
A Beautiful Bird in the Neighborhood: Canopy Cover and Vegetation Structure Predict Avian Presence in High-Vacancy City



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Vacant Lot Classification
2.3. Vegetation Structure of 1 Ha Plots
2.4. Bird Counts
2.5. Data Analysis: Vegetation
2.6. Data Analysis: Birds
2.7. Data Analysis: Predicting Species Abundance
3. Results
3.1. Vacant Lot Characteristics
3.2. One-Ha Study Plots
3.3. Birds
3.4. Principal Components Analysis (PCA) of Vacant Lot Characteristics
3.5. Detection and Abundance Models for the Five Most Abundant Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| GCC | Green City Coalition |
| NAIP | National Agriculture Imagery Program |
| PCA | Principal component analysis |
References
- Anderson, E.C.; Minor, E.S. Vacant Lots: An Underexplored Resource for Ecological and Social Benefits in Cities. Urban For. Urban Green. 2017, 21, 146–152. [Google Scholar] [CrossRef]
- Riley, C.B.; Perry, K.I.; Ard, K.; Gardiner, M.M. Asset or Liability? Ecological and Sociological Tradeoffs of Urban Spontaneous Vegetation on Vacant Land in Shrinking Cities. Sustainability 2018, 10, 2139. [Google Scholar] [CrossRef]
- Schell, C.J.; Dyson, K.; Fuentes, T.L.; Des Roches, S.; Harris, N.C.; Miller, D.S.; Woelfle-Erskine, C.A.; Lambert, M.R. The Ecological and Evolutionary Consequences of Systemic Racism in Urban Environments. Science 2020, 369, eaay4497. [Google Scholar] [CrossRef]
- Melles, S. Urban Bird Diversity as an Indicator of Human Social Diversity and Economic Inequality in Vancouver, British Columbia. Urban Habitats 2005, 3, 25–48. [Google Scholar]
- Kong, F.; Yin, H.; Nakagoshi, N.; Zong, Y. Urban Green Space Network Development for Biodiversity Conservation: Identification Based on Graph Theory and Gravity Modeling. Landsc. Urban Plan. 2010, 95, 16–27. [Google Scholar] [CrossRef]
- Lerman, S.B.; Warren, P.S. The Conservation Value of Residential Yards: Linking Birds and People. Ecol. Appl. 2011, 21, 1327–1339. [Google Scholar]
- Aronson, M.F.J.; La Sorte, F.A.; Nilon, C.H.; Katti, M.; Goddard, M.A.; Lepczyk, C.A.; Warren, P.S.; Williams, N.S.G.; Cilliers, S.; Clarkson, B.; et al. A Global Analysis of the Impacts of Urbanization on Bird and Plant Diversity Reveals Key Anthropogenic Drivers. Proc. R. Soc. B Biol. Sci. 2014, 281, 20133330. [Google Scholar] [CrossRef]
- Lepczyk, C.A.; Aronson, M.F.J.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; MacIvor, J.S. Biodiversity in the City: Fundamental Questions for Understanding the Ecology of Urban Green Spaces for Biodiversity Conservation. BioScience 2017, 67, 799–807. [Google Scholar] [CrossRef]
- Aronson, M.F.; Lepczyk, C.A.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; MacIvor, J.S.; Nilon, C.H.; Vargo, T. Biodiversity in the City: Key Challenges for Urban Green Space Management. Front. Ecol. Environ. 2017, 15, 189–196. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization as a Major Cause of Biotic Homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- Chamberlain, D.E.; Cannon, A.R.; Toms, M.P.; Leech, D.I.; Hatchwell, B.J.; Gaston, K.J. Avian Productivity in Urban Landscapes: A Review and Meta-Analysis. IBIS 2009, 151, 1–18. [Google Scholar] [CrossRef]
- Rega-Brodsky, C.C.; Nilon, C.H. Forest Cover Is Important across Multiple Scales for Bird Communities in Vacant Lots. Urban Ecosyst. 2017, 20, 561–571. [Google Scholar] [CrossRef]
- Burkholder, S. The New Ecology of Vacancy: Rethinking Land Use in Shrinking Cities. Sustainability 2012, 4, 1154–1172. [Google Scholar] [CrossRef]
- Cornelis, J.; Hermy, M. Biodiversity Relationships in Urban and Suburban Parks in Flanders. Landsc. Urban Plan. 2004, 69, 385–401. [Google Scholar]
- Fuller, R.A.; Irvine, K.N.; Devine-Wright, P.; Warren, P.H.; Gaston, K.J. Psychological Benefits of Greenspace Increase with Biodiversity. Biol. Lett. 2007, 3, 390–394. [Google Scholar] [CrossRef]
- Loram, A.; Warren, P.H.; Gaston, K.J. Urban Domestic Gardens (XIV): The Characteristics of Gardens in Five Cities. Environ. Manag. 2008, 42, 361–376. [Google Scholar] [CrossRef]
- Lerman, S.B.; Warren, P.S.; Gan, H.; Shochat, E. Linking Foraging Decisions to Residential Yard Bird Composition. PLoS ONE 2012, 7, e43497. [Google Scholar]
- Goddard, M.A.; Dougill, A.J.; Benton, T.G. Why Garden for Wildlife? Social and Ecological Drivers, Motivations and Barriers for Biodiversity Management in Residential Landscapes. Ecol. Econ. 2013, 86, 258–273. [Google Scholar] [CrossRef]
- Eakin, C.J.; Campa, H.; Linden, D.W.; Roloff, G.J.; Bradley Rowe, D.; Westphal, J. Avian Response to Green Roofs in Urban Landscapes in the Midwestern USA. Wildl. Soc. Bull. 2015, 39, 574–582. [Google Scholar] [CrossRef]
- Kowarik, I.; Buchholz, S.; von der Lippe, M.; Seitz, B. Biodiversity Functions of Urban Cemeteries: Evidence from One of the Largest Jewish Cemeteries in Europe. Urban For. Urban Green. 2016, 19, 68–78. [Google Scholar] [CrossRef]
- Rupprecht, C.D.D.; Byrne, J.A. Informal Urban Greenspace: A Typology and Trilingual Systematic Review of Its Role for Urban Residents and Trends in the Literature. Urban For. Urban Green. 2014, 13, 597–611. [Google Scholar] [CrossRef]
- Rega-Brodsky, C.; Nilon, C.; Warren, P. Avian Abundance Patterns in Relation to the Distribution of Small Urban Greenspaces. J. Urban Plan. Dev. 2015, 141, A4015002. [Google Scholar] [CrossRef]
- Greenberg, M.; Popper, F.; Schneider, D.; West, B. Community Organizing to Prevent TOADS in the United States. Community Dev. J. 1993, 28, 55–65. [Google Scholar] [CrossRef]
- Davis, A.Y.; Belaire, J.A.; Farfan, M.A.; Milz, D.; Sweeney, E.R.; Loss, S.R.; Minor, E.S. Green Infrastructure and Bird Diversity across an Urban Socioeconomic Gradient. Ecosphere 2012, 3, art105. [Google Scholar] [CrossRef]
- Németh, J.; Langhorst, J. Rethinking Urban Transformation: Temporary Uses for Vacant Land. Cities 2014, 40, 143–150. [Google Scholar] [CrossRef]
- Nassauer, J.I.; Raskin, J. Urban Vacancy and Land Use Legacies: A Frontier for Urban Ecological Research, Design, and Planning. Landsc. Urban Plan. 2014, 125, 245–253. [Google Scholar] [CrossRef]
- Tighe, J.R.; Ganning, J.P. The Divergent City: Unequal and Uneven Development in St. Louis. Urban Geogr. 2015, 36, 654–673. [Google Scholar] [CrossRef]
- Rothstein, R. The Color of Law: A Forgotten History of How Our Government Segregated America; Liveright Publishing: New York, NY, USA, 2017. [Google Scholar]
- Wilhelm Stanis, S.; Piontek, E.; Xu, S.; Mallinak, A.; Nilon, C.; Hall, D.M. Residents’ Perceptions of Urban Greenspace in a Shrinking City: Ecosystem Services and Environmental Justice. Land 2024, 13, 1554. [Google Scholar] [CrossRef]
- Cooperman, J. The Story of Segregation in St. Louis; St. Louis Magazine: St. Louis, MO, USA, 2014. [Google Scholar]
- Azerrad, J.; Nilon, C. Avian Community Characteristics of Urban Greenspaces in St. Louis, Missouri. In Avian Ecology and Conservation in an Urbanizing World; Springer: Boston, MA, USA, 2001; pp. 487–503. [Google Scholar]
- Oliver, A.J.; Hong-Wa, C.; Devonshire, J.; Olea, K.R.; Rivas, G.F.; Gahl, M.K. Avifauna Richness Enhanced in Large, Isolated Urban Parks. Landsc. Urban Plan. 2011, 102, 215–225. [Google Scholar] [CrossRef]
- Nilon, C.H.; Aronson, M.F.J.; Cilliers, S.S.; Dobbs, C.; Frazee, L.J.; Goddard, M.A.; O’Neill, K.M.; Roberts, D.; Stander, E.K.; Werner, P.; et al. Planning for the Future of Urban Biodiversity: A Global Review of City-Scale Initiatives. BioScience 2017, 67, 332–342. [Google Scholar] [CrossRef]
- Goddard, M.A.; Dougill, A.J.; Benton, T.G. Scaling up from Gardens: Biodiversity Conservation in Urban Environments. Trends Ecol. Evol. 2010, 25, 90–98. [Google Scholar] [CrossRef]
- Lerman, S.B.; Narango, D.L.; Avolio, M.L.; Bratt, A.R.; Engebretson, J.M.; Groffman, P.M.; Hall, S.J.; Heffernan, J.B.; Hobbie, S.E.; Larson, K.L.; et al. Residential Yard Management and Landscape Cover Affect Urban Bird Community Diversity across the Continental USA. Ecol. Appl. 2021, 31, e02455. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Miller, P.A.; Nowak, D.J. Urban Vacant Land Typology: A Tool for Managing Urban Vacant Land. Sustain. Cities Soc. 2018, 36, 144–156. [Google Scholar] [CrossRef]
- Haase, D.; Frantzeskaki, N.; Elmqvist, T. Ecosystem Services in Urban Landscapes: Practical Applications and Governance Implications. Ambio 2014, 43, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Kuras, E.R.; Warren, P.S.; Zinda, J.A.; Aronson, M.F.J.; Cilliers, S.; Goddard, M.A.; Nilon, C.H.; Winkler, R. Urban Socioeconomic Inequality and Biodiversity Often Converge, but Not Always: A Global Meta-Analysis. Landsc. Urban Plan. 2020, 198, 103799. [Google Scholar] [CrossRef]
- Green City Coalition. Green City Coalition—Vacancy to Vibrancy. Available online: https://www.greencitycoalition.org/ (accessed on 24 April 2025).
- Andersson, E.; Barthel, S.; Borgström, S.; Colding, J.; Elmqvist, T.; Folke, C.; Gren, Å. Reconnecting Cities to the Biosphere: Stewardship of Green Infrastructure and Urban Ecosystem Services. AMBIO 2014, 43, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Ellis-Soto, D.; Chapman, M.; Locke, D.H. Historical Redlining Is Associated with Increasing Geographical Disparities in Bird Biodiversity Sampling in the United States. Nat. Hum. Behav. 2023, 7, 1869–1877. [Google Scholar] [CrossRef]
- Wood, E.M.; Esaian, S.; Benitez, C.; Ethington, P.J.; Longcore, T.; Pomara, L.Y. Historical Racial Redlining and Contemporary Patterns of Income Inequality Negatively Affect Birds, Their Habitat, and People in Los Angeles, California. Ornithol. Appl. 2024, 126, duad044. [Google Scholar] [CrossRef]
- Gordon, C. St. Louis Blues: The Urban Crisis in the Gateway City. St. Louis Univ. Public Law Rev. 2013, 33, 81. [Google Scholar]
- Beauregard, R. Urban Population Loss in Historical Perspective: United States, 1820–2000. Environ. Plan. A 2009, 41, 514–528. [Google Scholar] [CrossRef]
- Vey, J. Restoring Prosperity: The State Role in Revitalizing America’s Older Industrial Cities; Brookings Institution: Washington, DC, USA, 2011. [Google Scholar]
- Thom, R.H.; Wilson, J.H. The Natural Divisions of Missouri. Nat. Areas J. 1983, 3, 44–51. [Google Scholar]
- Wayman, N. History of St. Louis Neighborhoods; St. Louis Community Development Agency: Saint Louis, MO, USA, 1979. [Google Scholar]
- Nelson, P.W. The Terrestrial Natural Communities of Missouri; Missouri Natural Areas Committee: Jefferson City, MO, USA, 2010. [Google Scholar]
- Rogers, G.F.; Rowntree, R.A. Intensive Surveys of Structure and Change in Urban Natural Areas. Landsc. Urban Plan. 1988, 15, 59–78. [Google Scholar] [CrossRef]
- DeGraaf, R.; Geis, A.; Healy, P. Bird Population and Habitat Surveys in Urban Areas. Landsc. Urban Plan. 1991, 21, 181–188. [Google Scholar] [CrossRef]
- Caratti, J. Line Intercept (LI) Sampling Method. Gen. Tech. Rep. (GTR) 2006, 56, 1–14. [Google Scholar]
- Sung, C.Y.; Li, M.-H.; Rogers, G.O.; Volder, A.; Wang, Z. Investigating Alien Plant Invasion in Urban Riparian Forests in a Hot and Semi-Arid Region. Landsc. Urban Plan. 2011, 100, 278–286. [Google Scholar] [CrossRef]
- Fernández-Juricic, E. Can Human Disturbance Promote Nestedness? A Case Study with Breeding Birds in Urban Habitat Fragments. Oecologia 2002, 131, 269–278. [Google Scholar] [CrossRef]
- White, J.G.; Antos, M.J.; Fitzsimons, J.A.; Palmer, G.C. Non-Uniform Bird Assemblages in Urban Environments: The Influence of Streetscape Vegetation. Landsc. Urban Plan. 2005, 71, 123–135. [Google Scholar] [CrossRef]
- Tabachnick, B.; Fidell, L.; Ullman, J. Using Multivariate Statistics, 7th ed.; Taylor and Francis: Boston, MA, USA, 2007; Volume 5. [Google Scholar]
- McCune, B.; Mefford, M.J. PC-ORD: Multivariate Analysis of Ecological Data; MjM Software Design: Gleneden Beach, OR, USA, 2011. [Google Scholar]
- Fiske, I.; Chandler, R. Unmarked: An R Package for Fitting Hierarchical Models of Wildlife Occurrence and Abundance. J. Stat. Softw. 2011, 43, 1–23. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, G.; Friendly, M.; Kindt, R.; Legendre, P.; Minchin, P.; Simpson, G.; Solymos, P.; Stevens, M.; Szöcs, E.; et al. Vegan Community Ecology Package Version 2.5-7. Available online: https://www.researchgate.net/publication/346579465_vegan_community_ecology_package_version_25-7_November_2020 (accessed on 24 February 2025).
- Rega-Brodsky, C.; Nilon, C.; Warren, P. Balancing Urban Biodiversity Needs and Resident Preferences for Vacant Lot Management. Sustainability 2018, 10, 1679. [Google Scholar] [CrossRef]
- Blair, R.B. Land Use and Avian Species Diversity Along an Urban Gradient. Ecol. Appl. 1996, 6, 506–519. [Google Scholar] [CrossRef]
- Rega-Brodsky, C.; Nilon, C. Vacant Lots as a Habitat Resource: Nesting Success and Body Condition of Songbirds. Ecosphere 2016, 7, e01578. [Google Scholar] [CrossRef]
- Kyle, G.Z.; Kyle, P.D. Rehabilitation and Conservation of Chimney Swifts (Chaetura pelagica), 4th ed.; Driftwood Wildlife Association: Austin, TX, USA, 2004. [Google Scholar]
- Pennington, D.N.; Hansel, J.; Blair, R.B. The Conservation Value of Urban Riparian Areas for Landbirds during Spring Migration: Land Cover, Scale, and Vegetation Effects. Biol. Conserv. 2008, 141, 1235–1248. [Google Scholar] [CrossRef]
- Meffert, P.J.; Dziock, F. The Influence of Urbanisation on Diversity and Trait Composition of Birds. Landsc. Ecol. 2013, 28, 943–957. [Google Scholar] [CrossRef]
- MacGregor-Fors, I.; Schondube, J.E. Gray vs. Green Urbanization: Relative Importance of Urban Features for Urban Bird Communities. Basic Appl. Ecol. 2011, 12, 372–381. [Google Scholar] [CrossRef]
- Strohbach, M.W.; Lerman, S.B.; Warren, P.S. Are Small Greening Areas Enhancing Bird Diversity? Insights from Community-Driven Greening Projects in Boston. Landsc. Urban Plan. 2013, 114, 69–79. [Google Scholar] [CrossRef]
- Kim, G. The Public Value of Urban Vacant Land: Social Responses and Ecological Value. Sustainability 2016, 8, 486. [Google Scholar] [CrossRef]
- McPhearson, T.; Pickett, S.T.A.; Grimm, N.B.; Niemelä, J.; Alberti, M.; Elmqvist, T.; Weber, C.; Haase, D.; Breuste, J.; Qureshi, S. Advancing Urban Ecology toward a Science of Cities. BioScience 2016, 66, 198–212. [Google Scholar] [CrossRef]
- Haase, D.; Kabisch, S.; Haase, A.; Andersson, E.; Banzhaf, E.; Baró, F.; Brenck, M.; Fischer, L.K.; Frantzeskaki, N.; Kabisch, N.; et al. Greening Cities—To Be Socially Inclusive? About the Alleged Paradox of Society and Ecology in Cities. Habitat Int. 2017, 64, 41–48. [Google Scholar] [CrossRef]
- Verweij, E.E.; Pena, J.C.; Assis, J.C. Identifying Urban Hotspots of Potential Bird and Pedestrian Encounters through Corridor Overlap. Urban Ecosyst. 2025, 28, 73. [Google Scholar] [CrossRef]
- Michalczuk, J. Which Habitat Factors Affect the Occurrence and Richness of Cavity Nesters in Parks along an Urbanization Gradient? Recommendations for the Management of Greenery in an Urban Landscape. Sci. Total Environ. 2024, 927, 172091. [Google Scholar] [CrossRef]
- Spirn, A. Restoring Mill Creek: Landscape Literacy, Environmental Justice and City Planning and Design. Landsc. Res. 2005, 30, 395–413. [Google Scholar] [CrossRef]
- Spirn, A.; Pollio, M. A Resource for Reshaping Urban Neighborhoods; West Philadelphia Landscape Plan: Philadelphia, PA, USA, 1991; pp. 1–35. [Google Scholar]
- Pierce, R.A.; Nilon, C.H.; Stelzer, H.E. Assessing Wildlife Habitats and Natural Resources in Neighborhoods and Urban Environments: A Planning Guide for Wildlife Habitats in Urban Areas; Extension Guide MP927; University of Missouri: Columbia, MO, USA, 2013; Available online: https://extension.missouri.edu/publications/mp927 (accessed on 15 May 2025).
- Pierce, R.; Nilon, C.; Navarrete-Tindall, N.; Denkler, S.; Fishel, E. Improving Habitats for Wildlife in Your Backyard and Neighborhood; Extension Publication G9500; University of Missouri: Columbia, MO, USA, 2015; Available online: https://extension.missouri.edu/publications/g9500 (accessed on 15 May 2025).
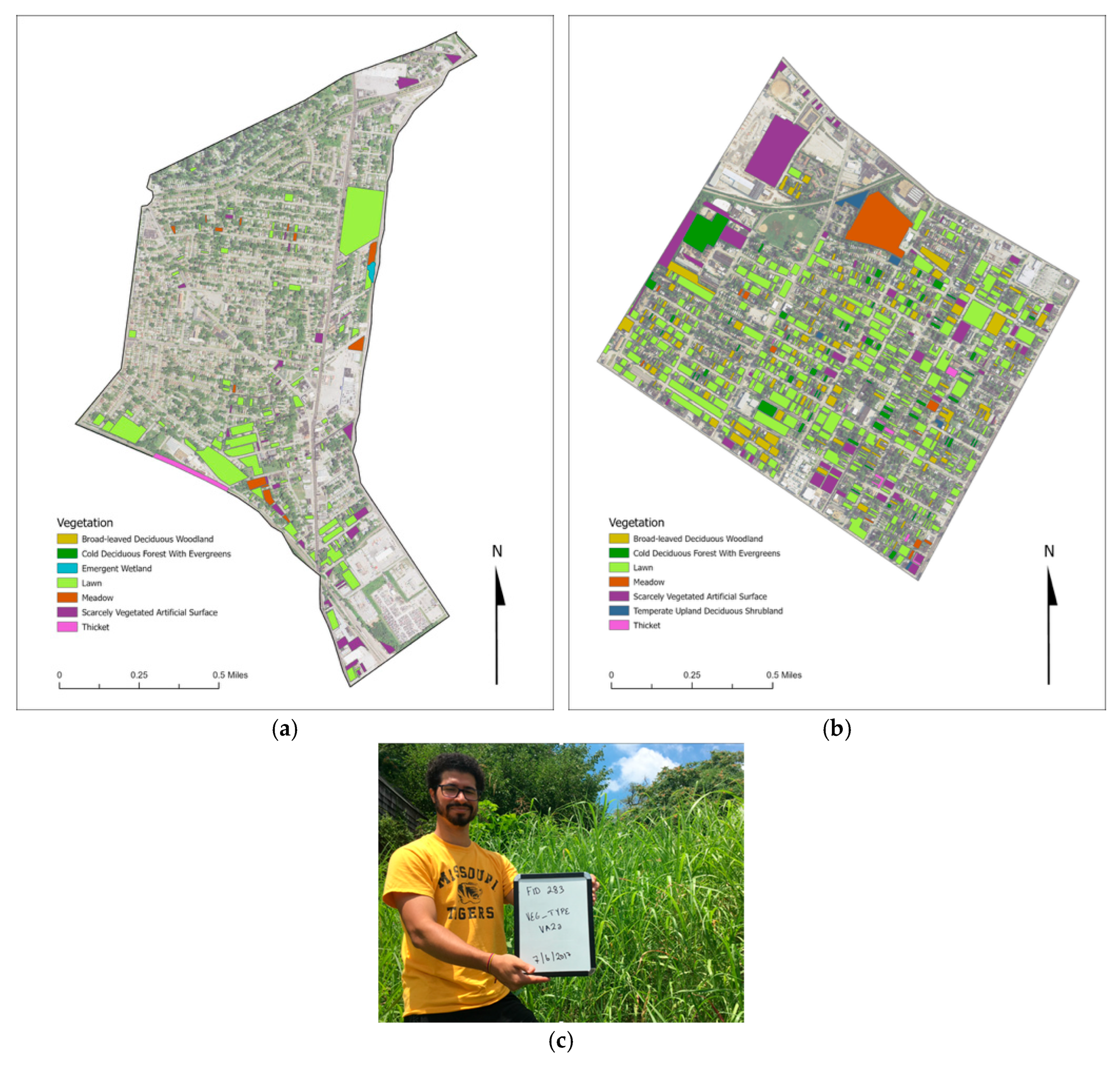

{kind=link}
{kind=link}
| Code | Form Class | Form Subclass | Form Group | Form Subgroup |
|---|---|---|---|---|
| VA2b | Herbaceous vegetation | Terrestrial herbaceous vegetation | Meadow and lawn | Lawn |
| VA2a | Herbaceous vegetation | Terrestrial herbaceous vegetation | Meadow and lawn | Meadow |
| VID | Beaches/scarcely vegetated | Scarcely vegetated artificial | ---------- | Artificial surface |
| IB2a | Closed forest | Temperate deciduous forest | Cold deciduous forest and pines | Cold deciduous forest with evergreen |
| IIB2a | Woodland | Deciduous woodland | Cold deciduous woodland | Broad-leaved deciduous woodland |
| IIIA1 | Scrub | Thicket | Mainly deciduous | ---------- |
| IIIB1a | Scrub | Shrubland | Mainly deciduous | Temperate upland deciduous shrubland |
| VB1a | Herbaceous vegetation | Semi-aquatic herbaceous vegetation | Freshwater vegetation | Emergent wetland |
| Vegetation Layer | Structure Type | Mean Cover (%) | Standard Deviation (%) |
|---|---|---|---|
| Ground | Artificial surface | 39 | 19.3 |
| Bare soil | 4.5 | 4.3 | |
| Forb | 8.7 | 7.8 | |
| Grass | 33.8 | 15.7 | |
| Leaf litter | 5.1 | 4.6 | |
| Woody litter | 1.6 | 1.9 | |
| Vine | 0.8 | 1.2 | |
| Rock | 3.2 | 9.1 | |
| Water | 0.01 | 0.09 | |
| Shrub | Shrub cover | 7.1 | 7.5 |
| Canopy | Canopy cover | 7.8 | 7.6 |
| Vacant Lot | Total vacant lot cover | 33.6 | 20.7 |
| Lawn (VA2b) | 16.5 | 20.9 | |
| Meadow (VA2a) | 4.3 | 10.9 | |
| Artificial surface (VID) | 3.4 | 8.4 | |
| Deciduous forest with evergreens (IB2a) | 1.8 | 5.4 |
| Vacant Lot Type | % Cover |
|---|---|
| Lawn (VA2b) | 49.1 |
| Meadow (VA2a) | 12.8 |
| Artificial surface (VID) | 10.6 |
| Deciduous forest with evergreens (IB2a) | 5.5 |
| Deciduous woodland (IIB2a) | 15.5 |
| Thicket (IIIA1) | 3.7 |
| Shrubland (IIIB1a) | 2.9 |
| Emergent wetland (VB1a) | 0.8 |
| Variable | PC1 | PC2 | PC3 | PC4 |
|---|---|---|---|---|
| Artificial surface | −0.400 | 0.306 | 0.120 | 0.080 |
| Bare soil | 0.251 | 0.054 | −0.337 | −0.126 |
| Forb | 0.314 | 0.054 | −0.337 | −0.126 |
| Grass | 0.314 | −0.034 | −0.067 | −0.034 |
| Leaf litter | 0.320 | −0.080 | 0.123 | −0.215 |
| Woody litter | 0.313 | 0.052 | −0.032 | −0.105 |
| Canopy layer | 0.271 | 0.113 | 0.096 | 0.360 |
| Shrub layer | 0.258 | 0.182 | −0.044 | 0.386 |
| Vine | 0.123 | 0.163 | 0.279 | −0.214 |
| Gravel | 0.080 | 0.137 | −0.532 | −0.245 |
| Water | −0.097 | 0.090 | 0.020 | −0.213 |
| Shrub cover | 0.348 | 0.259 | 0.033 | −0.084 |
| Canopy cover | 0.297 | 0.163 | 0.368 | −0.0091 |
| Vacant percentage | 0.083 | −0.459 | −0.139 | 0.052 |
| Lawn (VA2b) | 0.088 | −0.477 | −0.050 | −0.140 |
| Artificial surface (VID) | −0.219 | 0.118 | −0.047 | −0.125 |
| Deciduous forest with evergreens (IB2a) | 0.087 | 0.018 | −0.163 | −0.125 |
| Deciduous woodland (IIB2a) | 0.088 | −0.053 | 0.351 | 0.071 |
| Thicket (IIIA1) | 0.088 | 0.024 | −0.153 | −0.076 |
| Emergent wetland (VB1a) | 0.044 | 0.042 | −0.091 | 0.340 |
| Meadow (VA2a) | −0.045 | −0.066 | −0.250 | 0.353 |
| Shrubland (IIIB1a) | 0.087 | 0.108 | −0.151 | 0.360 |
| Eigenvalue | 3.125 | 2.668 | 1.743 | 1.667 |
| Percent variance explained | 14.2% | 12.1% | 7.9% | 7.6% |
| Species | Top Model Covariates | Effect Size (β) |
|---|---|---|
| American Robin | % Canopy Cover | 1.9 |
| Northern Cardinal | % Canopy Cover | 1.9 |
| Common Grackle | % Closed Forest (+), % Thicket (−) | 2.8, −6.3 |
| European Starling | % Shrub Cover (+), % Forest Cover (−) | 5.8, −3.2 |
| Chimney Swift | Global model with % Forest, % Thicket | 2.8 (forest), −5.9 (thicket) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno, S.; Mallinak, A.J.; Nilon, C.H.; Pierce, R.A., II. A Beautiful Bird in the Neighborhood: Canopy Cover and Vegetation Structure Predict Avian Presence in High-Vacancy City. Land 2025, 14, 1433. https://doi.org/10.3390/land14071433
Moreno S, Mallinak AJ, Nilon CH, Pierce RA II. A Beautiful Bird in the Neighborhood: Canopy Cover and Vegetation Structure Predict Avian Presence in High-Vacancy City. Land. 2025; 14(7):1433. https://doi.org/10.3390/land14071433
Chicago/Turabian StyleMoreno, Sebastian, Andrew J. Mallinak, Charles H. Nilon, and Robert A. Pierce, II. 2025. "A Beautiful Bird in the Neighborhood: Canopy Cover and Vegetation Structure Predict Avian Presence in High-Vacancy City" Land 14, no. 7: 1433. https://doi.org/10.3390/land14071433
APA StyleMoreno, S., Mallinak, A. J., Nilon, C. H., & Pierce, R. A., II. (2025). A Beautiful Bird in the Neighborhood: Canopy Cover and Vegetation Structure Predict Avian Presence in High-Vacancy City. Land, 14(7), 1433. https://doi.org/10.3390/land14071433