
A Monoclonal Antibody to Human DLK1 Reveals Differential Expression in Cancer and Absence in Healthy Tissues

Abstract
:1. Introduction
2. Results and Discussion
2.1. Antigen Expression and Characterization
2.2. Antibody Isolation and Characterization
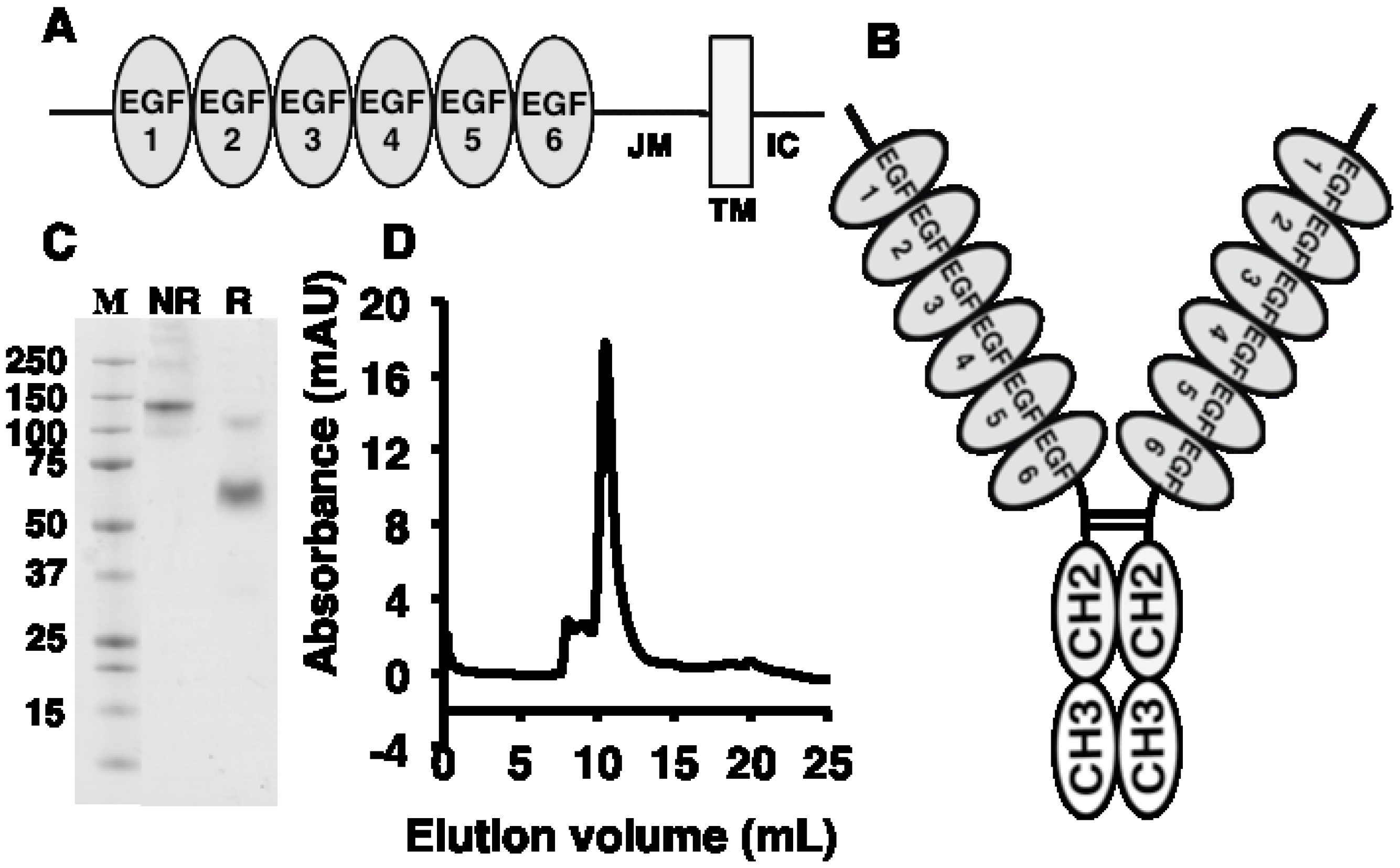

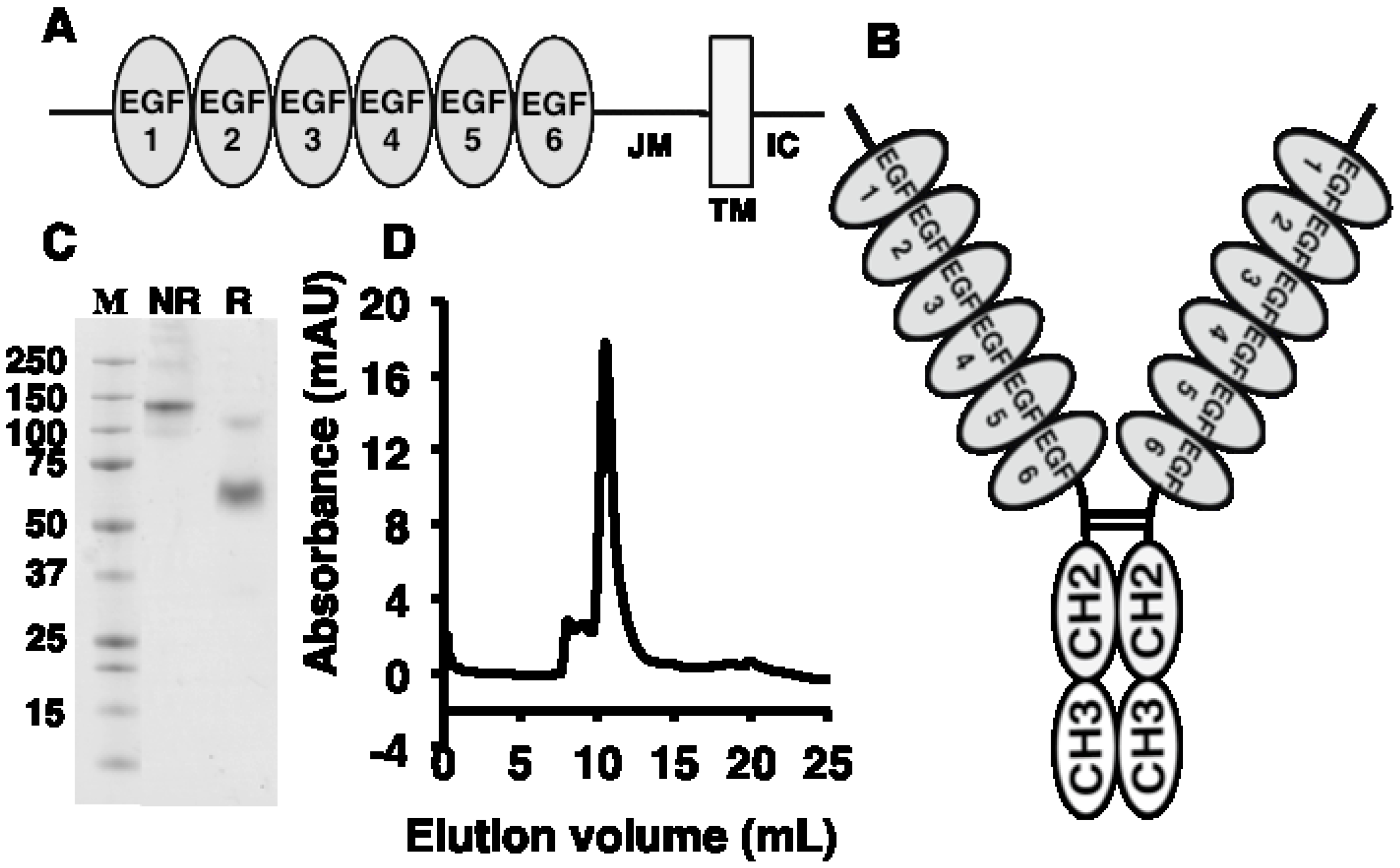{kind=link}
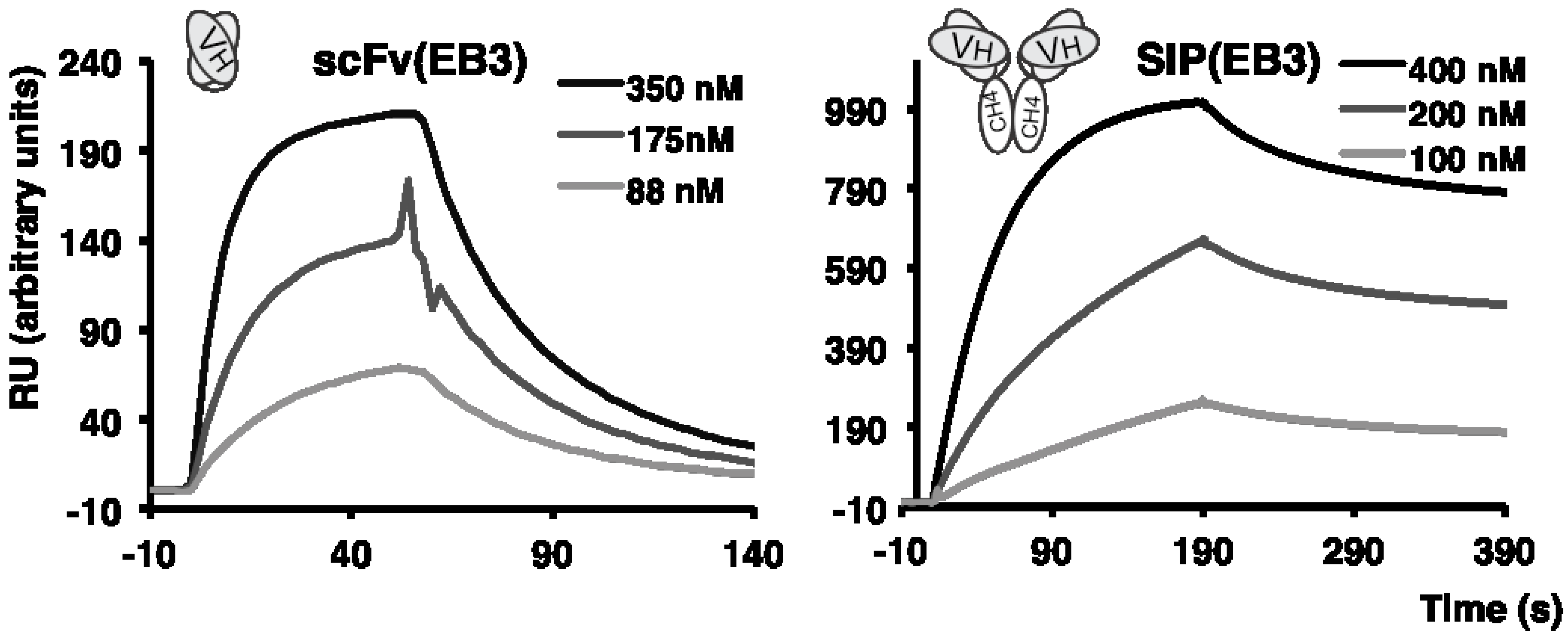{kind=link}
{kind=link}
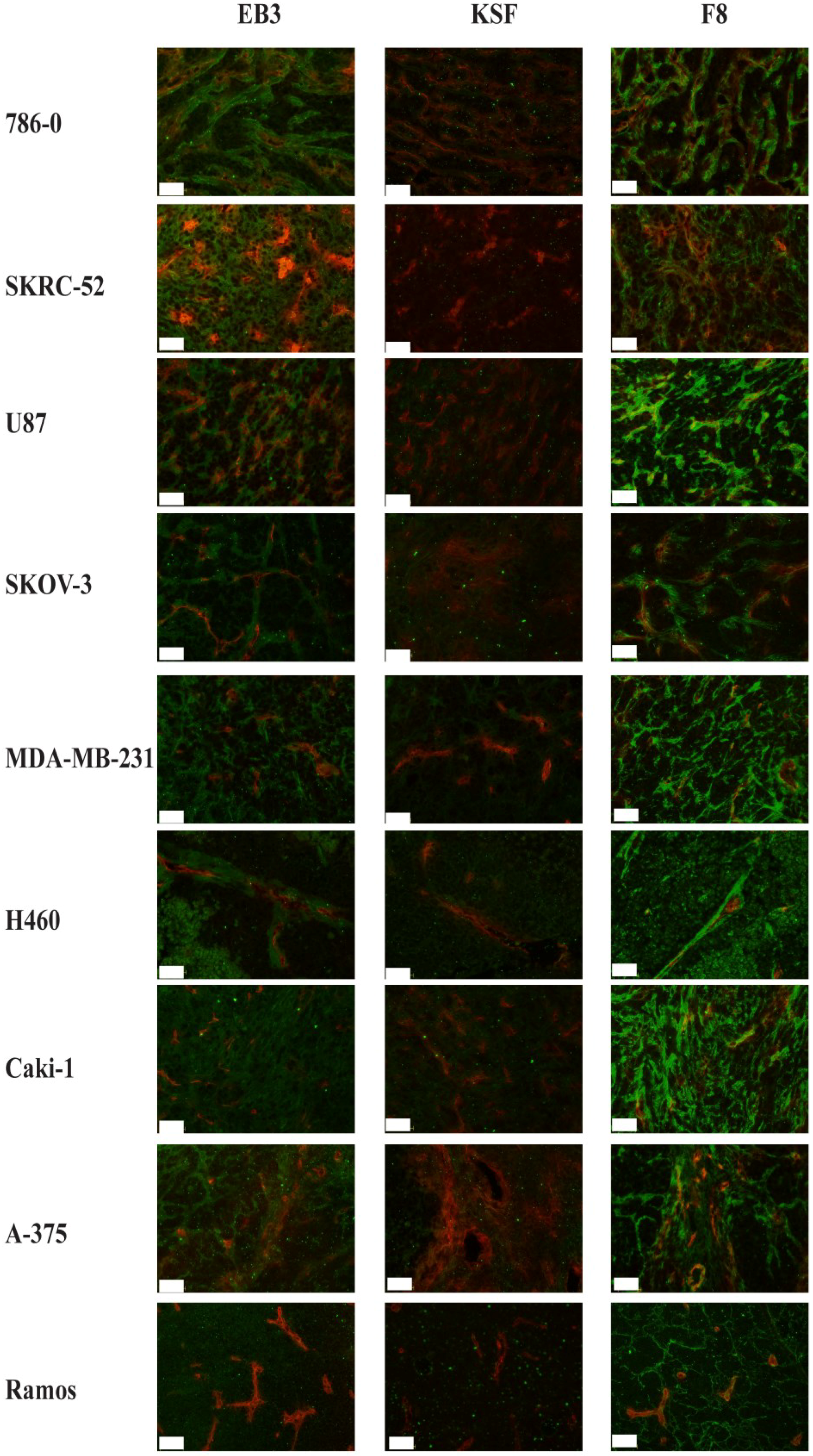{kind=link}
{kind=link}
| DLK1-Fc | Sequence | Start Sequence Position | End Sequence Position |
|---|---|---|---|
| 1 | AECFPACNPQNGFCEDDNVCR | 1 | 21 |
| 2 | KDGPCVINGSPCQHGGTCVDDEGR | 104 | 127 |
| 3 | DGPCVINGSPCQHGGTCVDDEGR | 105 | 127 |
| 4 | CRCPAGFIDK | 171 | 180 |
| 5 | CPAGFIDK | 173 | 180 |
| 6 | RALSPQQVTR | 225 | 234 |
| 7 | LPSGYGLAYR | 235 | 244 |
| 8 | KTPLLTEGQEPK | 272 | 283 |
| 9 | TPLLTEGQEPK | 273 | 283 |
| 10 | THTCPPCPAPELLGGPSVFLFPPKPK | 288 | 313 |
| 11 | DTLMISR | 314 | 320 |
| 12 | TPEVTCVVVDVSHEDPEVK | 321 | 339 |
| 13 | FNWYVDGVEVHNAK | 340 | 353 |
| 14 | EEQYNSTYR | 358 | 366 |
| 15 | VVSVLTVLHQDWLNGK | 367 | 382 |
| 16 | VVSVLTVLHQDWLNGKEYK | 367 | 385 |
| 17 | ALPAPIEK | 392 | 399 |
| 18 | EPQVYTLPPSR | 410 | 420 |
| 19 | EPQVYTLPPSRDELTK | 410 | 425 |
| 20 | NQVSLTCLVK | 426 | 435 |
| 21 | GFYPSDIAVEWESNGQPENNYK | 436 | 457 |
| 22 | TTPPVLDSDGSFFLYSK | 458 | 474 |
| 23 | LTVDKSR | 475 | 481 |
| 24 | WQQGNVFSCSVMHEALHNHYTQK | 482 | 504 |
| 25 | SLSLSPGK | 505 | 512 |
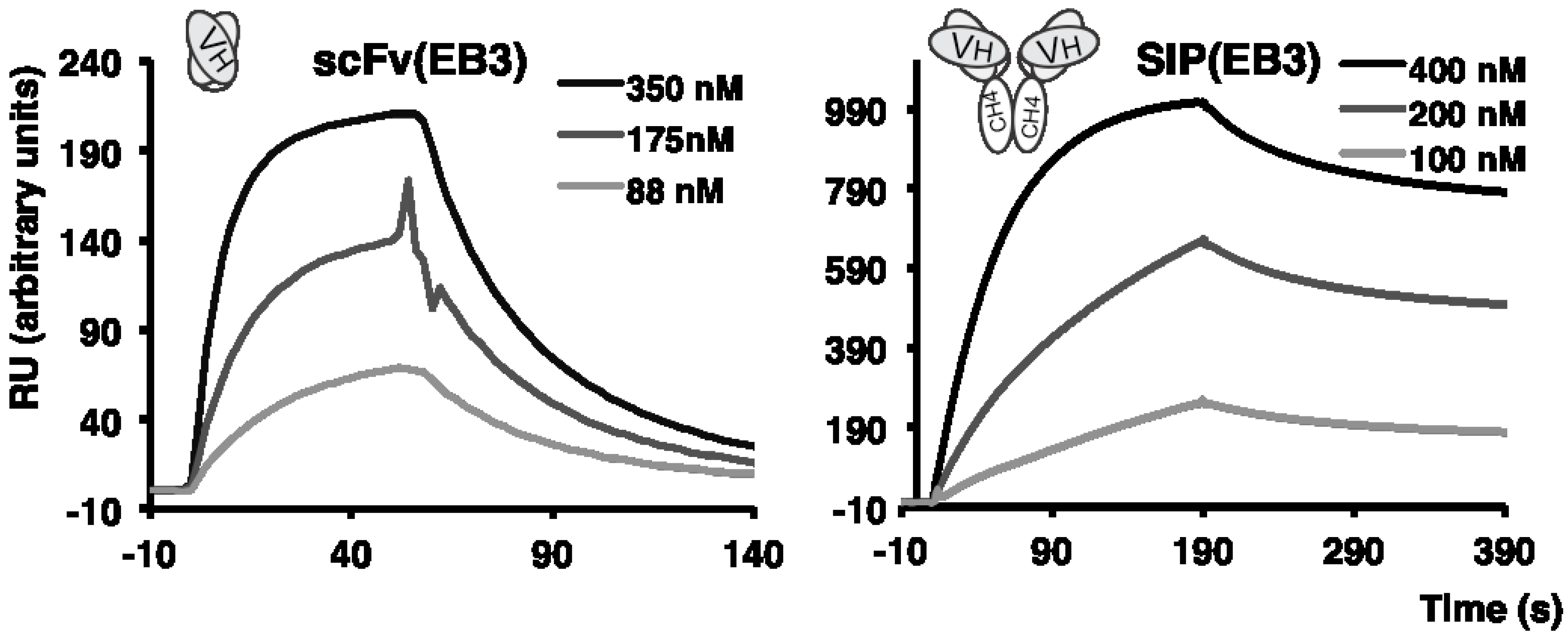
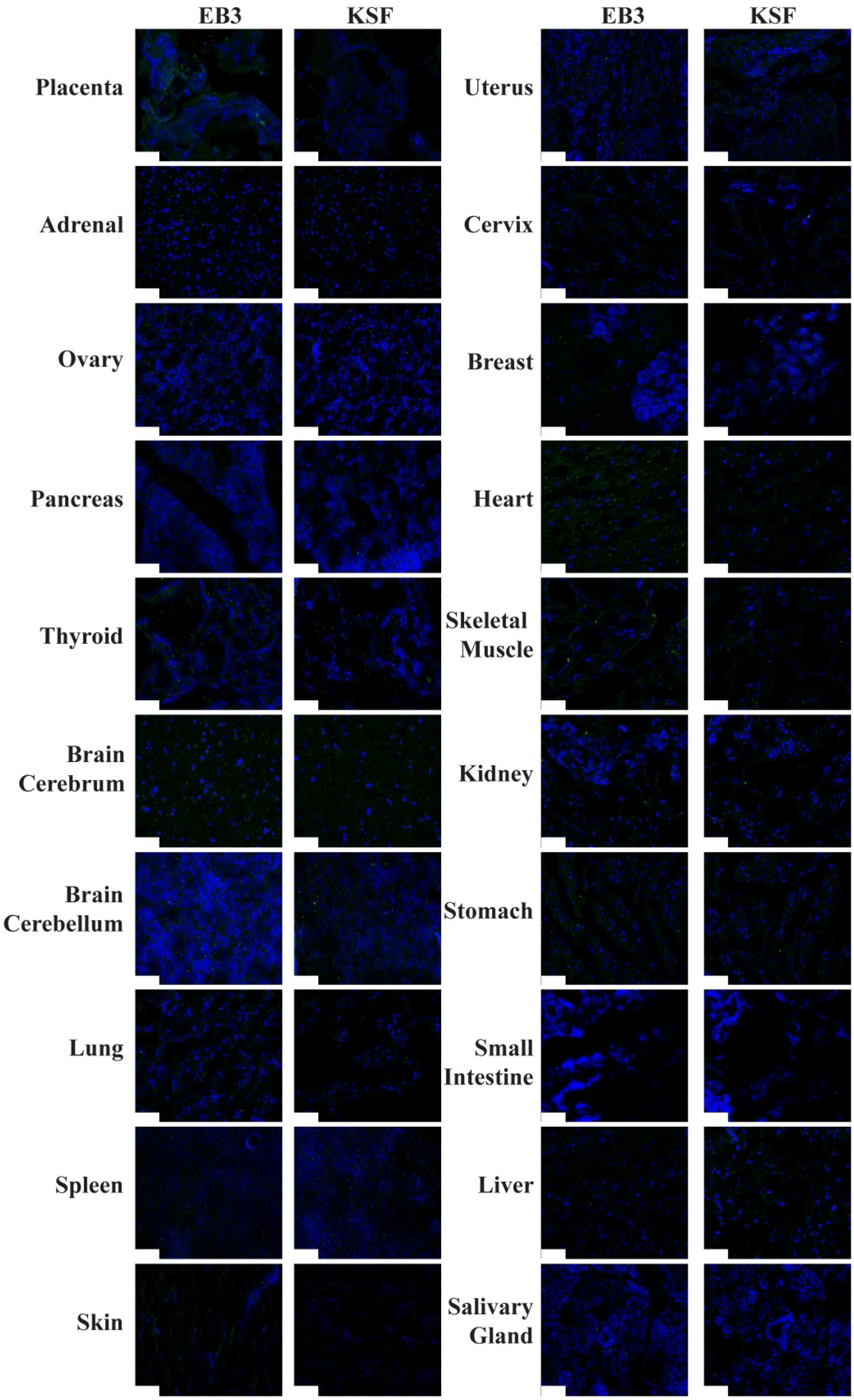
2.3. Immunofluorescence Findings
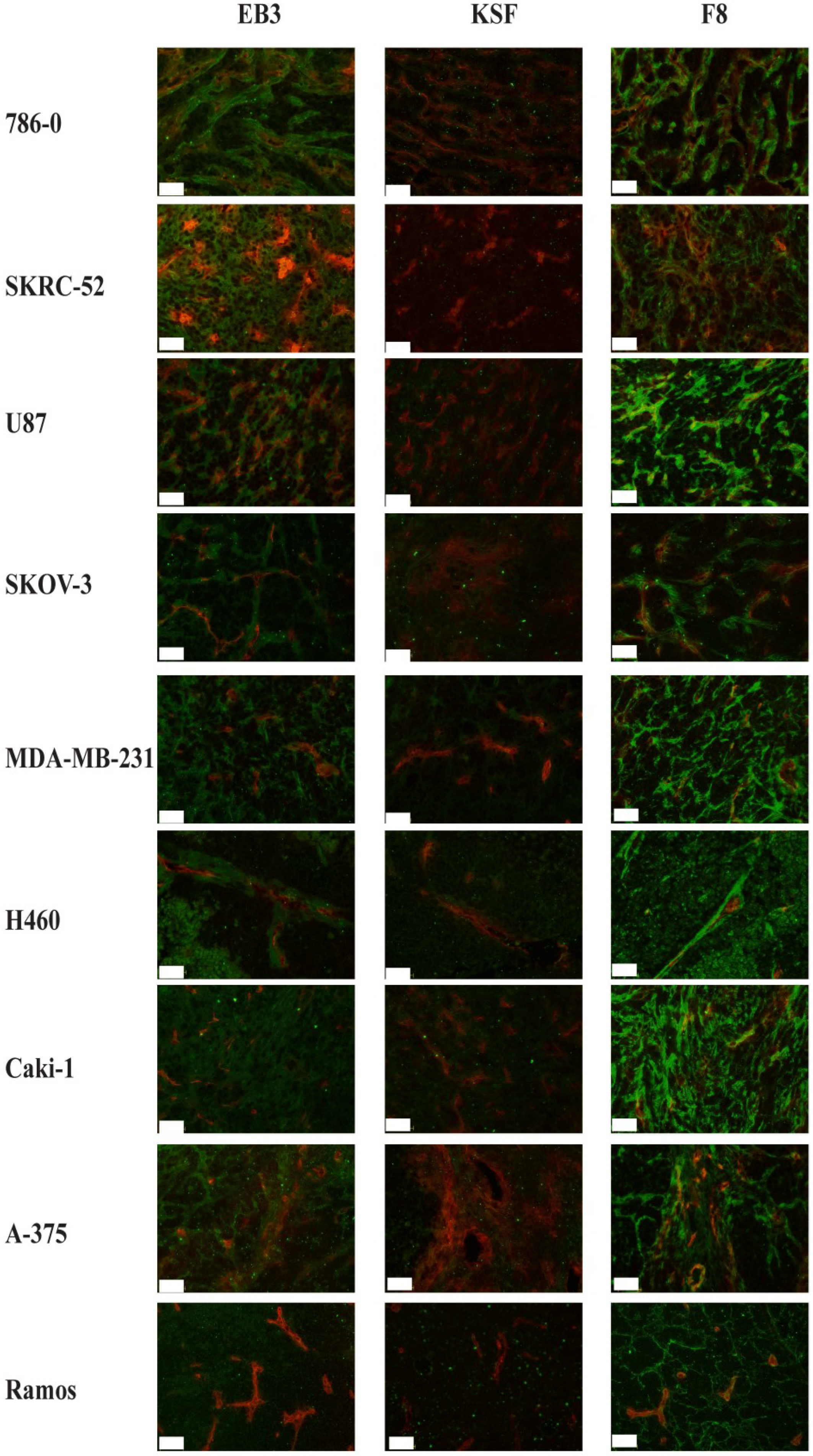
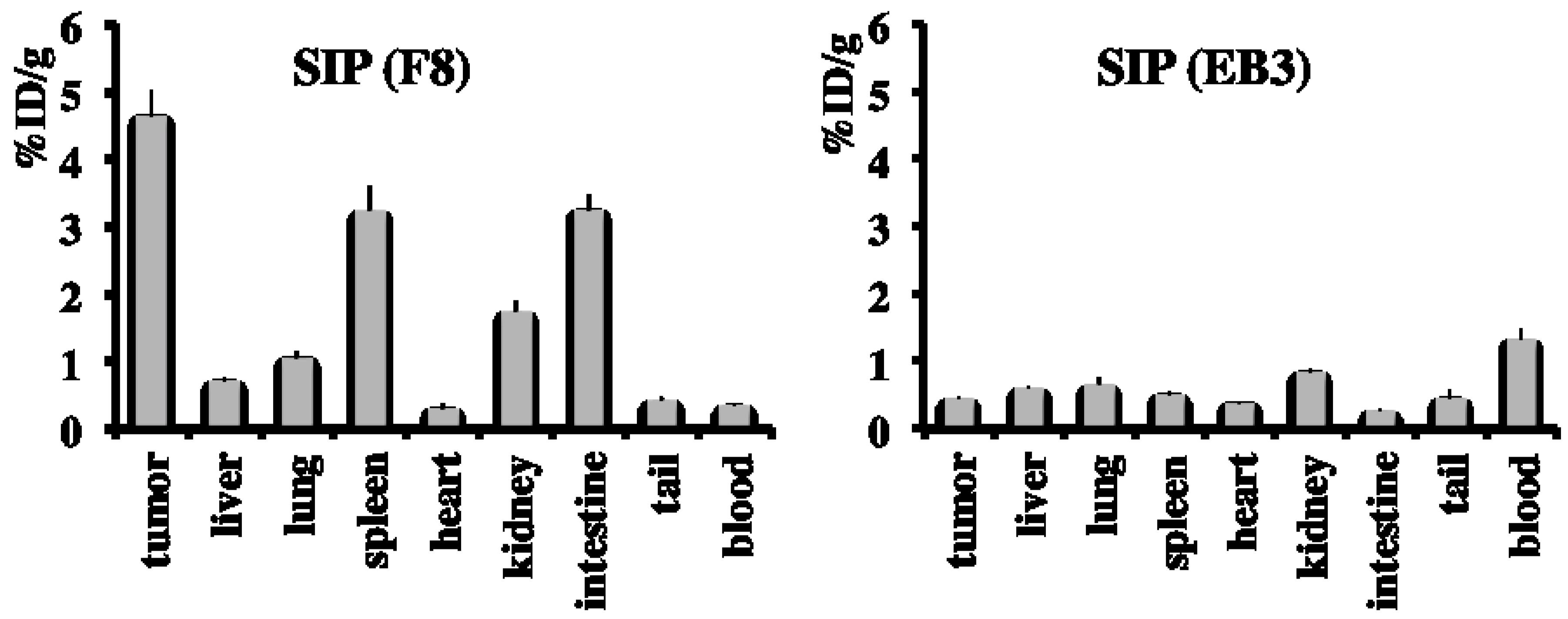
2.4. In Vivo Biodistribution Study
3. Experimental Section
3.1. Cloning and Expression of Human DLK1-Fc Fusion Protein
3.2. Mass Spectrometric Analysis of Human DLK1-Fc Fusion Protein
3.3. Screening of Phage Display Libraries and Selection of the Antibody
3.4. Characterization of scFv Antibody Fragments
3.5. BIAcore Analysis
3.6. Sequencing of scFv Genes
3.7. Cloning and Expression of Small Immune Proteins (SIP)
3.8. Immunofluorescence on Frozen Tissue Sections
3.9. Biodistribution Studies
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Aggarwal, S. What's fueling the biotech engine—2011 to 2012. Nat. Biotech. 2012, 30, 1191–1197. [Google Scholar] [CrossRef]
- Walsh, G. Biopharmaceutical benchmarks 2014. Nat. Biotech. 2014, 32, 992–1000. [Google Scholar] [CrossRef]
- Shim, H. One target, different effects: A comparison of distinct therapeutic antibodies against the same targets. Exp. Mol. Med. 2011, 43, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sul, H.S. Ectodomain Shedding of Preadipocyte Factor 1 (Pref-1) by Tumor Necrosis Factor Alpha Converting Enzyme (TACE) and Inhibition of Adipocyte Differentiation. Mol. Cell. Biol. 2006, 26, 5421–5435. [Google Scholar] [CrossRef] [PubMed]
- Laborda, J.; Sausville, E.A.; Hoffman, T.; Notario, V. dlk, a putative mammalian homeotic gene differentially expressed in small cell lung carcinoma and neuroendocrine tumor cell line. J. Biol. Chem. 1993, 268, 3817–3820. [Google Scholar] [PubMed]
- Baladrón, V.; Ruiz-Hidalgo, M.J.; Nueda, M.L.; Díaz-Guerra, M.J.M.; García-Ramírez, J.J.; Bonvini, E.; Gubina, E.; Labord, J. dlk acts as a negative regulator of Notch1 activation through interactions with specific EGF-like repeats. Exp. Cell Res. 2005, 303, 343–359. [Google Scholar] [CrossRef] [PubMed]
- Nueda, M.-L.; Baladrón, V.; Sánchez-Solana, B.; Ballesteros, M.-Á.; Laborda, J. The EGF-like Protein dlk1 Inhibits Notch Signaling and Potentiates Adipogenesis of Mesenchymal Cells. J. Mol. Biol. 2007, 367, 1281–1293. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Forman, S.J.; Bhatia, R. Expression of DLK1 in hematopoietic cells results in inhibition of differentiation and proliferation. Oncogene 2005, 24, 4472–4476. [Google Scholar] [CrossRef] [PubMed]
- Smas, C.M.; Sul, H.S. Pref-1, a protein containing EGF-like repeats, inhibits adipocyte differentiation. Cell 1993, 73, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Andersen, D.C.; Petersson, S.J.; Jørgensen, L.H.; Bollen, P.; Jensen, P.B.; Teisner, B.; Schroeder, H.D.; Jensen, C.H. Characterization of DLK1+ Cells Emerging During Skeletal Muscle Remodeling in Response to Myositis, Myopathies, and Acute Injury. Stem Cells 2009, 27, 898–908. [Google Scholar] [CrossRef] [PubMed]
- Tanimizu, N.; Nishikawa, M.; Saito, H.; Tsujimura, T.; Miyajima, A. Isolation of hepatoblasts based on the expression of dlk/pref-1. Journal of Cell Science 2003, 116, 1775–1786. [Google Scholar] [CrossRef] [PubMed]
- Yevtodiyenko, A.; Schmidt, J.V. Dlk1 expression marks developing endothelium and sites of branching morphogenesis in the mouse embryo and placenta. Dev. Dynam. 2006, 235, 1115–1123. [Google Scholar] [CrossRef]
- Floridon, C.; Jensen, C.H.; Thorsen, P.; Nielsen, O.; Sunde, L.; Westergaard, J.G.; Thomsen, S.G.; Teisner, B. Does Fetal antigen 1 (FA1) identify cells with regenerative, endocrine and neuroendocrine potentials? A study of FA1 in embryonic, fetal, and placental tissue and in maternal circulation. Differentiation 2000, 66, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Jensen, C.H.; Krogh, T.N.; Højrup, P.; Clausen, P.P.; Skjødt, K.; Larsson, L.-I.; Enghild, J.J.; Teisner, B. Protein Structure of Fetal Antigen 1 (FA1). Eur. J. Biochem. 1994, 225, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Jensen, C.H.; Teisner, B.; Højrup, P.; Rasmussen, H.B.; Madsen, O.D.; Nielsen, B.; Skjødt, K. Studies on the isolation, structural analysis and tissue localization of fetal antigen 1 and its relation to a human adrenal-specific cDNA, pG2. Hum. Reprod. 1993, 8, 635–641. [Google Scholar] [PubMed]
- Van Limpt, V.; Chan, A.; Caron, H.; Van Sluis, P.; Boon, K.; Hermus, M.-C.; Versteeg, R. SAGE analysis of neuroblastoma reveals a high expression of the human homologue of the Drosophila Delta gene. Med. Pediatr. Oncol. 2000, 35, 554–558. [Google Scholar] [CrossRef] [PubMed]
- Harel, A.; Dalah, I.; Pietrokovski, S.; Safran, M.; Lancet, D. Omics data management and annotation. In Bioinformatics for Omics Data; Mayer, B., Ed.; Humana Press: New York, NY, USA, 2011; Volume 719, pp. 71–96. [Google Scholar]
- Traustadottir, G.A.; Kosmina, R.; Sheikh, S.P.; Jensenab, C.H.; Andersen, D.C. Preadipocytes proliferate and differentiate under the guidance of Delta-like 1 homolog (DLK1). Adipocyte 2013, 2, 272–275. [Google Scholar] [CrossRef] [PubMed]
- Surmacz, B.; Noisa, P.; Risner-Janiczek, J.R.; Hui, K.; Ungless, M.; Cui, W.; Li, M. DLK1 promotes neurogenesis of human and mouse pluripotent stem cell-derived neural progenitors via modulating Notch and BMP signalling. Stem Cell Rev. 2012, 8, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.S.; Smas, C.M.; Lee, K.; Villena, J.A.; Kim, K.-H.; Yun, E.J.; Sul, H.S. Mice Lacking Paternally Expressed Pref-1/Dlk1 Display Growth Retardation and Accelerated Adiposity. Mol. Cell. Biol. 2002, 22, 5585–5592. [Google Scholar] [CrossRef]
- Waddell, J.N.; Zhang, P.; Wen, Y.; Gupta, S.K.; Yevtodiyenko, A.; Schmidt, J.V.; Bidwell, C.A.; Kumar, A.; Kuang, S. Dlk1 Is Necessary for Proper Skeletal Muscle Development and Regeneration. PLoS One 2010, 5, e15055. [Google Scholar] [CrossRef] [PubMed]
- Raghunandan, R.; Ruiz-Hidalgo, M.; Jia, Y.; Ettinger, R.; Rudikoff, E.; Riggins, P.; Farnsworth, R.; Tesfaye, A.; Laborda, J.; Bauer, S.R. Dlk1 influences differentiation and function of B lymphocytes. Stem Cells Dev. 2008, 17, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhang, X.; Zhang, M.; Zhu, J.-D.; Zhang, Y.-L.; Lin, Y.; Wang, K.-S.; Qi, X.-F.; Zhang, Q.; Liu, G.-Z.; et al. Up-regulation of DLK1 as an imprinted gene could contribute to human hepatocellular carcinoma. Carcinogenesis 2007, 28, 1094–1103. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.-H.; Yang, R.-J.; Dong, B.; Xing, B. Progenitor gene DLK1 might be an independent prognostic factor of liver cancer. Exp. Opin. Biol. Ther. 2008, 8, 371–377. [Google Scholar] [CrossRef]
- Chi Sabins, N.; Taylor, J.L.; Fabian, K.P.L.; Appleman, L.J.; Maranchie, J.K.; Stolz, D.B.; Storkus, W.J. DLK1: A Novel Target for Immunotherapeutic Remodeling of the Tumor Blood Vasculature. Mol. Ther. 2013, 21, 1958–1968. [Google Scholar] [CrossRef] [PubMed]
- Van Limpt, V.E.; Chan, A.J.; Van Sluis, P.G.; Caron, H.N.; van Noesel, C.J.M.; Versteeg, R. High delta-like 1 expression in a subset of neuroblastoma cell lines corresponds to a differentiated chromaffin cell type. Int. J. Cancer 2003, 105, 61–69. [Google Scholar] [CrossRef] [PubMed]
- López-Terrada, D.; Gunaratne, P.H.; Adesina, A.M.; Pulliam, J.; Hoang, D.M.; Nguyen, Y.; Mistretta, T.-A.; Margolin, J.; Finegold, M.J. Histologic subtypes of hepatoblastoma are characterized by differential canonical Wnt and Notch pathway activation in DLK+ precursors. Hum. Pathol. 2009, 40, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Chen, Z.; Liu, D.; Cen, J.; Gu, M. Expression of Dlk1 gene in myelodysplastic syndrome determined by microarray, and its effects on leukemia cells. Int. J. Mol. Med. 2008, 22, 61–68. [Google Scholar] [PubMed]
- Sakajiri, S.; O'kelly, J.; Yin, D.; Miller, C.W.; Hofmann, W.K.; Oshimi, K.; Shih, L.-Y.; Kim, K.-H.; Sul, H.S.; Jensen, C.H.; et al. Dlk1 in normal and abnormal hematopoiesis. Leukemia 2005, 19, 1404–1410. [Google Scholar] [CrossRef] [PubMed]
- Altenberger, T.; Bilban, M.; Auer, M.; Knospc, E.; Wolfsbergerc, S.; Gartnera, W.; Minevaa, I.; Zielinskid, C.; Wagnera, L.; Lugera, A. Identification of DLK1 variants in pituitary- and neuroendocrine tumors. Biochem. Biophys. Res. Commun. 2006, 340, 995–1005. [Google Scholar] [CrossRef] [PubMed]
- Yanai, H.; Nakamura, K.; Hijioka, S.; Kamei, A.; Ikari, T.; Ishikawa, Y.; Shinozaki, E.; Mizunuma, N.; Hatake, K.; Miyajima, A. Dlk-1, a cell surface antigen on foetal hepatic stem/progenitor cells, is expressed in hepatocellular, colon, pancreas and breast carcinomas at a high frequency. J. Biochem. 2010, 148, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.W.; Jo, K.; Lee, D.; Yu, J.; Park, J.H.; Park, C.-W.; Kim, E.J.; Park, Y.J. Method for inhibiting cancer metastasis by administration of the extracellular domain DLK1 or a DLK1-FC fusion protein. US 8785392 B2, 22 July 2014. [Google Scholar]
- Yu, F.; Hao, X.; Zhao, H.; Ge, C.; Yao, M.; Yang, S.; Li, J. Delta-like 1 contributes to cell growth by increasing the interferon-inducible protein 16 expression in hepatocellular carcinoma. Liver Int. 2010, 30, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Lin, Q.; Zelterman, D.; Yun, Z. Hypoxia-regulated delta-like 1 homologue enhances cancer cell stemness and tumorigenicity. Cancer Res. 2009, 69, 9271–9280. [Google Scholar] [CrossRef] [PubMed]
- Orr, B.; Grace, O.C.; Brown, P.; Riddick, A.C.P.; Stewart, G.D.; Franco, O.E.; Hayward, S.W.; Thomson, A.A. Reduction of pro-tumorigenic activity of human prostate cancer-associated fibroblasts using Dlk1 or SCUBE1. Dis. Model. Mech. 2013, 6, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, T.; Chano, T.; Minami, K.; Okabe, H.; Okada, Y.; Okamoto, K. Imprinted DLK1 is a putative tumor suppressor gene and inactivated by epimutation at the region upstream of GTL2 in human renal cell carcinoma. Hum. Mol. Genet. 2006, 15, 821–830. [Google Scholar] [CrossRef] [PubMed]
- Nueda, M.L.; Naranjo, A.I.; Baladron, V.; Labordab, J. The proteins DLK1 and DLK2 modulate NOTCH1-dependent proliferation and oncogenic potential of human SK-MEL-2 melanoma cells. Biochim. Biophys. Acta 2014, 1843, 2674–2684. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Oksvold, P.; Fagerberg, L.; Lundberg, E.; Jonasson, K.; Forsberg, M.; Zwahlen, M.; Kampf, C.; Wester, K.; Hober, S.; et al. Towards a knowledge-based Human Protein Atlas. Nat Biotech. 2010, 28, 1248–1250. [Google Scholar] [CrossRef]
- Villa, A.; Trachsel, E.; Kaspar, M.; Schliemann, C.; Sommavilla, R.; Rybak, J.-N.; Rösli, C.; Borsi, L.; Neri, D. A high-affinity human monoclonal antibody specific to the alternatively spliced EDA domain of fibronectin efficiently targets tumor neo-vasculature in vivo. Int. J. Cancer 2008, 122, 2405–2413. [Google Scholar] [CrossRef] [PubMed]
- Schwager, K.; Villa, A.; Rosli, C.; Neri, D.; Rösli-Khabas, M.; Moser, G. A comparative immunofluorescence analysis of three clinical-stage antibodies in head and neck cancer. Head Neck Oncol. 2011, 3, 25. [Google Scholar] [CrossRef] [PubMed]
- Schliemann, C.; Neri, D. Antibody-based targeting of the tumor vasculature. Biochim. Biophys. Acta 2007, 1776, 175–192. [Google Scholar] [PubMed]
- Weber, M.; Bujak, E.; Putelli, A.; Villa, A.; Matasci, M.; Gualandi, L.; Hemmerle, T.; Wulhfard, S.; Neri, D. A highly functional synthetic phage display library containing over 40 billion human antibody clones. PLoS One 2014, 9, e100000. [Google Scholar] [CrossRef] [PubMed]
- Huston, J.S.; Levinson, D.; Mudgett-Hunter, M.; Tai, M.S.; Novotný, J.; Margolies, M.N.; Ridge, R.J.; Bruccoleri, R.E.; Haber, E.; Crea, R. Protein engineering of antibody binding sites: recovery of specific activity in an anti-digoxin single-chain Fv analogue produced in Escherichia coli. Proc. Natl. Acad. Sci. USA 1988, 85, 5879–5883. [Google Scholar] [CrossRef] [PubMed]
- Silacci, M.; Brack, S.; Schirru, G.; Mårlind, J.; Ettorre, A.; Merlo, A.; Viti, F.; Neri, D. Design, construction, and characterization of a large synthetic human antibody phage display library. Proteomics 2005, 5, 2340–2350. [Google Scholar] [CrossRef] [PubMed]
- Borsi, L.; Balza, E.; Bestagno, M.; Castellani, P.; Carnemolla, B.; Biro, A.; Leprini, A.; Sepulveda, J.; Burrone, O.; Neri, D.; et al. Selective targeting of tumoral vasculature: comparison of different formats of an antibody (L19) to the ED-B domain of fibronectin. Int. J. Cancer 2002, 102, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Frey, K.; Zivanovic, A.; Schwager, K.; Neri, D. Antibody-based targeting of interferon-alpha to the tumor neovasculature: A critical evaluation. Integr. Biol. 2011, 3, 468–478. [Google Scholar] [CrossRef]
- Schliemann, C.; Neri, D. Antibody-based vascular tumor targeting. Recent Results Cancer Res. 2010, 180, 201–216. [Google Scholar] [PubMed]
- Pfaffen, S.; Hemmerle, T.; Weber, M.; Neri, D. Isolation and characterization of human monoclonal antibodies specific to MMP-1A, MMP-2 and MMP-3. Exp. Cell Res. 2010, 316, 836–847. [Google Scholar] [CrossRef] [PubMed]
- Deiuliis, J.A.; Li, B.; Lyvers-Peffer, P.A.; Moellera, S.J.; Lee, K. Alternative splicing of delta-like 1 homolog (DLK1) in the pig and human. Comp. Biochem. Physi. B 2006, 145, 50–59. [Google Scholar] [CrossRef]
- Bujak, E.; Pretto, F.; Ritz, D.; Gualandi, L.; Wulhfard, S.; Neri, D. Monoclonal antibodies to murine thrombospondin-1 and thrombospondin-2 reveal differential expression patterns in cancer and low antigen expression in normal tissues. Exp. Cell Res. 2014, 327, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Anderson, L.; Seilhamer, J. A comparison of selected mRNA and protein abundances in human liver. Electrophoresis 1997, 18, 533–537. [Google Scholar] [CrossRef] [PubMed]
- Gygi, S.P.; Rochon, Y.; Franza, B.R.; Aebersold, R. Correlation between protein and mRNA abundance in yeast. Mol. Cell. Biol. 1999, 19, 1720–1730. [Google Scholar] [PubMed]
- Pini, A.; Viti, F.; Santucci, A.; Carnemoll, B.; Zardi, L.; Neri, P.; Neri, D. Design and Use of a Phage Display Library: Human antibodies with subnanomolar affinity against a marker of angiogenesis eluted from a two-dimensional gel. J. Biol. Chem. 1998, 273, 21769–21776. [Google Scholar] [CrossRef] [PubMed]
- Brack, S.S.; Silacci, M.; Birchler, M.; Neri, D. Tumor-Targeting Properties of Novel Antibodies Specific to the Large Isoform of Tenascin-C. Clin. Cancer Res. 2006, 12, 3200–3208. [Google Scholar] [CrossRef] [PubMed]
- Bujak, E.; Matasci, M.; Neri, D.; Wulhfard, S. Reformatting of scfv antibodies into the scfv-fc format and their downstream purification. In Monoclonal Antibodies; Ossipow, V., Fischer, N., Eds.; Humana Press: New York, NY, USA, 2014; Volume 1131, pp. 315–334. [Google Scholar]
- Silacci, M.; Brack, S.; Spath, N.; Buck, A.; Hillinger, S.; Arni, S.; Weder, W.; Zardi, L.; Neri, D. Human monoclonal antibodies to domain C of tenascin-C selectively target solid tumors in vivo. Protein Eng. Des. Sel. 2006, 19, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Zuberbühler, K.; Palumbo, A.; Bacci, C.; Giovannoni, L.; Sommavilla, R.; Kaspar, M.; Trachsel, E.; Neri, D. A general method for the selection of high-level scFv and IgG antibody expression by stably transfected mammalian cells. Protein Eng. Des. Sel. 2009, 22, 169–174. [Google Scholar] [CrossRef] [PubMed]
- List, T.; Neri, D. Biodistribution studies with tumor-targeting bispecific antibodies reveal selective accumulation at the tumor site. mAbs 2012, 4, 775–783. [Google Scholar] [CrossRef] [PubMed]
- Tarli, L.; Balza, E.; Viti, F.; Borsi, L.; Castellani, P.; Berndorff, D.; Dinkelborg, L.; Neri, D.; Zardi, L. A High-Affinity Human Antibody That Targets Tumoral Blood Vessels. Blood 1999, 94, 192–198. [Google Scholar] [PubMed]
- Fraker, P.J. Reprint of “Protein and Cell Membrane Iodinations with a Sparingly Soluble Chloroamide, 1,3,4,6-Tetrachloro-3a,6a-Diphenylglycoluril”. Biochem. Biophys. Res. Commun. 2012, 425, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Salacinski, P.R.P.; Mclean, C.; Sykes, J.E.C.; Clement-Jones, V.V.; Lowry, P.J. Iodination of proteins, glycoproteins, and peptides using a solid-phase oxidizing agent, 1,3,4,6-tetrachloro-3α,6α-diphenyl glycoluril (Iodogen). Anal. Biochem. 1981, 117, 136–146. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bujak, E.; Ritz, D.; Neri, D. A Monoclonal Antibody to Human DLK1 Reveals Differential Expression in Cancer and Absence in Healthy Tissues. Antibodies 2015, 4, 71-87. https://doi.org/10.3390/antib4020071
Bujak E, Ritz D, Neri D. A Monoclonal Antibody to Human DLK1 Reveals Differential Expression in Cancer and Absence in Healthy Tissues. Antibodies. 2015; 4(2):71-87. https://doi.org/10.3390/antib4020071
Chicago/Turabian StyleBujak, Emil, Danilo Ritz, and Dario Neri. 2015. "A Monoclonal Antibody to Human DLK1 Reveals Differential Expression in Cancer and Absence in Healthy Tissues" Antibodies 4, no. 2: 71-87. https://doi.org/10.3390/antib4020071
APA StyleBujak, E., Ritz, D., & Neri, D. (2015). A Monoclonal Antibody to Human DLK1 Reveals Differential Expression in Cancer and Absence in Healthy Tissues. Antibodies, 4(2), 71-87. https://doi.org/10.3390/antib4020071