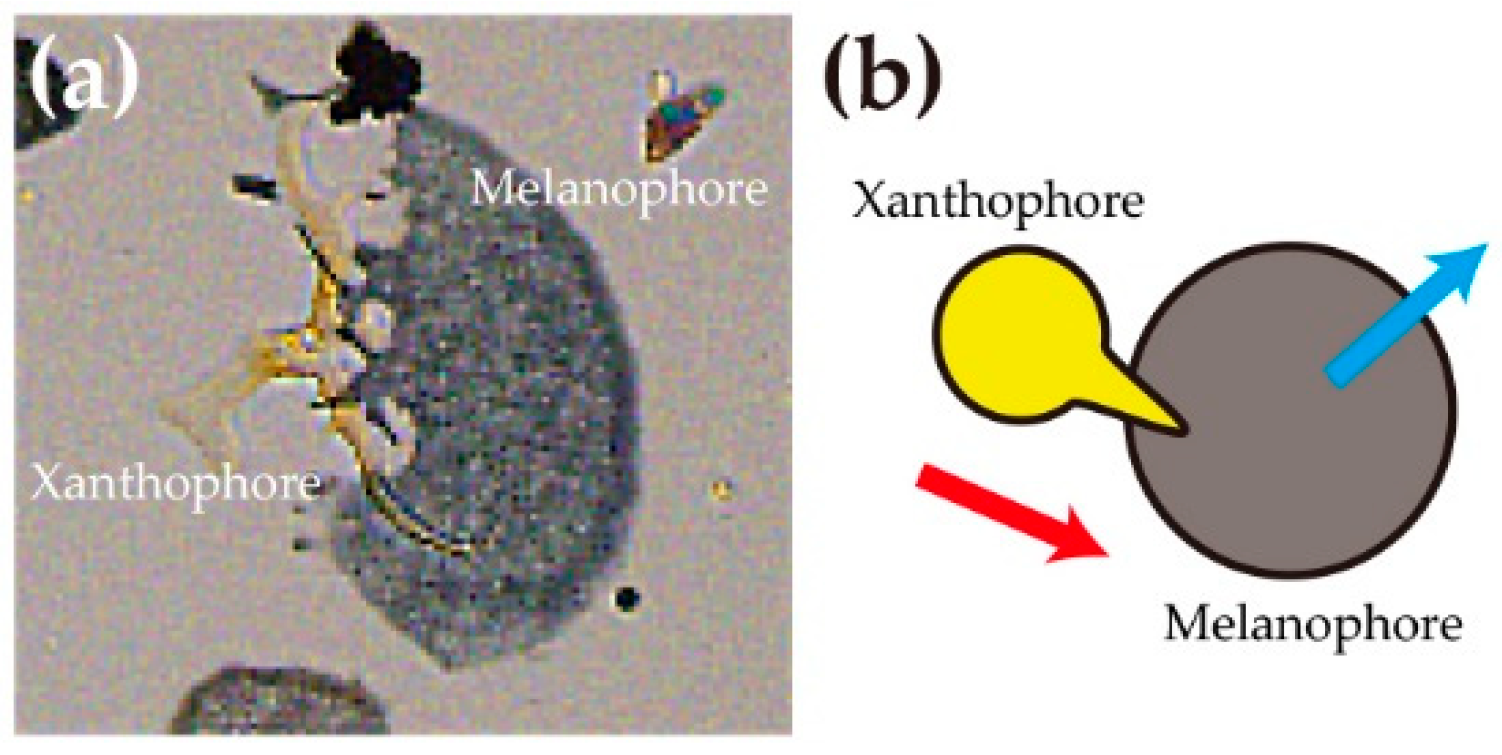
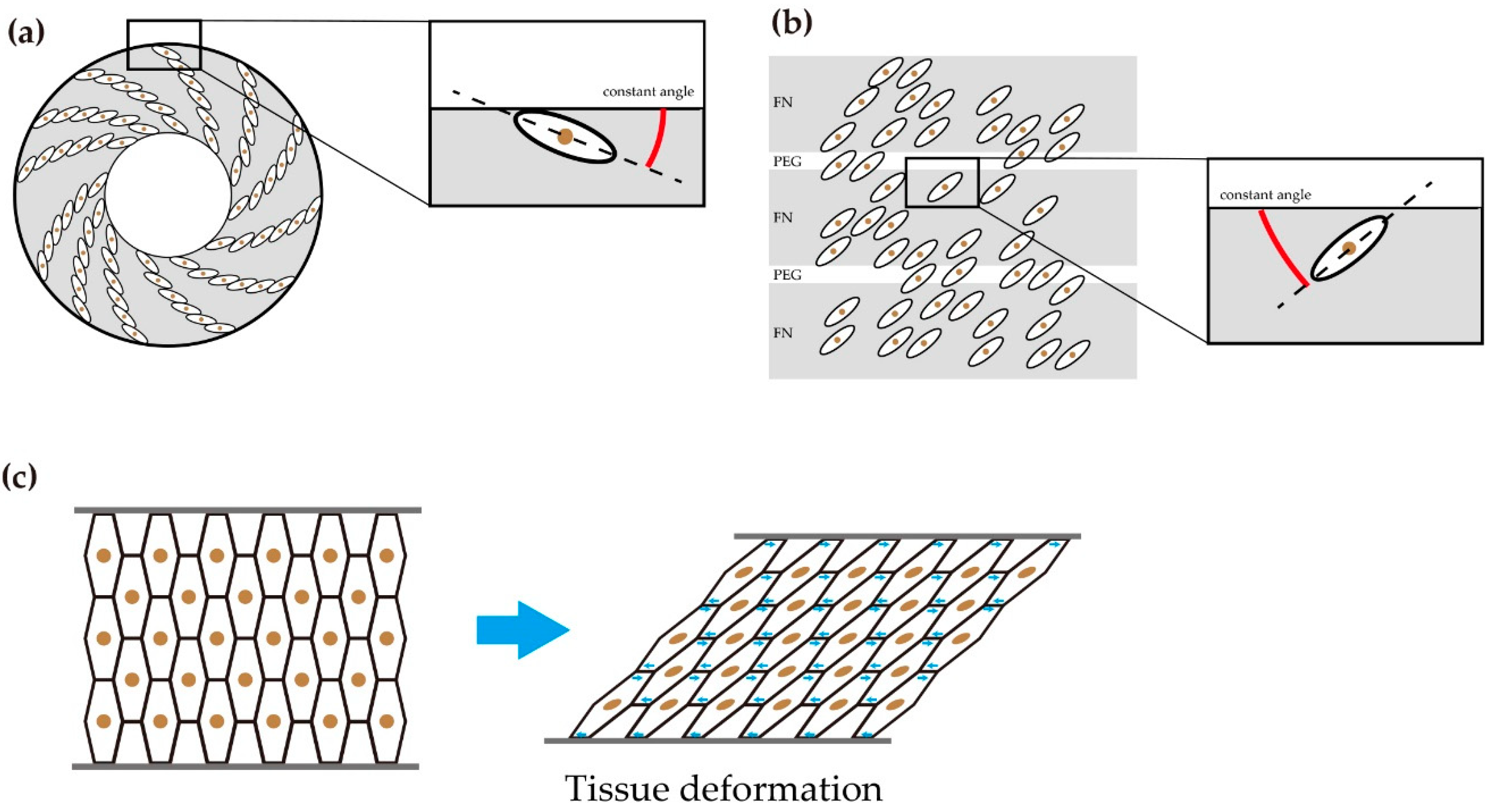
2.3. The Function of Chirality of Melanophore during Pigment Pattern Formation
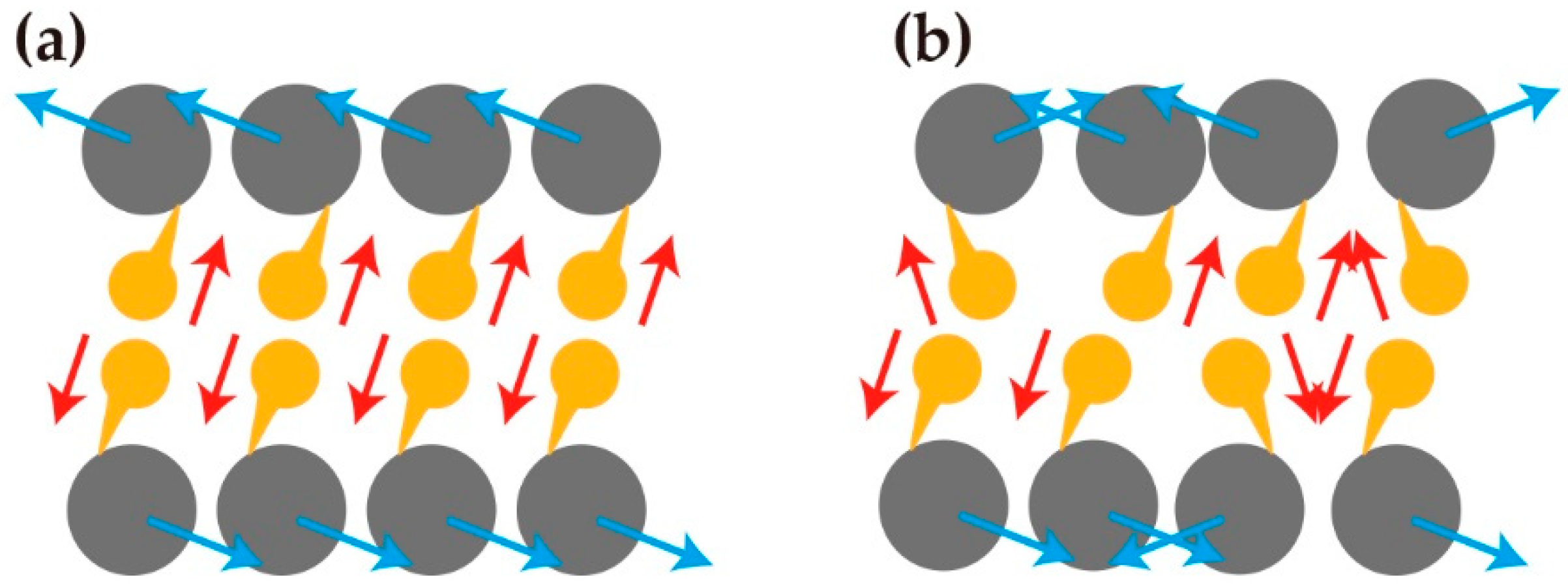
2.4. Chirality in a Group of Cells Induces the Deformation of Tissue
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hagmann, J. Pattern formation and handedness in the cytoskeleton of human platelets. Proc. Natl. Acad. Sci. USA 1993, 90, 3280–3283. [Google Scholar] [CrossRef] [PubMed]
- Tee, Y.H.; Shemesh, T.; Thiagarajan, V.; Hariadi, R.F.; Anderson, K.L.; Page, C.; Volkmann, N.; Hanein, D.; Sivaramakrishnan, S.; Kozlov, M.M.; et al. Cellular chirality arising from the self-organization of the actin cytoskeleton. Nat. Cell Biol. 2015, 17, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.Q.; Ronaldson, K.; Park, M.; Taylor, G.; Zhang, Y.; Gimble, J.M.; Vunjak-Novakovic, G. Micropatterned mammalian cells exhibit phenotype-specific left-right asymmetry. Proc. Natl. Acad. Sci. USA 2011, 108, 12295–12300. [Google Scholar] [CrossRef] [PubMed]
- Heacock, A.M.; Agranoff, B.W. Clockwise growth of neurites from retinal explants. Science 1977, 198, 64–66. [Google Scholar] [CrossRef] [PubMed]
- Tamada, A.; Kawase, S.; Murakami, F.; Kamiguchi, H. Autonomous right-screw rotation of growth cone filopodia drives neurite turning. J. Cell Biol. 2010, 188, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Van Keymeulen, A.; Wakida, N.M.; Carlton, P.; Berns, M.W.; Bourne, H.R. Polarity reveals intrinsic cell chirality. Proc. Natl. Acad. Sci. USA 2007, 104, 9296–9300. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, S.; Tanaka, Y.; Okada, Y.; Takeda, S.; Harada, A.; Kanai, Y.; Kido, M.; Hirokawa, N. Randomization of left-right asymmetry due to loss of nodal cilia generating leftward flow of extraembryonic fluid in mice lacking KIF3B motor protein. Cell 1998, 95, 829–837. [Google Scholar] [CrossRef]
- Taniguchi, K.; Maeda, R.; Ando, T.; Okumura, T.; Nakazawa, N.; Hatori, R.; Nakamura, M.; Hozumi, S.; Fujiwara, H.; Matsuno, K. Chirality in planar cell shape contributes to left-right asymmetric epithelial morphogenesis. Science 2011, 333, 339–341. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, H.; Kondo, S. Rotating pigment cells exhibit an intrinsic chirality. Genes Cells 2015, 20, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Patterson, L.B.; Parichy, D.M. Interactions with iridophores and the tissue environment required for patterning melanophores and xanthophores during zebrafish adult pigment stripe formation. PLoS Genet. 2013, 9, e1003561. [Google Scholar] [CrossRef] [PubMed]
- Volkening, A.; Sandstede, B. Iridophores as a source of robustness in zebrafish stripes and variability in Danio patterns. Nat. Commun. 2018, 9, 3231. [Google Scholar] [CrossRef] [PubMed]
- Inaba, M.; Yamanaka, H.; Kondo, S. Pigment pattern formation by contact-dependent depolarization. Science 2012, 335, 677. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, H.; Kondo, S. In vitro analysis suggests that difference in cell movement during direct interaction can generate various pigment patterns in vivo. Proc. Natl. Acad. Sci. USA 2014, 111, 1867–1872. [Google Scholar] [CrossRef] [PubMed]
- Irion, U.; Frohnhöfer, H.G.; Krauss, J.; Çolak Champollion, T.; Maischein, H.M.; Geiger-Rudolph, S.; Weiler, C.; Nüsslein-Volhard, C. Gap junctions composed of connexins 41.8 and 39.4 are essential for colour pattern formation in zebrafish. Elife 2014, 3, e05125. [Google Scholar] [CrossRef] [PubMed]
- Usui, Y.; Aramaki, T.; Kondo, S.; Watanabe, M. The minimal gap-junction network among melanophores and xanthophores required for stripe pattern formation in zebrafish. Development 2019, 146. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Kondo, S. Changing clothes easily: Connexin41.8 regulates skin pattern variation. Pigment Cell Melanoma Res. 2012, 25, 326–330. [Google Scholar] [CrossRef] [PubMed]
- Shibazaki, Y.; Shimizu, M.; Kuroda, R. Body handedness is directed by genetically determined cytoskeletal dynamics in the early embryo. Curr. Biol. 2004, 14, 1462–1467. [Google Scholar] [CrossRef] [PubMed]
- Woolley, T.E.; Maini, P.K.; Gaffney, E.A. Is pigment cell pattern formation in zebrafish a game of cops and robbers? Pigment Cell Melanoma Res. 2014, 27, 686–687. [Google Scholar] [CrossRef] [PubMed]
- Volkening, A.; Sandstede, B. Modelling stripe formation in zebrafish: An agent-based approach. J. R. Soc. Interface 2015, 12. [Google Scholar] [CrossRef] [PubMed]
- Woolley, T.E. Pattern production through a chiral chasing mechanism. Phys. Rev. E 2017, 96, 032401. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamanaka, H.; Kondo, S. Zebrafish Melanophores Suggest Novel Functions of Cell Chirality in Tissue Formation. Symmetry 2021, 13, 130. https://doi.org/10.3390/sym13010130
Yamanaka H, Kondo S. Zebrafish Melanophores Suggest Novel Functions of Cell Chirality in Tissue Formation. Symmetry. 2021; 13(1):130. https://doi.org/10.3390/sym13010130
Chicago/Turabian StyleYamanaka, Hiroaki, and Shigeru Kondo. 2021. "Zebrafish Melanophores Suggest Novel Functions of Cell Chirality in Tissue Formation" Symmetry 13, no. 1: 130. https://doi.org/10.3390/sym13010130
APA StyleYamanaka, H., & Kondo, S. (2021). Zebrafish Melanophores Suggest Novel Functions of Cell Chirality in Tissue Formation. Symmetry, 13(1), 130. https://doi.org/10.3390/sym13010130