Fluctuating Asymmetry and Population Dynamics of the Common Shrew, Sorex araneus, in Central Siberia under Climate Change Conditions


{kind=link}
Abstract
1. Introduction
2. Material and Methods
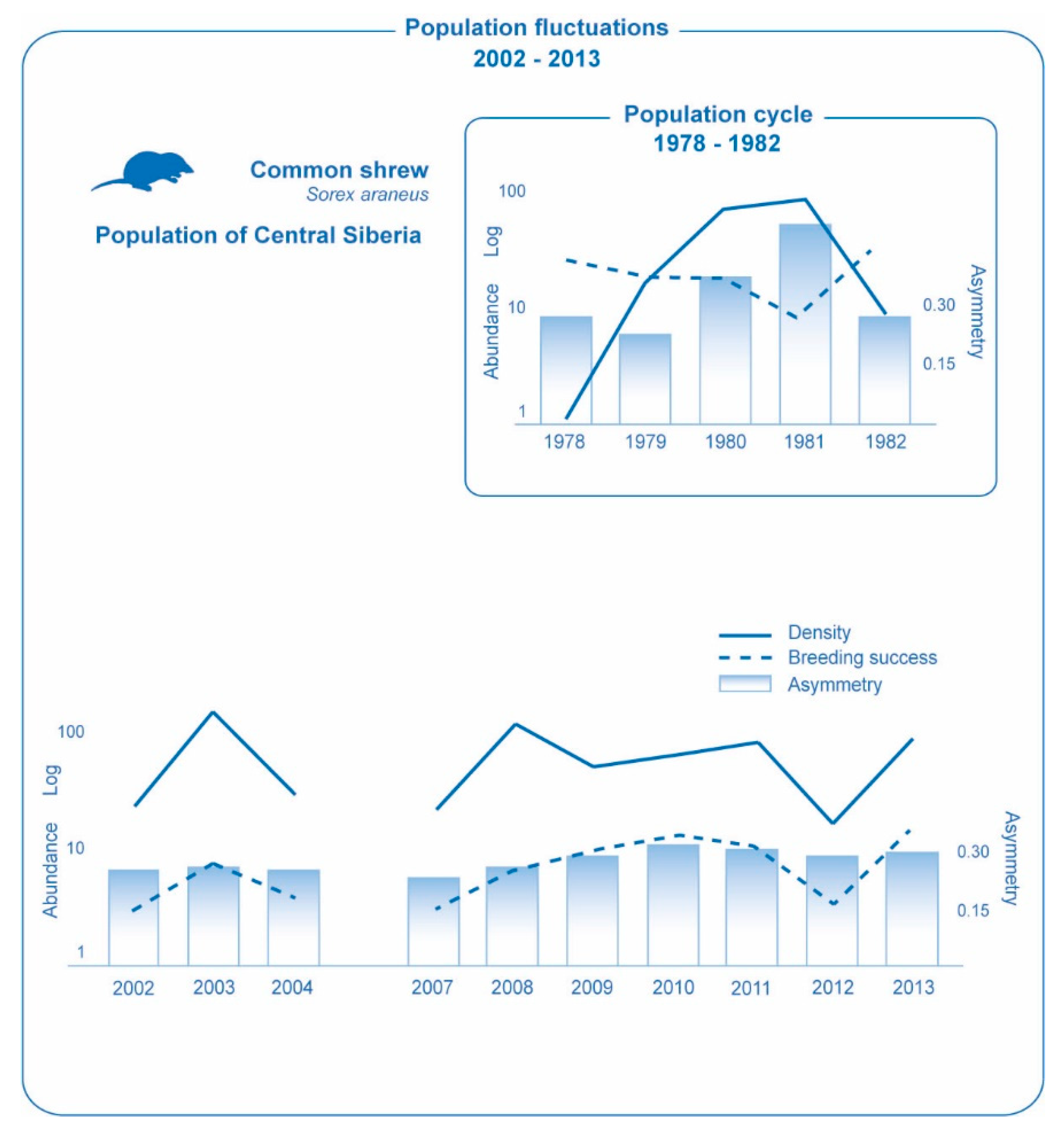
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hansson, L.; Henttonen, H. Rodent dynamics as community processes. Trends Ecol. Evol. 1988, 3, 195–200. [Google Scholar] [CrossRef]
- Kendall, B.E.; Briggs, C.J.; Murdoch, W.W.; Turchin, P.; Ellner, S.P.; McCauley, E.; Nisbet, R.M.; Wood Simon, N. Why do populations cycle? A synthesis of statistical and mechanistic modeling approaches. Ecology 1999, 80, 1789–1805. [Google Scholar] [CrossRef]
- Henttonen, H.; Wallgren, H. Small rodent dynamics and communities in the birch forest zone of northern Fennoscandia. In Nordic Mountain Birch Ecosystem; Wielgolaski, F.E., Ed.; UNESCO Man and Biosphere Series 27; Paris and Parthenon Publishing Group: New York, NY, USA; London, UK, 2001; pp. 261–278. [Google Scholar]
- Ims, R.A.; Fuglei, E. Trophic Interaction Cycles in Tundra Ecosystems and the Impact of Climate Change. BioScience 2005, 55, 311–322. [Google Scholar] [CrossRef]
- Ims, R.A.; Henden, J.-A.; Killengreen, S.T. Collapsing population cycles. Trends Ecol. Evol. 2008, 23, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Pinot, A.; Gauffre, B.; Bretagnolle, V. The interplay between seasonality and density: Consequences for female breeding decisions in a small cyclic herbivore. BMC Ecol. 2014, 14, 17. [Google Scholar] [CrossRef]
- Zárybnická, M.; Riegert, J.; Bejček, V.; Sedláček, F.; Šťastný, K.; Šindelář, J.; Heroldová, M.; Vilímová, J.; Zima, J. Long-term changes of small mammal communities in heterogenous landscapes of Central Europe. Eur. J. Wildl. Res. 2017, 63, 89. [Google Scholar] [CrossRef]
- Giraudoux, P.; Villette, P.; Quéré, J.P.; Damange, J.P.; Delattre, P. Weather influences M. arvalis reproduction but not population dynamics in a 17-year time series. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Mather, K. Genetical control of stability in development. Heredity 1953, 7, 297–336. [Google Scholar] [CrossRef]
- Soule, M.E. Phenetics of Natural Populations. II. Asymmetry and Evolution in a Lizard. Am. Nat. 1967, 101, 141–160. [Google Scholar] [CrossRef]
- Pankakoski, E. Epigenetic asymmetry as an ecological indicator in muskrats. J. Mammal. 1985, 66, 52–57. [Google Scholar] [CrossRef]
- Pankakoski, E. Relationship between some meteorological factors and population dynamics of Sorex araneus in southern Finland. Acta Zool. Fenn. 1985, 173, 287–289. [Google Scholar]
- Zakharov, V.M.; Yablokov, A.V. (Eds.) Developmental homeostasis in natural populations of mammals: Phenetic approach. Acta Theriol. 1997, 4, 92. [Google Scholar]
- Zakharov, V.M.; Pankakoski, E.; Sheftel, B.I.; Peltonen, A.; Hanski, I. Developmental stability and population dynamics in the common shrew Sorex Araneus. Am. Nat. 1991, 138, 797–810. [Google Scholar] [CrossRef]
- Sheftel, B.I. Zonal features of the population of insectivorous mammals of the Yenisei taiga and forest-tundra. In Fauna of the Yenisei Taiga and Forest-Tundra and Natural Zoning; Science: Moscow, Russia, 1983; pp. 184–203. (In Russian) [Google Scholar]
- Sheftel, B.I. Long-term and seasonal dynamics of shrews in Central Siberia. Ann. Zool. Fenn. 1989, 26, 357–369. [Google Scholar]
- Palmer, A.R.; Strobeck, C. Fluctuating asymmetry: Measurement, Analysis, Patterns. Annu. Rev. Ecol. Syst. 1986, 17, 391–421. [Google Scholar] [CrossRef]
- Zakharov, V.M.; Shadrina, E.G.; Trofimov, I.E. Fluctuating Asymmetry, Developmental Noise and Developmental Stability: Future Prospects for the Population Developmental Biology Approach. Symmetry 2020, 12, 1376. [Google Scholar] [CrossRef]
- Frolov, A.V. (Ed.) Second Roshydromet Assessment Report on Climate Change and Its Consequences in Russian Federation. General Summary; Roshydromet: Moscow, Russia, 2014; 56p. [Google Scholar]
- Sheftel, B.I. Role of different mechanisms in type determination of population dynamics for small mammals from boreal forestry zone. In Biological Diversity and Nature Conservation: Theory and Practice for Teaching; KMK: Moscow, Russia, 2010; pp. 107–117. [Google Scholar]
- Zakharov, V.M.; Zhdanova, N.P.; Kirik, E.F.; Shkil’, F.N. Ontogenesis and Population: Evaluation of Developmental Stability in Natural Populations. Russ. J. Dev. Biol. 2001, 32, 336–351. [Google Scholar] [CrossRef]
- Solonen, T. Overwinter population change of small mammals in southern Finland. Ann. Zool. Fenn. 2006, 43, 295–302. [Google Scholar]
- Hansson, L.; Henttonen, H. Gradients in density variations of small rodents: The importance of latitude and snow cover. Oecologia 1985, 67, 394–402. [Google Scholar] [CrossRef]
- Aars, J.; Ims, R.A. Intrinsic and climatic determinants of population demography: The winter dynamics of tundra voles. Ecology 2002, 83, 3449–3456. [Google Scholar] [CrossRef]
- Putkonen, J.; Roe, G. Rain-on-snow events impact soil temperatures and affect ungulate survival. Geophys. Res. Lett. 2003, 30. [Google Scholar] [CrossRef]
- Korslund, L.; Steen, H. Small rodent winter survival: Snow conditions limit access to food resources. J. Anim. Ecol. 2006, 75, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Badyaev, A.V.; Foresman, K.R.; Fernandes, M.V. Rapid environmental change and developmental stability. Vegetation removal causes increased fluctuating asymmetry in a free-living shrew population. Ecology 2000, 81, 336–345. [Google Scholar] [CrossRef]
- Wojcik, J.M.; Polly, P.D.; Wojcik, A.M.; Sikorski, M.D. Epigenetic variation of the common shrew, Sorex araneus, in different habitats. Russ. J. 2007, 6, 43–49. [Google Scholar] [CrossRef]
- Sheftel, B.I. Changes in the species composition in the community of shrews (Soricidae) of the Middle Yenisei taiga. In Theriofauna of Russia and Adjacent Territories, Proceedings of the International Meeting X Congress of the Theriological Society at the Russian Academy of Sciences, Moscow, Russia, 1–5 February 2016; KMK: Moscow, Russia, 2016; p. 460. [Google Scholar]
- Zakharov, V.M.; Sheftel, B.I.; Dmitriev, S.G. Climate change and population dynamics: Possible consequences (with particular references to study of small mammals in Central Siberia). Uspekhi Sovrem. Biol. 2011, 131, 435–439. [Google Scholar]
- Hörnfeldt, B. Long-term decline in numbers of cyclic voles in boreal Sweden: Analysis and presentation of hypotheses. Oikos 2004, 107, 376–392. [Google Scholar] [CrossRef]
- Berteaux, D.; Humphries, M.M.; Krebs, C.J.; Lima, M.; McAdam, A.G.; Pettorelli, N.; Reale, D.; Saitoh, T.; Tkadlec, E.; Weladji, R.B.; et al. Constraints to projecting the effects of climate change on mammals. Clim. Res. 2006, 32, 151–158. [Google Scholar] [CrossRef]
- Bierman, S.M.; Fairbairn, J.P.; Petty, S.J.; Elston, D.A.; Tidhar, D.; Lambin, X. Changes over time in the spatiotemporal dynamics of cyclic populations of field voles (Microtus agrestis L.). Am. Nat. 2006, 167, 583–590. [Google Scholar] [CrossRef]
- Saitoh, T.; Cazelles, B.; Vik, J.O.; Viljugrein, H.; Stenseth, N.C. Effects of regime shifts on the population dynamics of the grey-sided vole in Hokkaido, Japan. Clim. Res. 2006, 32, 109–118. [Google Scholar] [CrossRef]
- Bobretsov, A.V. Population Ecology of Small Mammals of Plain and Mountain Landscapes in the North-East of the European Part of Russia (Populyatsionnaya Ekologiya Melkikh Mlekopitayushchikh Ravninnykh i Gornykh Landshaftov Severo-Vostoka Evropeyskoy Chasti Rossii); KMK: Moscow, Russia, 2016; 381p. (In Russian) [Google Scholar]
- Safronov, V.M. Climate change and mammals of Yakutia. Biol. Bull. Russ. Acad. Sci. 2016, 43, 1256–1270. [Google Scholar] [CrossRef]
- Mertens, S.K.; Yearsley, J.M.; van den Bosch, F.; Gilligan, C.A. Transient population dynamics in periodic matrix models: Methodology and effects of cyclic permutations. Ecology 2006, 87, 2338–2348. [Google Scholar] [CrossRef]
- Brommer, J.E.; Pietiäinen, H.; Ahola, K.; Karell, P.; Karstinen, T.; Kolunen, H. The return of the vole cycle in southern Finland refutes the generality of the loss of cycles through ‘climatic forcing’. Glob. Chang. Biol. 2010, 16, 577–586. [Google Scholar] [CrossRef]
- Korpela, K.; Delgado, M.; Henttonen, H.; Korpimäki, E.; Koskela, E.; Ovaskainen, O.; Pietiäinen, H.; Sundell, J.; Yoccoz, N.G.; Huitu, O. Nonlinear effects of climate on boreal rodent dynamics: Mild winters do not negate high-amplitude cycles. Glob. Chang. Biol. 2013, 19, 697–710. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zakharov, V.M.; Trofimov, I.E.; Sheftel, B.I. Fluctuating Asymmetry and Population Dynamics of the Common Shrew, Sorex araneus, in Central Siberia under Climate Change Conditions. Symmetry 2020, 12, 1960. https://doi.org/10.3390/sym12121960
Zakharov VM, Trofimov IE, Sheftel BI. Fluctuating Asymmetry and Population Dynamics of the Common Shrew, Sorex araneus, in Central Siberia under Climate Change Conditions. Symmetry. 2020; 12(12):1960. https://doi.org/10.3390/sym12121960
Chicago/Turabian StyleZakharov, Vladimir M., Ilya E. Trofimov, and Boris I. Sheftel. 2020. "Fluctuating Asymmetry and Population Dynamics of the Common Shrew, Sorex araneus, in Central Siberia under Climate Change Conditions" Symmetry 12, no. 12: 1960. https://doi.org/10.3390/sym12121960
APA StyleZakharov, V. M., Trofimov, I. E., & Sheftel, B. I. (2020). Fluctuating Asymmetry and Population Dynamics of the Common Shrew, Sorex araneus, in Central Siberia under Climate Change Conditions. Symmetry, 12(12), 1960. https://doi.org/10.3390/sym12121960