1. Introduction
It is timely to bring together and discuss the evidence for asymmetry of brain function in the zebra finch for two reasons. Firstly, the zebra finch is a model species used frequently to understand the links between neural structure and behaviour. Secondly, early research reporting lateral asymmetries in the species was equivocal, largely because it seemed to be at odds with lateralities reported in other avian species and because results of different studies were not always consistent. Therefore, we decided to summarise the available literature showing, or not showing, lateralisation in the zebra finch and to add some data on eye preference to view a predator.
The zebra finch has featured amongst those songbirds investigated for song learning, song production, and perception. Zebra finch song is stereotyped and has a rich spectro–temporal structure, which some researchers have compared to human speech sounds [
1]. Furthermore, male zebra finches learn their song from other birds, by imitating the song of a tutor heard during a sensitive period of development [
2,
3]. These and other aspects of zebra finch song have been studied in considerable detail and compared to speech in humans [
4,
5,
6,
7].
Another feature of song is differential control of its production and processing by the left and right hemispheres. This has been studied in number of avian species, and studies of species other than the zebra finch have demonstrated a dominant role of song centres in the left hemisphere for controlling song production [
8,
9] and differential roles of the hemispheres in perception of song [
10]. However, lateralisation of the song system in the zebra finch seemed not to fit this pattern.
Initially, Nottebohm et al. [
10] cited unpublished observations that indicated little hemispheric asymmetry of song control in the zebra finch, and differing from the canary, zebra finches were reported to have no asymmetry in the size of the left and right hypoglossal nuclei, i.e., the collections of cell bodies with axons that form the hypoglossal nerves, a branch of which innervates the syringeal muscles used to produce song [
11]. However, later research revealed the presence of asymmetry for song in the zebra finch, albeit not the same as that found in other passerine species.
Williams et al. [
12] found right hemispheric control of song production in zebra finches; opposite to the direction of asymmetry reported for other songbirds. Lesioning the auditory areas of the right hemisphere of zebra finches was found to decrease the birds’ ability to process harmonic structure in song [
13]. Floody and Arnold [
14] also reported evidence that the right song system is dominant in the zebra finch. Using functional magnetic resonance imaging (fMRI), Voss et al. [
15] revealed hemispheric asymmetry in neural activity during stimulation by song: significant discrimination between songs was found only in the right hemisphere. Recognition of the zebra finch’s own song versus the song of a conspecific was also found to be biased to the right hemisphere [
16]. All of these studies indicated that perceptual production and processing was a function of the right hemisphere in zebra finches, and thus the asymmetry seemed to be reversed compared to other songbird species studied. However, measuring expression of the immediate early gene ZENK in zebra finches exposed to the auditory and/or visual aspects of courtship, Avery et al. [
17] found left hemispheric dominance (i.e., hearing courtship song and seeing dancing by the courting male causes more neural activity in the left than the right hemisphere). Recent studies have demonstrated that both hemispheres attend to song but to different aspects of it [
1], and that the direction of asymmetry depends on whether the memory of song is old or new [
6].
It is possible that variation in the direction of asymmetry occurs depending on previous exposure to song and to what extent the birds recalled previous exposure to song. Demonstrating that the direction of lateralisation depends on learning and memory, Moorman et al. [
18] reported left-sided dominance of ZENK expression in the higher vocal centre of juvenile males exposed to their tutor’s song but not in those exposed to unfamiliar song. Olson et al. [
6] found that the direction of laterality of song memory depends on strength of learning; the more the zebra finches learnt and remembered the song of their first tutor, the more right lateralised they were, as assessed by ZENK expression. By contrast, the more they learnt from a second tutor, the more left-lateralised they were. Hence, new and old memories of song appear to be located in opposite hemispheres; older memories in the right hemisphere and newer memories in the left hemisphere. In fact, Yang and Vicario [
7] showed that exposure of adult zebra finches to novel hetero-specific sounds (vocalisations of canaries) can shift lateralisation for song processing from the right to the left hemisphere.
Using fMRI measurements of neural processing of song in zebra finches, Van Ruijevelt et al. [
1] provided evidence that the spectral aspect of song is processed in the right hemisphere. By comparison, presentation of song with the spectral component filtered out, but with the temporal component remaining, led to greater neural activity in the left hemisphere [
1]. Hence, the left hemisphere processes the temporal domain of song, whereas the right hemisphere processes the spectral component of song. This role of the left hemisphere in processing temporal aspects of song is supported by finding higher expression of ZENK in regions of the left hemisphere in males when they responded to arrhythmic, but not rhythmic, song [
19].
A female zebra finch hearing a male’s song directed towards her (as compared to the song produced by the male when he is alone) expresses a higher level of activity in the caudocentral–nidopallial region of the left hemisphere and the caudomedial–mesopallial region of the right hemisphere, as shown by functional magnetic imaging (fMRI) and early gene expression [
20]. This result demonstrates that both hemispheres respond to hearing the song, but they process the information in different regions of the forebrain.
None of the above studies considered a possible role of visual asymmetry in association with lateralised auditory perception, learning, and production of song. Although it was known that fledgling zebra finches learn their tutor’s song only when they can see the tutor [
21], possible eye preference and lateralisation was not studied at this time. It was in the late 1970s that asymmetry of visual behaviour in an avian species was discovered (i.e., in the domestic chick [
22]). The first asymmetry recorded for visual behaviour of the zebra finch concerned courtship. Workman and Andrew [
23] reported evidence that during courtship males show a preference to use the right eye when viewing their female. This asymmetry was found by measuring the approach of the male to his female partner as he moved along a perch. Using a different method (viz., direction of movement in a circular corridor around cages containing females), Ten Cate et al. [
24] failed to find any evidence for asymmetry, a result which Workman and Andrew [
25] attributed to the males being tested with females that were not necessarily their own partners (see reply by Ten Cate [
26]). A subsequent study by George et al. [
27] resolved this debate by measuring brain activity during courtship singing by male zebra finches, and by measuring the amount of singing when only the left or right eye could be used. Males sang more song motifs when they could see a female with their right eye only than they did when they could see her with their left eye only. However, birds tested binocularly sang with more motifs than either of these groups.
Right eye preference during courtship approach to the female and in producing song with more motifs implies that this behaviour depends on the left hemisphere, since inputs from the eyes are largely processed by the contralateral hemisphere. In the study by George et al. [
27], brain activity during courtship singing was assessed by assaying the expression of the immediate early genes, egr-1 and c-fos, in the optic tecta. The optic tectum on each side of the brain is the first relay station for visual inputs from the retina, and each eye sends its inputs to the contralateral optic tectum [
28]. In males able to view the female with both eyes, neural activity was found to be higher in the left than the right optic tectum [
27], and also in other regions of the left hemisphere [
29]. This result is consistent with the preference to use the right eye during courtship singing, as found by Workman and Andrew [
23]. It might be explained by the ability of the left hemisphere (and right eye) to sustain attention on a preferred and familiar stimulus [
30,
31,
32] and to maintain attention on a stimulus towards which a motor response is planned [
27].
A right eye preference during courtship by zebra finch males was confirmed by testing birds with monocular eye patches [
33]. Birds that could see with their right eye only courted females more than those using their left eye only. The former also expressed preferences for orange-beaked (high quality) females over grey-beaked (low quality) females, whereas birds using their left eye only expressed no such preference [
33].
Asymmetry in the motor behaviour of the zebra finch has also been reported. Alonso [
34] measured side biases in allopreening and bill wiping and found significant preference to turn to the right side of the body for both of these responses. Since no left-right bias had been found for turning in a Y-maze, Alonso [
34] interpreted her results as reflecting visual and not motor asymmetry, arguing that allopreening demands visual precision or that there is a need to keep the bird being preened in the right visual field. In the case of bill wiping, it was argued that the right-side preference reflects a right eye preference for pecking at food.
The zebra finch also uses the right eye and left hemisphere when it pecks at grain scattered amongst distracting pebbles. Alonso [
35] found zebra finches tested with monocular eye patches could distinguish grain from pebbles when they were able to use their right eye only but not when they were able to use their left eye only [
35]. This asymmetry reflected the role of the left hemisphere in performing this task. Indeed, this result replicated earlier research on domestic chicks, which had shown specialisation of the left hemisphere for learning a pebble-grain discrimination task [
22,
36], found also in pigeons [
37,
38].
Since attention and response to predators have been shown to be a specialisation of the right hemisphere, and left eye, in other avian species (domestic chicks [
30], Australian magpies [
39,
40], as well as in amphibian [
41] and mammalian [
42] species), it seemed appropriate to test zebra finches for possible lateralisation of attention to a predator. Lombardi and Curio [
43] have described the zebra finch’s response to seeing a live owl as including side to side movements of the head which allow monocular fixation. Fixation refers to holding the head still after a rapid movement of the head.
We therefore needed to assess not only single monocular fixations of a stimulus, as in the species tested so far, but also biocular alternating fixations. Biocular alternating fixation refers to swapping from the lateral field of one eye to the lateral field of the other eye, as defined by Butler et al. [
44], and to be distinguished from binocular fixation that uses the frontal visual field of both eyes.
Zebra finches have many predators in their natural environment, including snakes, monitor lizards, raptors, and a variety of other avian species [
45], and they are particularly vulnerable to predation when they are fledglings [
46]. Snakes and monitor lizards are especially well-documented nest predators of the species [
45,
47]. Therefore, we decided to present a taxidermic specimen of a lace monitor lizard,
Varanus varius, to zebra finches.
The zebra finch has laterally positioned eyes, and in the horizontal plane, each eye has a visual field of 170° [
48]. The binocular field is 30° to 40° in front and the optical axis and fovea are at 62° from the sagittal axis of the head [
48]. Hence, acute vision, especially of moving stimuli, requires monocular vision, whereas the binocular field is myopic. Even grain is viewed with the lateral monocular field before the bird pecks [
48]. Potential predators are also viewed in the lateral, monocular field of vision.
Therefore, our aim was to assess eye use and eye preference to view a model predator and so determine whether or not the zebra finch shows the same left eye (right hemisphere) preference to view this stimulus as found in other vertebrate species.
2. Materials and Methods
2.1. Subjects
Twenty adult zebra finches were purchased from a breeder who housed the birds in outdoor aviaries in rural NSW, Australia. They had been exposed to species occurring naturally in this environment. These species included predators (monitor lizards and raptors), rodents, and other free-ranging animals (i.e., dogs, cats, and other native birds). At the University of New England, the zebra finches were housed in same-sex groups, in four aviaries (1.5 m × 1 m × 2 m) located in a single room.
Each aviary was furnished with branches for perching and nest baskets. Ambient temperature was maintained within a range of 18–27 °C. The light cycle was 13L:11D, the main lights were turned on at 06.00 h and off at 19.00 h. A small lamp with a 40 W globe was placed in the centre of the room and was switched on at 19.00 h and switched off at 19.30 h. The latter was to provide a cue to the birds to begin to roost and to provide some light for a brief period once the main light source had been switched off. A Hitachi 40 W fluorescent light with an UV output of 7.5 was activated for 30 min from 07.30 h to 08.00 h daily to provide a source of UV light. Food, water, and cuttlefish bone were supplied ad libitum. The seed used was a mixture of two commercial brands of food for finches (Lovitt and Trill). Vitamin and calcium supplements were provided once every 2 weeks and lettuce was provided once a week. The cage floor was lined with newspaper and replaced once a week.
Individual birds were identified using the following features: Female zebra finches were identified by noting their colour morph (wild type, fawn, or white morph), beak colour and size, and shape of the markings on their heads/faces. Males were first categorised by their colour morph and then the characteristics that were used to identify individual males of the same morph were beak colour, size, and shape of chest band, presence or absence of a white patch of feathers under beak, and the colour or the pattern underneath the wings. All of these markings were individually distinctive, and since the birds were housed in small home groups, identifying individuals was accurate. Ring bands were not used to identify individual birds since this could have influenced their behaviour [
49].
Housing and testing of the zebra finches was conducted with the authority of the Animal Ethics Committee of the University of New England (AEC numbers 06/091, 07/014, 06/090).
2.2. Testing Room and Aviary
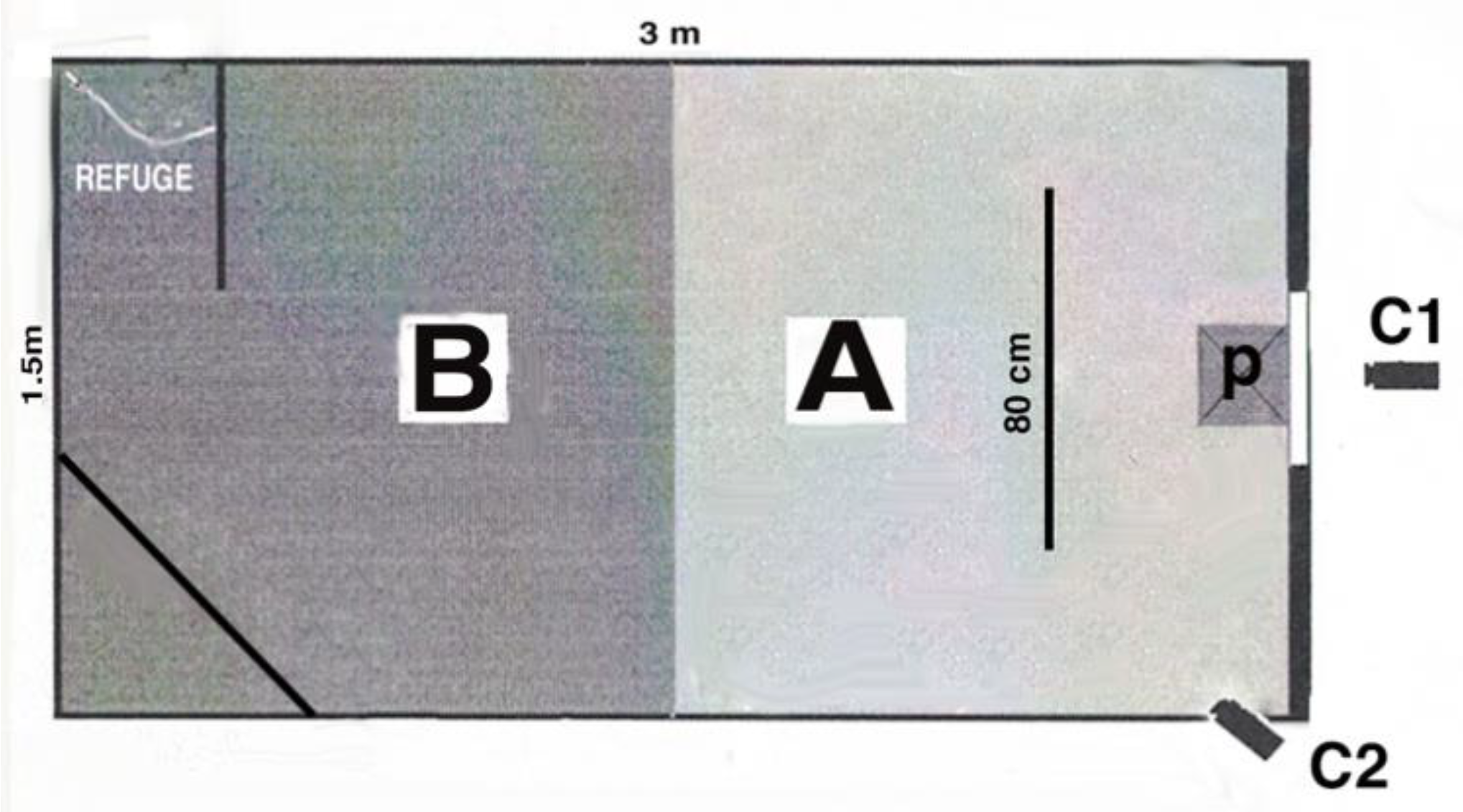
Testing occurred in the birds’ second year of life after some pilot tests had been conducted (see below) and 9 months after the birds had been purchased and housed at the university. All experiments were conducted in an aviary (3 m × 1.5 m × 2 m) located in a room separated from where the home aviaries were kept (
Figure 1). The testing aviary was divided into two virtual sections of equal size (1.5 m × 1.5 m × 2 m). Section A was the half of the aviary where the stimuli were presented, and Section B was the other half of the aviary. Two perches were placed in Section B, in the corners of the aviary furthest from location of the stimulus, from one of which the bird could see into Section A of the aviary and could see the stimulus at a distance. The other perch in Section B was located behind a visual barrier to provide a place of refuge at some distance from the stimulus.
Section A contained some branches spanning from the floor to the roof of the aviary and located at the border of Section A and B. The main perch was 80 cm in length, 40 cm from the shorter side of the aviary where the stimulus was presented and at 30 cm above a platform on which the stimulus was presented (i.e., the bird was approximately 50 cm from the platform and stimulus). The platform was 40 cm × 60 cm, located midway in the shorter side of the cage and 140 cm from the floor of the aviary.
Two digital video recorders (Panasonic NVGS35) were located outside the aviary (120 cm from it) behind the stimulus presentation area. One video recorder was placed 30cm above the platform so that it recorded a clear, close-up view of the bird on the main perch near the stimulus. The other was placed so that a wide-range view incorporated most of the aviary with the exception of the refuge area. The experimenter was located behind a visual barrier and could observe the finches via a monitor.
2.3. Testing
Some pilot tests were conducted using birds tested alone but this was found to be unsuccessful because separation from the group caused stress. The isolated birds tended to freeze and not move or feed in the testing aviary and this was considered undesirable for the bird, and counterproductive for the testing procedure. Therefore, pairs were tested (each pair treated as N = 1) in order to avoid confounding our results with effects of stress through social isolation [
50]. During these preliminary tests, the birds were between 6 and 12 months of age.
In all cases, the pairs were same-sex cage mates. Zebra finches in captivity may form same-sex pair bonds [
51]. To determine whether individuals had formed a bond, each cage group was observed for 30 min per day over four consecutive days and the interactions between the individuals were noted. Seven bonded pairs (five same-sex male pairs and two same-sex female pairs) were identified. Individuals that had formed a bond were seen allopreening and no agonistic behaviour between the pair was recorded. By contrast, agonistic events were regularly directed toward individuals not part of the pair. Only those that had formed pair-bonds were selected for testing
Collecting the finches for transport to the testing room and testing aviary was achieved by turning off the lights. Since zebra finches have poor eyesight in dim light, they could be collected from their perches without undue stress (no flight response). The experimenter was able to take each finch and place it in a small transport box and then release it into the testing aviary. The pair then remained in the test-cage for 5 days before the actual tests were conducted. This allowed them to adjust to the new surroundings before the stimuli were presented.
To present the stimuli, the experimenter placed the stimulus in the aviary through a door next to the presentation platform. The stimulus was retrieved using the same procedure. Note, only the arm of the experimenter was visible to the birds briefly while the stimulus was placed on the platform or retrieved from it. The experimenter was otherwise visually isolated from the birds throughout testing. Behaviour was scored from the video-footage.
2.4. Stimuli and Their Presentation
The non-threatening stimulus was a white opaque plastic container, hereafter referred to as a jar, with a red lid (height 16 cm, base diameter 9 cm, lid diameter 5 cm;
Figure 2A). The zebra finches had been previously exposed to similar containers although not this particular one.
The predator used, as already indicated, was a taxidermic specimen of a monitor lizard,
Varanus varanus, 125 cm in length (
Figure 2B,C).
Total time for a test was 15 min. At the end of a 5-min pre-test period, and when both the birds were in Section B of the cage, the experimenter opened a small door cut into the aviary wire and placed the stimulus on the platform. The test period was of 5-min duration, after which the stimulus was removed and then there was a post-test period of 5 min. Behaviour performed in all three periods was recorded on videotape.
Each stimulus was presented to each pair once per day at 9.30 a.m. or 10.30 a.m. for a total of 6 days. The order of presentation was random.
2.5. Data Collection
For a zebra finch to fixate a stimulus monocularly, it needs to make an exaggerated head movement. This provided an opportunity to score whether there was an eye preference to view the stimulus. After approaching a stimulus (i.e., landing on the perch directly in front of the stimulus), the zebra finches performed monocular fixation movements, described by Lombardi and Curio [
43,
52]. This was classified as inspection behaviour. To ensure that the bird was fixating the stimulus monocularly (i.e., for 1 or more seconds) during an inspection event, the angle of the bird’s beak to the stimulus had to be 90° ± 20° (
Figure 3).
Scoring used playback of video recordings which allowed frame-by-frame analysis, particularly for scoring eye use (Numbers 4 and 5 below). The following behaviour was scored from the videotapes:
Latency for the first bird to enter Section A (i.e., the Section in which the stimulus was presented); see
Figure 1.
Time spent in Section A in the pre-test, test, and post-test periods. Each bird was scored individually, and the final tally was the total for the pair.
Number of visits into Section A.
Number of monocular fixation events and alternating monocular fixation bouts per pair. These were scored only when the bird was in Section A on the perch directly in front of the stimulus.
Eye used in each fixation event of at least one second duration.
2.6. Statistical Analysis
Data were analysed for normality and equal variances and if the assumptions for parametric tests were not met, non-parametric statistics were conducted. The non-parametric data were analysed using Friedman’s test with testing day as the repeated measure. Post hoc analyses used two-sample Wilcoxon signed ranks tests. The parametric data were analysed using t-tests. Note that the sample size used was the number of pairs, not the number of individuals.
4. Discussion
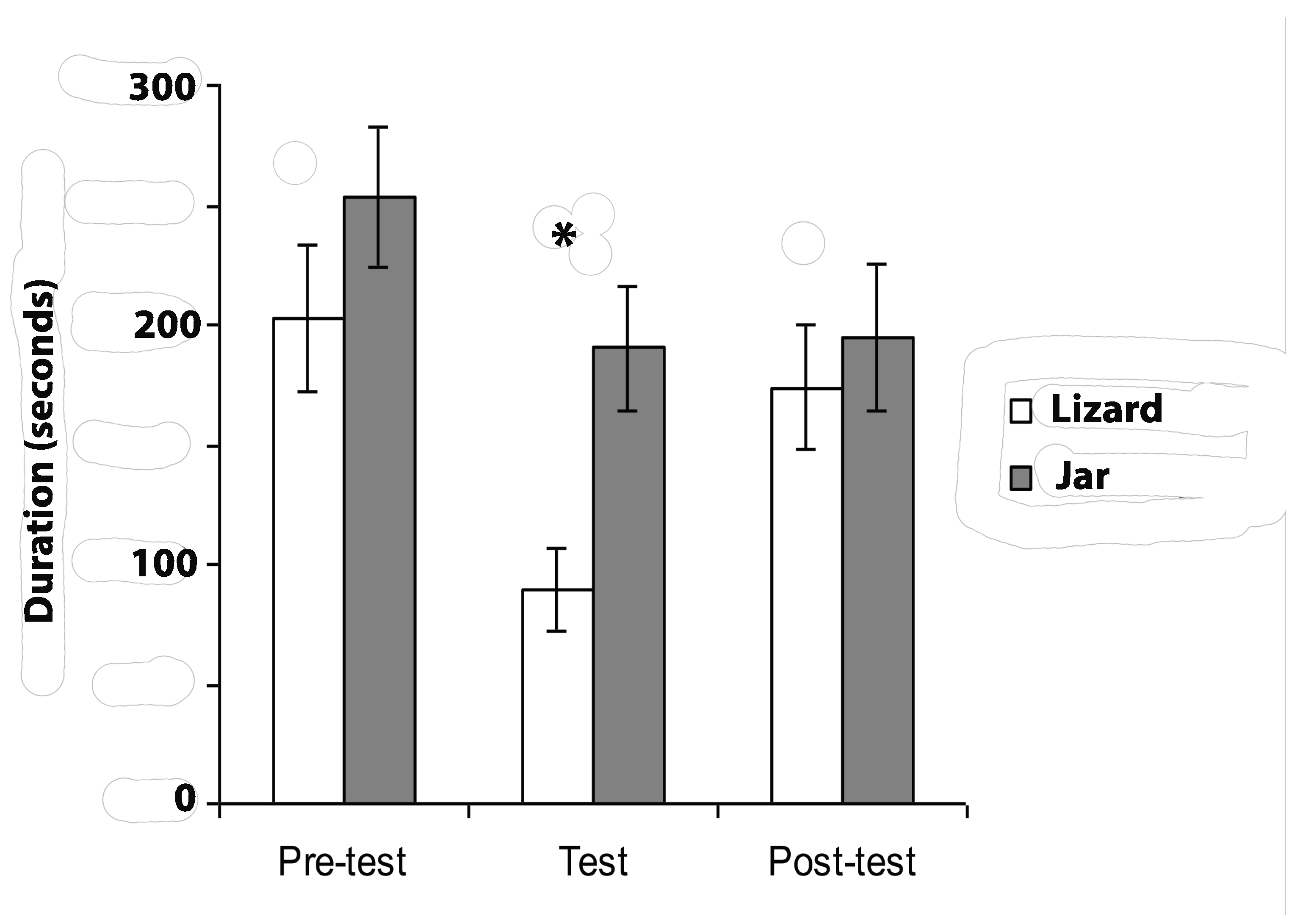
First it was important to obtain evidence that the zebra finches perceived the lizard, and not the jar, as a threat. Therefore, approach to the stimuli was measured. Latency to move into Section A after presentation of the stimulus was one such measure. This latency was twice as long when the lizard was presented than it was when the jar was presented. Also, the birds spent significantly less time in Section A of the cage when the lizard was presented compared to the time that they spent in Section A during the pre-test and post-test periods. By contrast, no avoidance of Section A occurred when the jar was presented. Both of these measures confirmed that the zebra finches perceived the lizard, and not the jar, as a threat (i.e., recognised it as a predator).
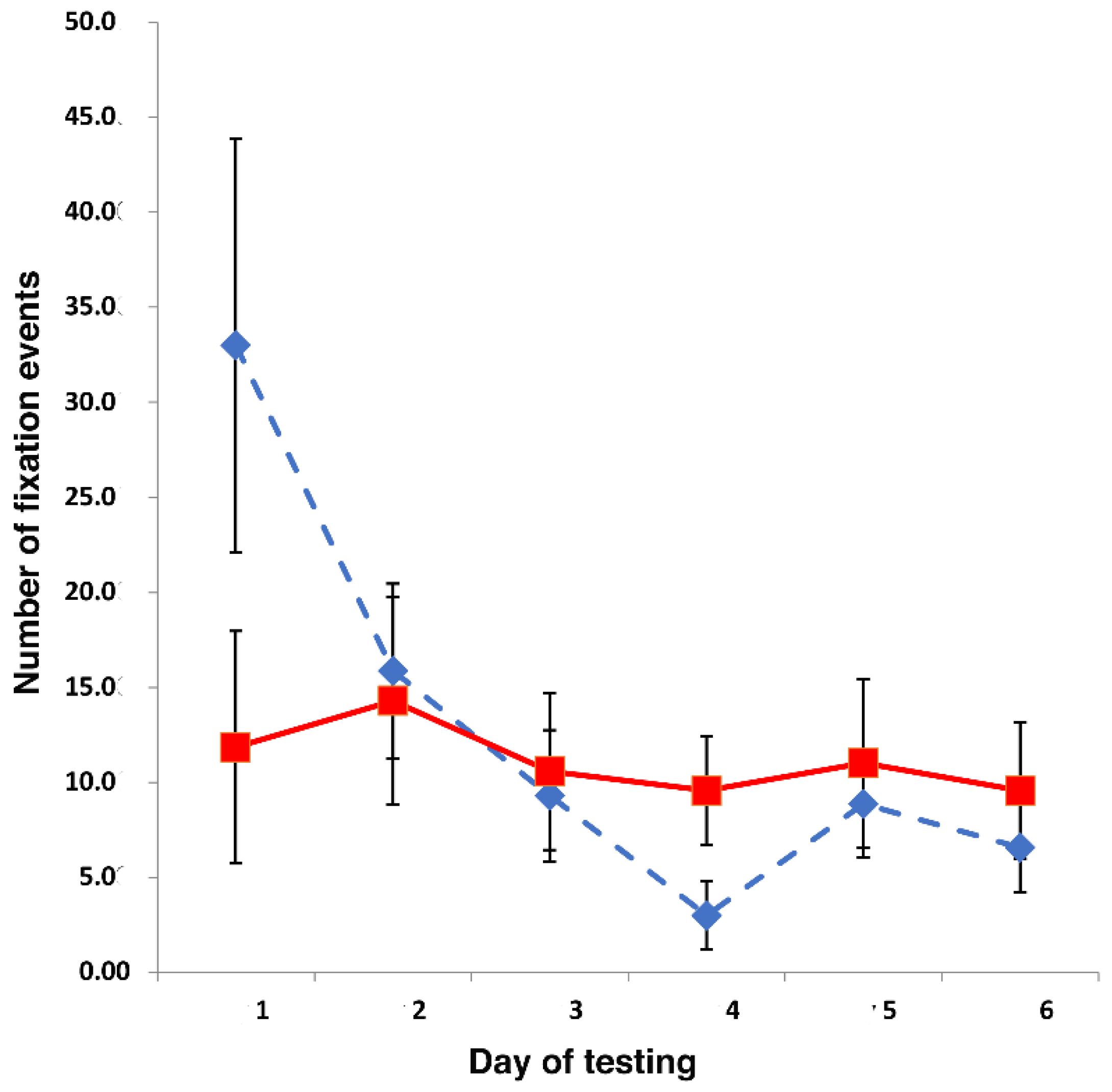
On the first presentation of the jar, the birds fixated it more times than they did the lizard or than they did on any subsequent presentation. This suggests that, on first presentation, the jar might have been perceived as somewhat novel, but not threatening. By contrast, response to the lizard remained unchanged across the 6 days of testing; this lack of habituation of response to the lizard is likely to be caused by elevated levels of fear induced by this potential predator [
53]. Despite these differences in response to the two stimuli, the total number of times the birds fixed the stimuli, over all days of testing, were the same for the lizard and the jar. However, use of the eyes to view the lizard compared to the jar did differ. There was a significant preference to view the lizard using the left eye but no significant eye preference to view the jar. This finding of a left eye preference to view the lizard was reinforced by occasions when the birds caught sight of this stimulus with the right eye and immediately switched to fixate it with the left eye. Although switches were also observed on presentation of the jar, there was no directional preference to switch from right to left eye versus from left to right eye to fixate the stimulus.
Lombardi and Curio [
43,
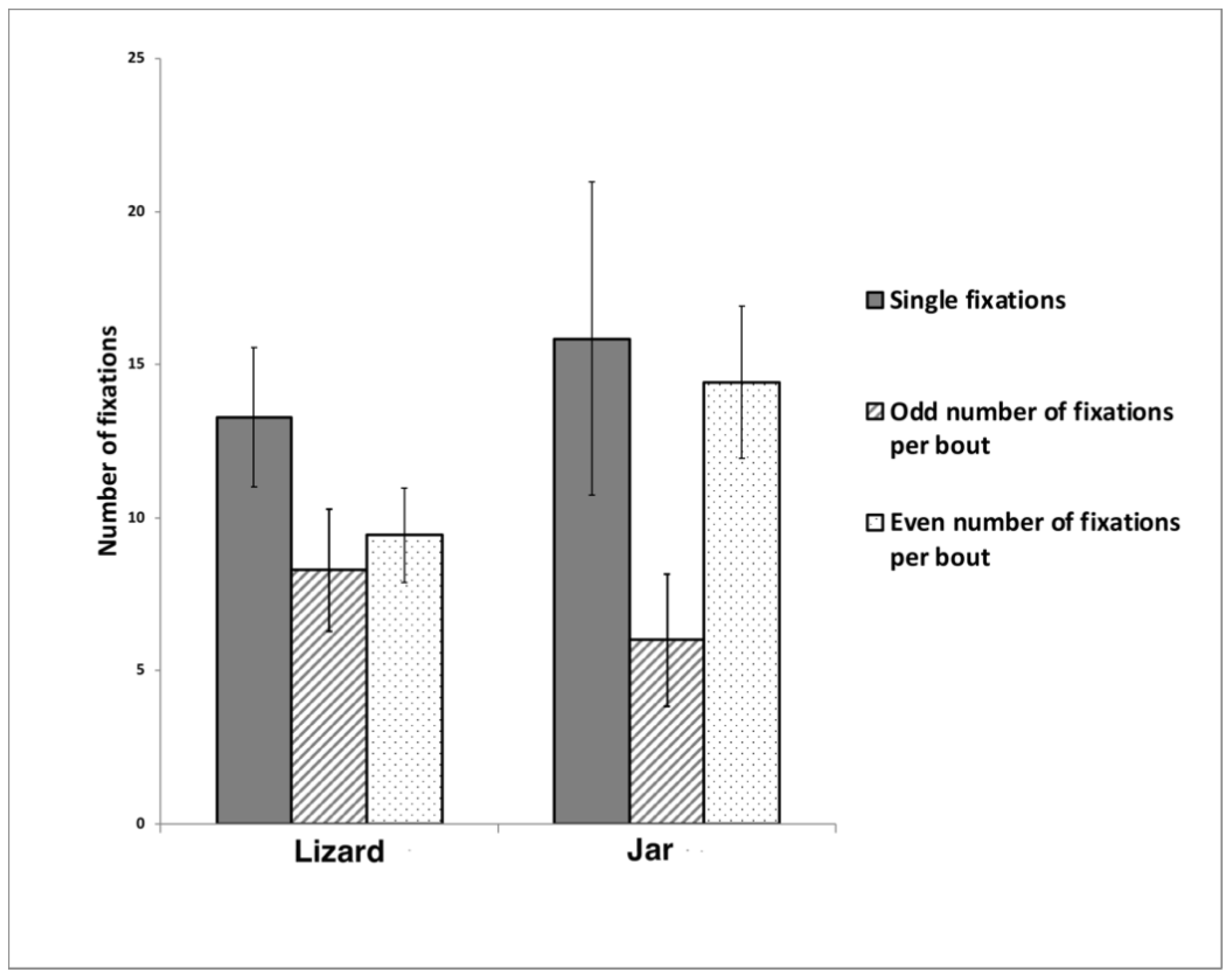
52] described the zebra finch’s response to seeing a live owl as including side to side movements of the head which allow monocular fixation. This sequential use of the left and right monocular visual fields was also observed in our study but single monocular fixations with one eye were also common. When the stimuli were viewed using monocular alternating fixation bouts, these differed for viewing the lizard versus the jar. Such bouts to view the jar most often involved a fixation with one eye and then the other, whereas for viewing the lizard the birds often looked with one eye (left eye preference), then the other eye, and again with the first eye (left eye again).
In fact, the finding of a left eye preference to view the lizard versus no preferred eye to view the jar remained consistent across all days of testing. A preference to use the left eye to view a predator has also been reported in other avian species. Australian magpies,
Gymnorhina tibicen, display a left eye preference to view their main predator, a wedge-tailed eagle, prior to withdrawing from it [
39,
40,
54], and also in our tests the zebra finches moved away from the monitor lizard after they had inspected it from the vantage point of the perch in Section A of the cage. Domestic chicks show a left-eye preference to monitor overhead for a model of a raptor, and this contrasts with use of the right eye to search for grains of food [
30,
55]. Domestic chicks also show a left eye preference to monitor biological motion [
56], a finding that fits well with use of the left eye to view predators.
Such left eye preference for detecting and viewing predators extends to other vertebrate species. A Dasyurid marsupial has been shown to respond more strongly to a predator, a model snake, seen in its left lateral field of vision compared to the same stimulus seen its right lateral field of vision [
42], and the same has been found in the cane toad,
Bufo marinus [
41]. Therefore, together with our current finding in the zebra finch, the evidence for left eye preference to view a predator before responding by withdrawing from it is consistent across a range of species. Since inputs from the left eye are processed almost entirely by the right hemisphere, left-eye preference reflects specialisation of the right hemisphere for processing of and retreating from predators (summarised in [
32]).
Butler et al. [
44] also found a left eye preference in starlings viewing model predators (hawks), but they used a different method—a number of fixations of the stimulus made with one eye (using different regions of the retina) before switching to view the stimulus with the other eye. The left eye preference measured in this way was not specific to viewing predators since it was also found when the stimulus was simply a patch of grass similar to one on which one of the predators had been presented. The researchers suggested that the left eye preference resulted from a higher concentration of single cones in the left eye compared to the right eye [
57]. However, we think this is unlikely to be the sole reason for the left eye preference, since by intracranial injections into the left or right hemisphere of the chick, it has been shown that asymmetry of eye use for visual discrimination learning depends on hemispheric differences in processing stimuli and controlling response (viz., right eye and left hemisphere for visual discrimination learning [
22], summarised in [
58]). Furthermore, if the response to a predator is close approach with contact during attack rather than withdrawal, as is often the case in magpies, there is a preference for use of the right eye and left hemisphere just before attacking [
40].
Considerable research of the regions of the zebra finch brain receiving visual inputs has been undertaken. As mentioned in the Introduction, most of the visual input from an eye is processed by the contralateral hemisphere. In fact, the first relay station for visual inputs in the main visual pathway is entirely to the optic tectum contralateral to the eye. By far the majority of neurones from each optic tectum projects to the ipsilateral nucleus rotundus, although a minority do cross the midline to project to the contralateral nucleus rotundus [
59]. Electrophysiological recordings of neurones in the nucleus rotundus have shown that, while there are neurones that respond to inputs from both the ipsilateral and contralateral eye, there are no neurones that respond exclusively to the ipsilateral eye [
60]. Consequently, Schmidt and Bischof [
60] suggested that at rotundal level, there is inhibition of input from the ipsilateral eye by contralateral input and thereby only one eye engages the bird’s attention.
From each nucleus rotundus projections go to the forebrain only on the same side. Therefore, inputs from one eye are processed on the contralateral side of the forebrain (in the contralateral hemisphere). While one eye is attending to a particular stimulus, visual processing of inputs by the other eye is suppressed, as shown by Voss and Bischof [
61]. In fact, the suppressed eye moves in a saccade counter to the attending eye so that foveal inputs from only the attending eye are processed at forebrain level [
62]. Hence, we may deduce that attention to a predator using the left eye engages the right hemisphere and suppresses information coming from the right eye.
Taking all of the available evidence of asymmetry in zebra finches into account indicates that lateralised brain function is the same as in other avian species. At the least, use of the right eye and left hemisphere to discriminate grain from pebbles [
34] and the left eye and right hemisphere to attend to predators is consistent with the pattern of lateralisation in chicks, magpies [
63], and other avian species (see Introduction). One apparent discrepancy remains, and that concerns copulation and courtship. Whereas male zebra finches use their right eye to approach a female in courtship and their songs have more motifs when they see the female by using their right eye (discussed in the Introduction), the evidence from the domestic chick is that the left eye and right hemisphere control the copulation response [
64]. It is possible that the difference depends on courtship behaviour versus actual performance of copulation. During courtship performance, copulation behaviour must be suppressed and that could be achieved by the left hemisphere’s ability to suppress the right hemisphere [
31], and hence the right eye/left hemisphere is used during courtship. Consistent with this explanation, in sage-grouse Krakauer et al. [
65] found significant left-eye preference during courtship only in males that mated successfully and not in those that were unsuccessful in mating.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






