
Multidimensional Bird Habitat Network Resilience Assessment and Ecological Strategic Space Identification in International Wetland City



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
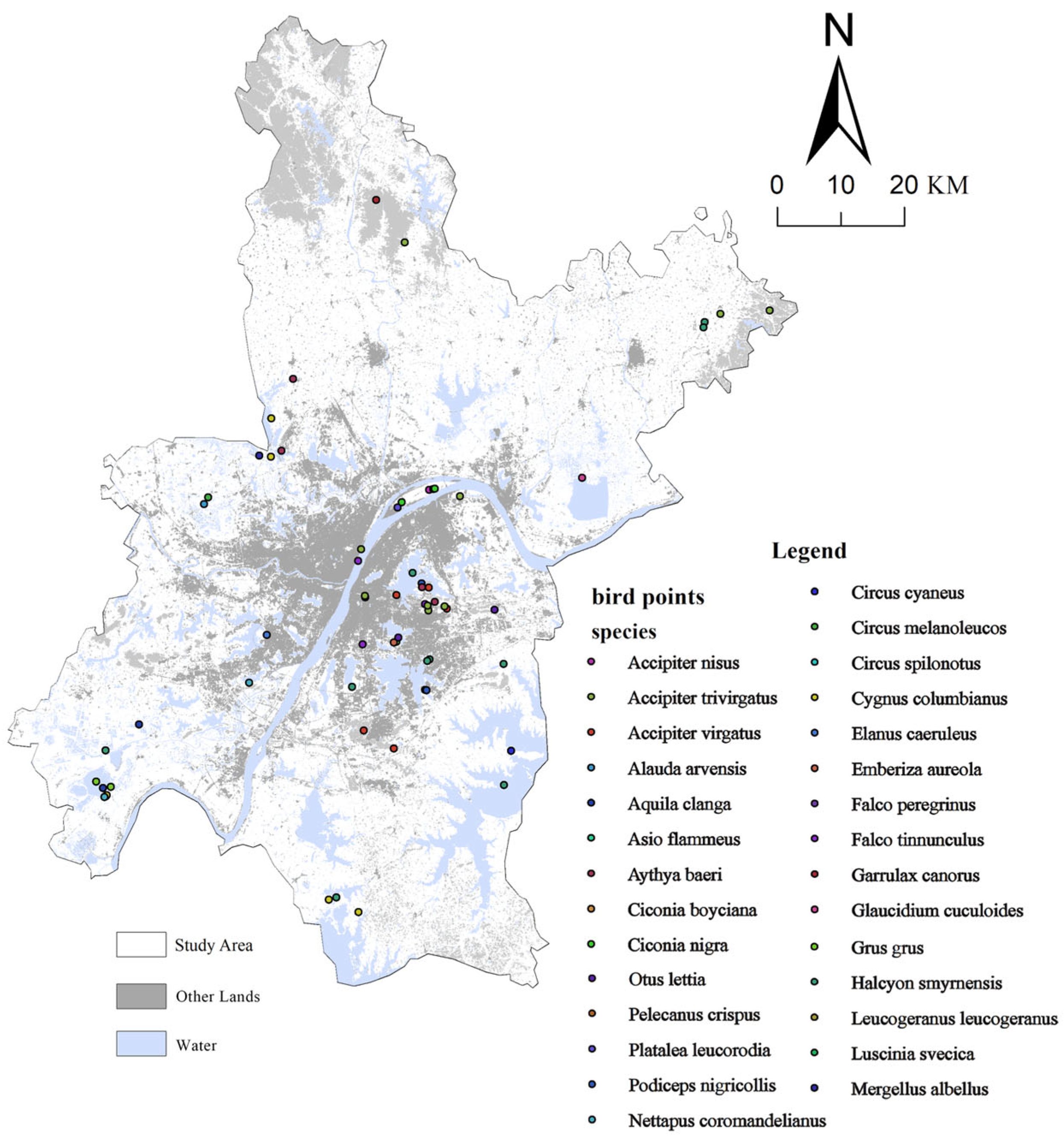
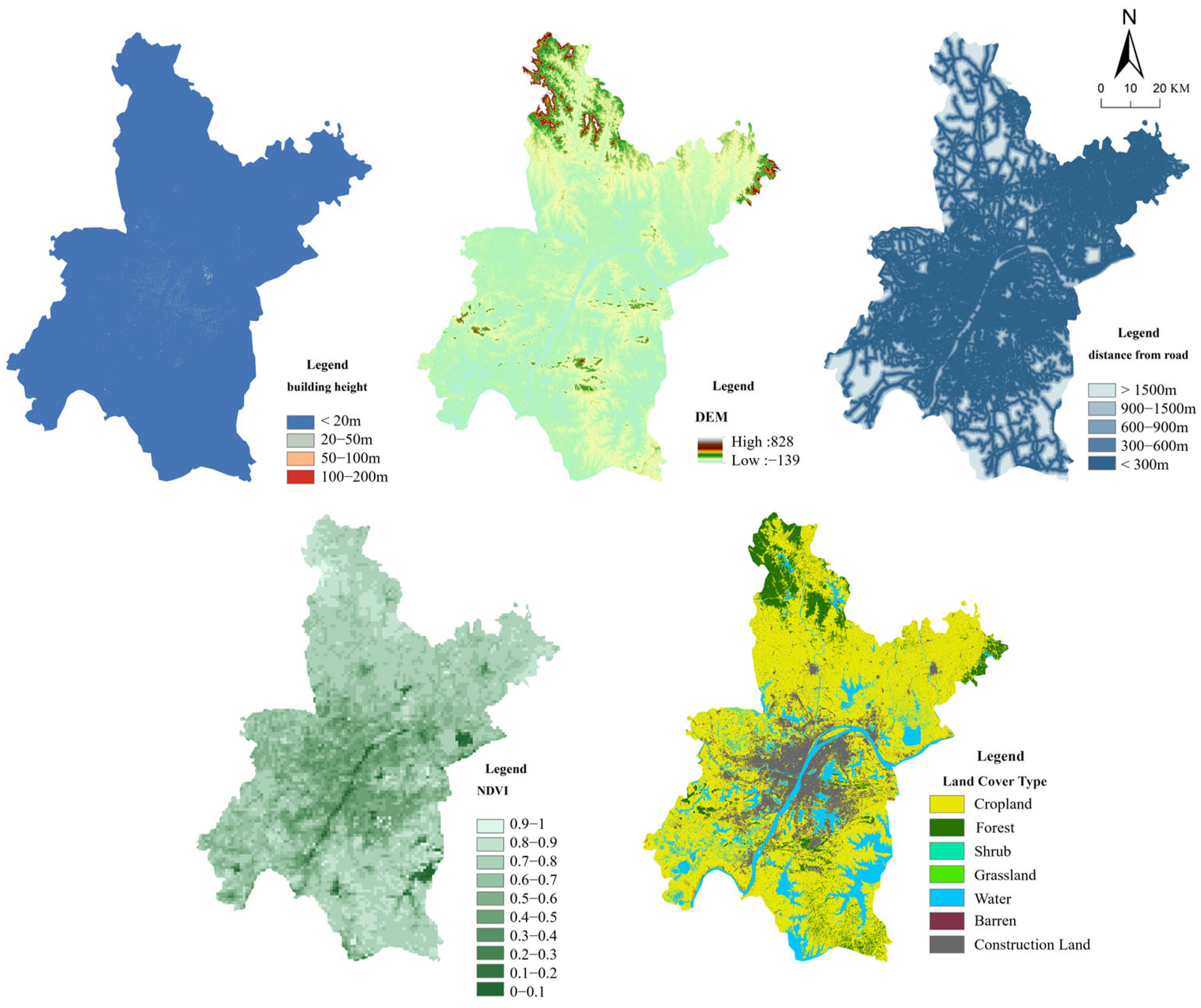
2.2. Data Sources and Preprocessing
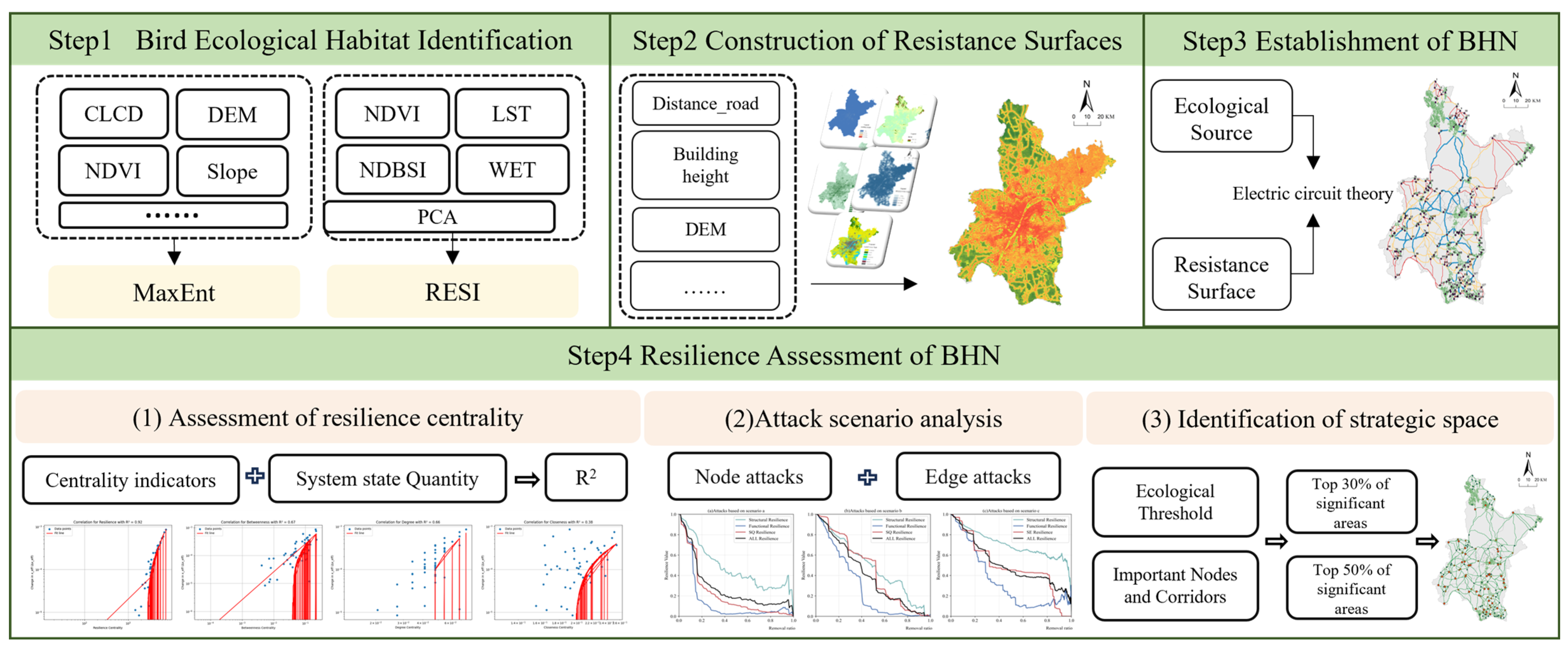
2.3. Methodology
2.3.1. Identification of Bird Habitat Patches
Habitat Suitability Assessment Based on MaxEnt
Habitat Patch Extraction Based on RSEI
2.3.2. Resistance Surface Construction
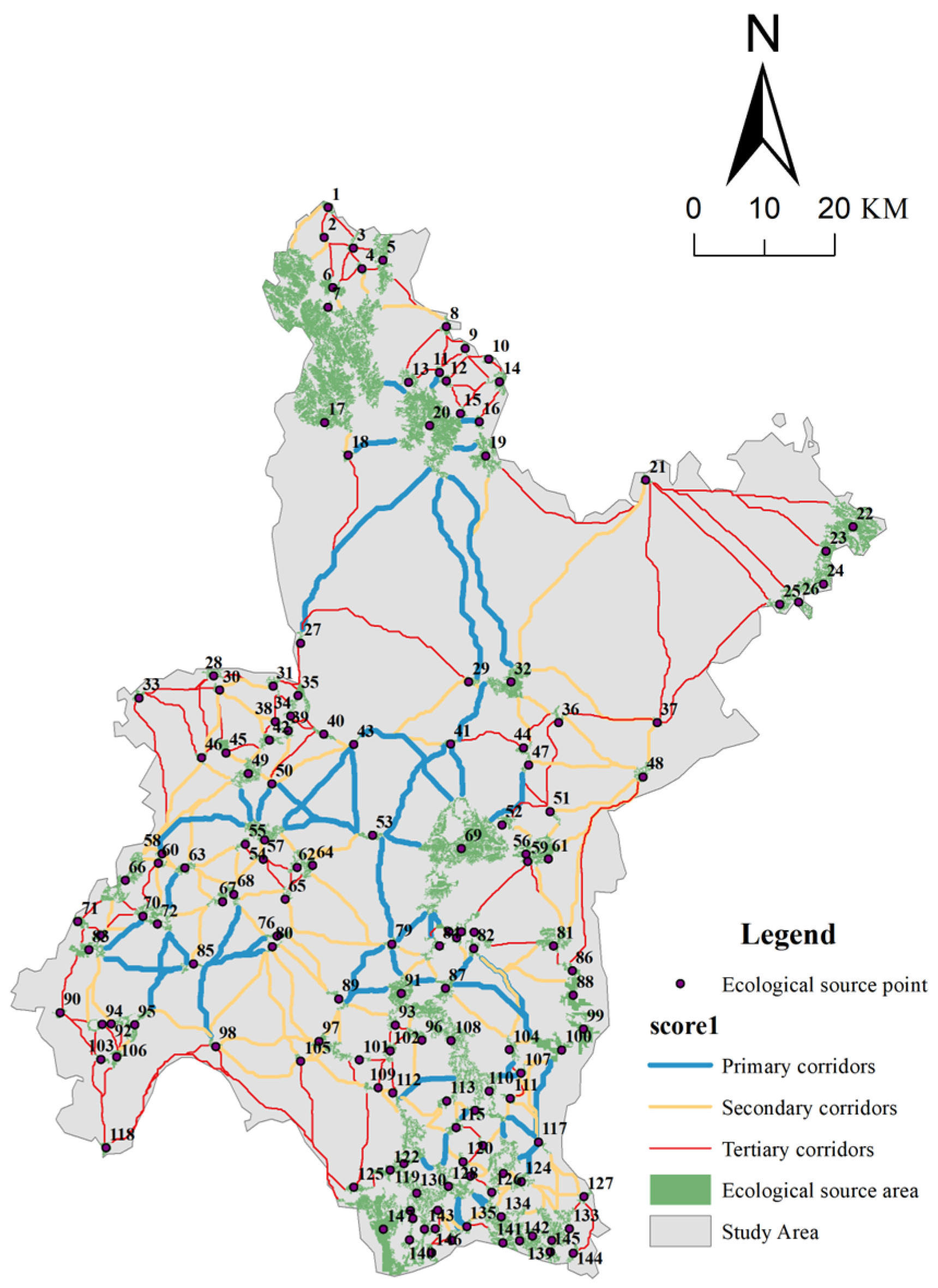
2.3.3. Identification of Ecological Corridors on Circuit Theory
2.3.4. Resilience Assessment of the Bird Habitat Network
Evaluating Resilience Centrality in the BHN
Ecological Network Disturbance Scenarios
Ecological Network Resilience Indicator Calculation
3. Results
3.1. Habitat Identification
3.2. Ecological Network Construction
3.3. Network Resilience Analysis
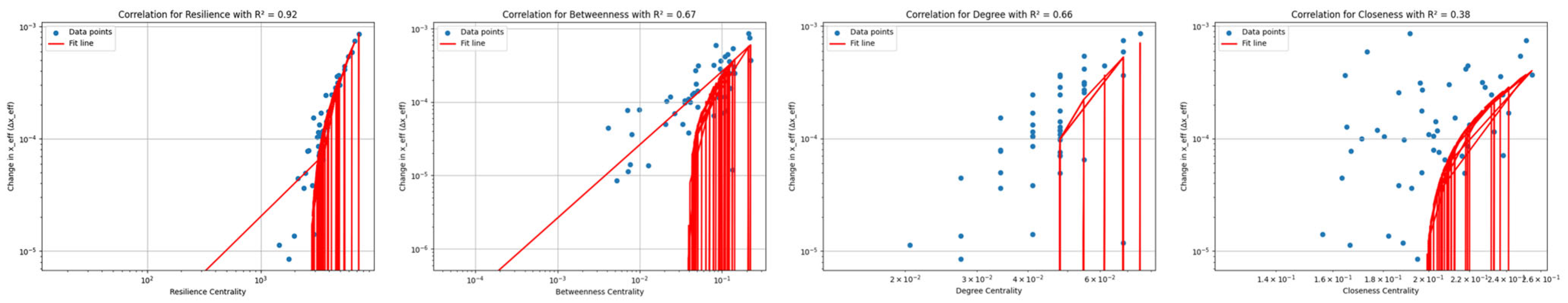
3.3.1. Advantages of Resilience Centrality
3.3.2. Network Resilience Assessment
4. Discussion
4.1. Ecological Security Pattern in International Wetland Cities
4.2. Contributions to the Study of Spatial Resilience
4.3. Spatial Management Policies to Enhance Urban Ecological Network Resilience
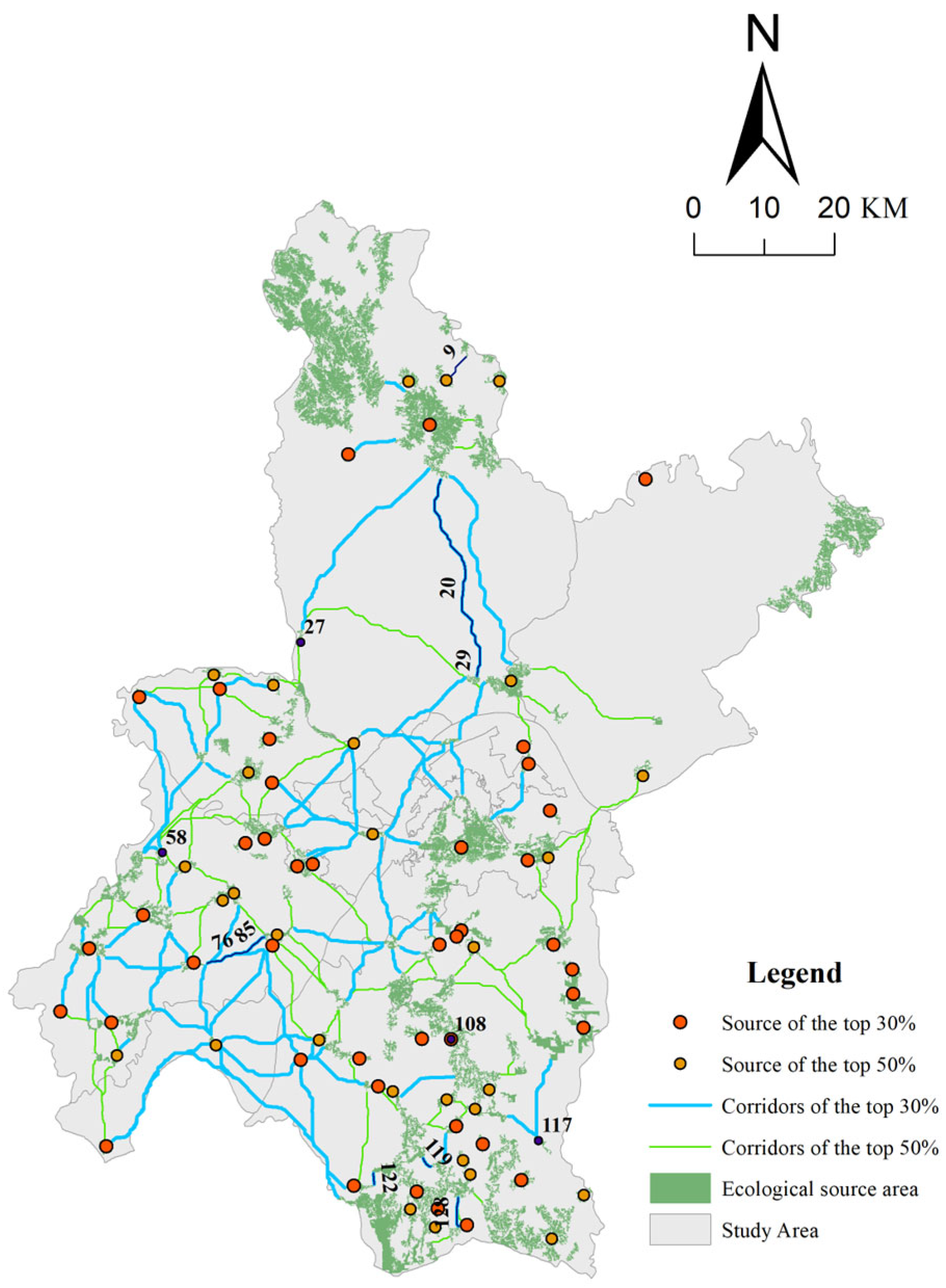
4.3.1. Strategic Space Identification
4.3.2. Planning Control Strategies
4.4. Limitations and Future Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| BHN | Bird Habitat Network |
| EAAF | East Asian–Australasian Flyway |
| RSEI | Remote Sensing Ecological Index |
| MSPA | morphological spatial pattern analysis |
| AHP | analytic hierarchy process |
| MaxEnt | Maximum Entropy |
| SDMs | species distribution models |
References
- Goyette, J.-O.; Cimon-Morin, J.; Mendes, P.; Thériault, M.; Pellerin, S.; Poulin, M. Planning wetland protection and restoration for the safeguard of ecosystem service flows to beneficiaries. Landsc. Ecol. 2021, 36, 2691–2706. [Google Scholar] [CrossRef]
- Luo, J.; Zhu, L.; Fu, H. Construction of wetland ecological network based on MSPA-Conefor-MCR: A case study of Haikou City. Ecol. Indic. 2024, 166, 112329. [Google Scholar] [CrossRef]
- Qu, Y.; Gong, H.; Zheng, Y.; Shi, J.; Zeng, X.; Yang, H.; Wang, J.; Zhenguo, N.; Li, L.; Wang, S.; et al. Global conservation priorities for wetlands and setting post-2025 targets. Commun. Earth Environ. 2024, 5, 4. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, X.; Cui, L.; Lei, Y.; Guo, Z.; Wang, J.; Li, J.; Zhai, X.; Wang, R.; Li, W. Identification and optimization of urban wetland ecological networks in highly urbanized areas: A case study of Haidian District, Beijing. Ecol. Indic. 2025, 170, 113028. [Google Scholar] [CrossRef]
- Zhu, L.; Zhu, K.; Zeng, X. Evolution of landscape pattern and response of ecosystem service value in international wetland cities: A case study of Nanchang City. Ecol. Indic. 2023, 155, 110987. [Google Scholar] [CrossRef]
- Xu, W.; Yang, H.; Chen, Z.; Shi, R.; Liu, Y.; Chen, J. Enhancing bird habitat networks in metropolitan areas: Resilience assessment and improvement strategies—A case study from Shanghai. Sci. Total Environ. 2024, 954, 176316. [Google Scholar] [CrossRef]
- Jiang, X.; Mao, D.; Zhen, J.; Wang, J.; Van de Voorde, T. Exploring the conservation of historic avian corridors under urbanization threats in China: A case study of egrets in the Greater Bay Area. Sci. Total Environ. 2024, 948, 174921. [Google Scholar] [CrossRef]
- Ikin, K.; Yong, D.L.; Lindenmayer, D.B. Effectiveness of woodland birds as taxonomic surrogates in conservation planning for biodiversity on farms. Biol. Conserv. 2016, 204, 411–416. [Google Scholar] [CrossRef]
- Eastough, A.; Hughes, M.; Newsome, D. Investigating the extent that different recreational users of urban wetlands value birds and bird habitat. Int. J. Geoheritage Parks 2024, 12, 434–445. [Google Scholar] [CrossRef]
- Wu, Z.; Cheng, S.; Xu, K.; Qian, Y. Ecological network resilience evaluation and ecological strategic space identification based on complex network theory: A case study of Nanjing city. Ecol. Indic. 2024, 158, 111604. [Google Scholar] [CrossRef]
- Forman, R.T.T. Some general principles of landscape and regional ecology. Landsc. Ecol. 1995, 10, 133–142. [Google Scholar] [CrossRef]
- Yu, K. Security patterns and surface model in landscape ecological planning. Landsc. Urban Plan. 1996, 36, 1–17. [Google Scholar] [CrossRef]
- Peng, J.; Yang, Y.; Liu, Y.X.; Hu, Y.N.; Du, Y.Y.; Meersmans, J.; Qiu, S.J. Linking ecosystem services and circuit theory to identify ecological security patterns. Sci. Total Environ. 2018, 644, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Wang, X.; Li, J.; Zhang, M.; Chen, S.; Xu, B.; Nie, W. Multi-Scale Applicability Analysis of Three Ecological Network Construction Methods in Resilience Assessment. Land Degrad. Dev. 2025, 36, 1594–1613. [Google Scholar] [CrossRef]
- Hashemi, R.; Darabi, H.; Hashemi, M.; Wang, J. Graph theory in ecological network analysis: A systematic review for connectivity assessment. J. Clean. Prod. 2024, 472, 143504. [Google Scholar] [CrossRef]
- Wang, S.; Wu, M.; Hu, M.; Fan, C.; Wang, T.; Xia, B. Promoting landscape connectivity of highly urbanized area: An ecological network approach. Ecol. Indic. 2021, 125, 107487. [Google Scholar] [CrossRef]
- Li, S.; Zhao, Y.; Xiao, W.; Yue, W.; Wu, T. Optimizing ecological security pattern in the coal resource-based city: A case study in Shuozhou City, China. Ecol. Indic. 2021, 130, 108026. [Google Scholar] [CrossRef]
- Dong, X.; Wang, F.; Fu, M. Research progress and prospects for constructing ecological security pattern based on ecological network. Ecol. Indic. 2024, 168, 112800. [Google Scholar] [CrossRef]
- Yang, H.; Xu, W.; Chen, Z.; Xie, X.; Yu, J.; Lei, X.; Guo, S.; Ding, Z. Ecological network construction for bird communities in high-density urban areas: A perspective of integrated approaches. Ecol. Indic. 2024, 158, 111592. [Google Scholar] [CrossRef]
- Shen, Z.; Yin, H.; Kong, F.; Wu, W.; Sun, H.; Su, J.; Tian, S. Enhancing ecological network establishment with explicit species information and spatially coordinated optimization for supporting urban landscape planning and management. Landsc. Urban Plan. 2024, 248, 105079. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, Y.; Zeng, D.; Yang, Y.; Li, W.; Kang, Y.; Wei, G.; Yuan, X.; Bo, S.; Si, X. Characterizing bird species for achieving the win-wins of conserving biodiversity and enhancing regulating ecosystem services in urban green spaces. Urban For. Urban Green. 2023, 87, 128064. [Google Scholar] [CrossRef]
- Pan, J.; Liang, J.; Zhao, C. Identification and optimization of ecological security pattern in arid inland basin based on ordered weighted average and ant colony algorithm: A case study of Shule River basin, NW China. Ecol. Indic. 2023, 154, 110588. [Google Scholar] [CrossRef]
- Song, S.; Gong, Y.; Yu, Y. Integrating pattern, process, and function in urban landscape ecological network planning: A case study of Harbin central city. Ecol. Indic. 2024, 159, 111671. [Google Scholar] [CrossRef]
- Liu, Z.; Huang, Q.; Tang, G. Identification of urban flight corridors for migratory birds in the coastal regions of Shenzhen city based on three-dimensional landscapes. Landsc. Ecol. 2021, 36, 2043–2057. [Google Scholar] [CrossRef]
- Lin, J.; Wang, Y.; Lin, Z.; Li, S. National-scale connectivity analysis and construction of forest networks based on graph theory: A case study of China. Ecol. Eng. 2025, 216, 107639. [Google Scholar] [CrossRef]
- Geng, J.; Yu, K.; Sun, M.; Xie, Z.; Huang, R.; Wang, Y.; Zhao, Q.; Liu, J. Construction and Optimisation of Ecological Networks in High-Density Central Urban Areas: The Case of Fuzhou City, China. Remote Sens. 2023, 15, 5666. [Google Scholar] [CrossRef]
- Men, D.; Pan, J. Incorporating network topology and ecosystem services into the optimization of ecological network: A case study of the Yellow River Basin. Sci. Total Environ. 2024, 912, 169004. [Google Scholar] [CrossRef]
- Su, H.; Du, M.; Liu, Q.; Kang, X.; Zhao, L.; Zheng, W.; Liao, Z. Assessment of regional Ecosystem Service Bundles coupling climate and land use changes. Ecol. Indic. 2024, 169, 112844. [Google Scholar] [CrossRef]
- Zhao, J.; Yu, Q.; Xu, C.; Ma, J.; Liu, W.; Sun, W.; Miao, Y.; Nawaz, T. Integrated approach for ecological restoration and ecological spatial network optimization with multiple ecosystem functions in mining areas. Ecol. Indic. 2023, 156, 111141. [Google Scholar] [CrossRef]
- Yang, H.; Yan, S.; Wang, X.; Li, C.; Meng, H.; Yao, Q. Constructing Ecological Networks Based on Ecosystem Services and Network Analysis in Chongqing, China. Land 2024, 13, 662. [Google Scholar] [CrossRef]
- Lu, Y.; Liu, Y.; Huang, D.; Liu, Y. Evolution Analysis of Ecological Networks Based on Spatial Distribution Data of Land Use Types Monitored by Remote Sensing in Wuhan Urban Agglomeration, China, from 2000 to 2020. Remote Sens. 2022, 14, 2618. [Google Scholar] [CrossRef]
- Xiang, Q.; Yu, H.; Huang, H.; Li, F.; Ju, L.; Hu, W.; Yu, P.; Deng, Z.; Chen, Y. Assessing the resilience of complex ecological spatial networks using a cascading failure model. J. Clean. Prod. 2024, 434, 140014. [Google Scholar] [CrossRef]
- Shi, J.; Zhang, D.; Yin, J.; Wang, Z.; Feng, H.; Liu, Y.; Shi, P. Regional ecosystem health improvement pathway design based on ecological network optimization. Ecol. Indic. 2025, 173, 113377. [Google Scholar] [CrossRef]
- Wang, T.; Li, H.; Huang, Y. The complex ecological network’s resilience of the Wuhan metropolitan area. Ecol. Indic. 2021, 130, 108101. [Google Scholar] [CrossRef]
- Zhou, Y.; Zheng, Z.; Wu, Z.; Guo, C.; Chen, Y. Construction and evaluation of ecological networks in highly urbanised regions: A case study of the Guangdong-Hong Kong-Macao greater Bay Area, China. Ecol. Indic. 2023, 152, 110336. [Google Scholar] [CrossRef]
- Wang, T.; Huang, Y.; Cheng, J.; Xiong, H.; Ying, Y.; Feng, Y.; Wang, J. Construction and optimization of watershed-scale ecological network based on complex network method: A case study of Erhai Lake Basin in China. Ecol. Indic. 2024, 160, 111794. [Google Scholar] [CrossRef]
- Cumming, G.S. Spatial resilience: Integrating landscape ecology, resilience, and sustainability. Landsc. Ecol. 2011, 26, 899–909. [Google Scholar] [CrossRef]
- Kéfi, S.; Saade, C.; Berlow, E.; Sarmento Cabral, J.; Fronhofer, E. Scaling up our understanding of tipping points. Philos. Trans. R. Soc. B Biol. Sci. 2022, 377, 20210386. [Google Scholar] [CrossRef]
- Huang, L.; Wang, J.; Cheng, H. Spatiotemporal changes in ecological network resilience in the Shandong Peninsula urban agglomeration. J. Clean. Prod. 2022, 339, 130681. [Google Scholar] [CrossRef]
- McCloy, M.W.D.; Andringa, R.K.; Maness, T.J.; Smith, J.A.; Grace, J.K. Promoting urban ecological resilience through the lens of avian biodiversity. Front. Ecol. Evol. 2024, 12, 1302002. [Google Scholar] [CrossRef]
- McCloy, M.W.D.; Andringa, R.K.; Grace, J.K. Resilience of Avian Communities to Urbanization and Climate Change: An Integrative Review. Front. Conserv. Sci. 2022, 3, 918873. [Google Scholar] [CrossRef]
- Epele, L.B.; Grech, M.G.; Manzo, L.M.; Macchi, P.A.; Hermoso, V.; Miserendino, M.L.; Bonada, N.; Cañedo-Argüelles, M. Identifying high priority conservation areas for Patagonian wetlands biodiversity. Biodivers. Conserv. 2021, 30, 1359–1374. [Google Scholar] [CrossRef]
- Hong, W.; Guo, R.; Li, X.; Liao, C. Measuring urban ecological network resilience: A disturbance scenario simulation method. Cities 2022, 131, 104057. [Google Scholar] [CrossRef]
- Tan, C.; Ma, J.; Yan, Y.; Hong, G.; Wu, X. Identifying priority wetlands for the conservation of waterbirds in Wuhan, China. Ecol. Indic. 2023, 157, 111240. [Google Scholar] [CrossRef]
- Greening, W. Wuhan Shares Wetland Conservation Experience with the World. Available online: https://ylj.wuhan.gov.cn/xwdt/mtbd_12295/202410/t20241017_2469620.shtml (accessed on 10 March 2025).
- Cui, X.Y.; Li, H.B.; Wang, T. Structural Evolution and Stability Simulation of Wetland Ecological Network: A Case Study of Wuhan City, China. Wetlands 2022, 42, 73. [Google Scholar] [CrossRef]
- Zhai, H.; Lv, C.; Liu, W.; Yang, C.; Fan, D.; Wang, Z.; Guan, Q. Understanding Spatio-Temporal Patterns of Land Use/Land Cover Change under Urbanization in Wuhan, China, 2000–2019. Remote Sens. 2021, 13, 3331. [Google Scholar] [CrossRef]
- Gao, J.; Gong, J.; Li, Y.; Yang, J.; Liang, X. Ecological network assessment in dynamic landscapes: Multi-scenario simulation and conservation priority analysis. Land Use Policy 2024, 139, 107059. [Google Scholar] [CrossRef]
- Poisson, A.C.; McCullough, I.M.; Cheruvelil, K.S.; Elliott, K.C.; Latimore, J.A.; Soranno, P.A. Quantifying the contribution of citizen science to broad-scale ecological databases. Front. Ecol. Environ. 2020, 18, 19–26. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef]
- Xu, C.; Yu, Q.; Wang, F.; Qiu, S.; Ai, M.; Zhao, J. Identifying and optimizing ecological spatial patterns based on the bird distribution in the Yellow River Basin, China. J. Environ. Manag. 2023, 348, 119293. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Liu, M.; Qu, M.; Guang, X.; Chen, J.; Zhao, Y.; Huang, B. Identifying the potential soil pollution areas derived from the metal mining industry in China using MaxEnt with mine reserve scales (MaxEnt_MRS). Environ. Pollut. 2023, 329, 121687. [Google Scholar] [CrossRef]
- Shi, F.; Liu, S.; Sun, Y.; An, Y.; Zhao, S.; Liu, Y.; Li, M. Ecological network construction of the heterogeneous agro-pastoral areas in the upper Yellow River basin. Agric. Ecosyst. Environ. 2020, 302, 107069. [Google Scholar] [CrossRef]
- Shen, Z.; Wu, W.; Chen, S.; Tian, S.; Wang, J.; Li, L. A static and dynamic coupling approach for maintaining ecological networks connectivity in rapid urbanization contexts. J. Clean. Prod. 2022, 369, 133375. [Google Scholar] [CrossRef]
- Sun, D.; Wu, X.; Wen, H.; Ma, X.; Zhang, F.; Ji, Q.; Zhang, J. Ecological Security Pattern based on XGBoost-MCR model: A case study of the Three Gorges Reservoir Region. J. Clean. Prod. 2024, 470, 143252. [Google Scholar] [CrossRef]
- Tremblay, M.A.; St. Clair, C.C. Permeability of a heterogeneous urban landscape to the movements of forest songbirds. J. Appl. Ecol. 2011, 48, 679–688. [Google Scholar] [CrossRef]
- Bahía, R.; Lambertucci, S.A.; Speziale, K.L. Anthropogenic city noise affects the vocalizations of key forest birds. Biodivers. Conserv. 2024, 33, 2405–2421. [Google Scholar] [CrossRef]
- Konstantopoulos, K.; Moustakas, A.; Vogiatzakis, I.N. A spatially explicit impact assessment of road characteristics, road-induced fragmentation and noise on birds species in Cyprus. Biodiversity 2020, 21, 61–71. [Google Scholar] [CrossRef]
- Cicort-Lucaciu, A.; Keshta, H.V.; Popovici, P.V.; Munkácsi, D.; Telcean, I.C.; Gache, C. Urban avifauna distribution explained by road noise in an Eastern European city. Avian Res. 2022, 13, 100067. [Google Scholar] [CrossRef]
- Fijn, R.C.; Krijgsveld, K.L.; Poot, M.J.M.; Dirksen, S. Bird movements at rotor heights measured continuously with vertical radar at a Dutch offshore wind farm. Ibis 2015, 157, 558–566. [Google Scholar] [CrossRef]
- Hager, S.B.; Cosentino, B.J.; McKay, K.J.; Monson, C.; Zuurdeeg, W.; Blevins, B. Window area and development drive spatial variation in bird-window collisions in an urban landscape. PLoS ONE 2013, 8, e53371. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Carruthers-Jones, J.; Carver, S.; Wu, J. Constructing urban ecological corridors to reflect local species diversity and conservation objectives. Sci. Total Environ. 2024, 907, 167987. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, G. Human-Caused High Direct Mortality in Birds: Unsustainable Trends and Ameliorative Actions. Animals 2024, 15, 73. [Google Scholar] [CrossRef]
- Wang, X.; Xie, X.; Wang, Z.; Lin, H.; Liu, Y.; Xie, H.; Liu, X. Construction and Optimization of an Ecological Security Pattern Based on the MCR Model: A Case Study of the Minjiang River Basin in Eastern China. Int. J. Environ. Res. Public Health 2022, 19, 8370. [Google Scholar] [CrossRef]
- Li, J.; Nie, W.; Zhang, M.; Wang, L.; Dong, H.; Xu, B. Assessment and optimization of urban ecological network resilience based on disturbance scenario simulations: A case study of Nanjing city. J. Clean. Prod. 2024, 438, 140812. [Google Scholar] [CrossRef]
- Liu, H.; Wang, Z.; Zhang, L.; Tang, F.; Wang, G.; Li, M. Construction of an ecological security network in the Fenhe River Basin and its temporal and spatial evolution characteristics. J. Clean. Prod. 2023, 417, 137961. [Google Scholar] [CrossRef]
- Zhang, Y.; Shao, C.; He, S.; Gao, J. Resilience centrality in complex networks. Phys. Rev. E 2020, 101, 022304. [Google Scholar] [CrossRef] [PubMed]
- De Montis, A.; Ganciu, A.; Cabras, M.; Bardi, A.; Peddio, V.; Caschili, S.; Massa, P.; Cocco, C.; Mulas, M. Resilient ecological networks: A comparative approach. Land Use Policy 2019, 89, 104207. [Google Scholar] [CrossRef]
- Wang, Y.; Cai, Y.; Xie, Y.; Chen, L.; Zhang, P. An integrated approach for evaluating dynamics of urban eco-resilience in urban agglomerations of China. Ecol. Indic. 2023, 146, 109859. [Google Scholar] [CrossRef]
- Chen, H.; Yan, W.; Li, Z.; Wende, W.; Xiao, S. A framework for integrating ecosystem service provision and connectivity in ecological spatial networks: A case study of the Shanghai metropolitan area. Sustain. Cities Soc. 2024, 100, 105018. [Google Scholar] [CrossRef]
- Carey, M.; Boland, J.; Keppel, G. Generalized Logarithmic Species-Area Relationship Resolves the Arrhenius-Gleason Debate. Environ. Model. Assess. 2023, 28, 491–499. [Google Scholar] [CrossRef]
- Gurka, M.J.; Edwards, L.J.; Nylander-French, L. Testing transformations for the linear mixed model. Comput. Stat. Data Anal. 2007, 51, 4297–4307. [Google Scholar] [CrossRef]
- Kong, F.; Yin, H.; Nakagoshi, N.; Zong, Y. Urban green space network development for biodiversity conservation: Identification based on graph theory and gravity modeling. Landsc. Urban Plan. 2010, 95, 16–27. [Google Scholar] [CrossRef]
- Zheng, Q.; Zeng, J.; Luo, J.; Cui, J.; Sun, X. Spatial Structure and Space Governance of Ecological Network in Wuhan City. Econ. Geogr. 2018, 38, 191–199. [Google Scholar] [CrossRef]
- Yan, Y.; Gan, X.; Zhao, Y.; Liao, Q.; Huang, C. Wetland Landscape Dynamics and Multi-Functional Assessment of Wuhan City, China. Water 2024, 16, 2914. [Google Scholar] [CrossRef]
- Wang, L.; Li, Z.; Zhang, Z. City profile: Wuhan 2004–2020. Cities 2022, 123, 103585. [Google Scholar] [CrossRef]
- Qian, M.; Huang, Y.; Cao, Y.; Wu, J.; Xiong, Y. Ecological network construction and optimization in Guangzhou from the perspective of biodiversity conservation. J. Environ. Manag. 2023, 336, 117692. [Google Scholar] [CrossRef]
- Saunders, D.A.; Hobbs, R.J.; Margules, C.R. Biological Consequences of Ecosystem Fragmentation: A Review. Conserv. Biol. 1991, 5, 18–32. [Google Scholar] [CrossRef]
- Tarabon, S.; Dutoit, T.; Isselin-Nondedeu, F. Pooling biodiversity offsets to improve habitat connectivity and species conservation. J. Environ. Manag. 2021, 277, 111425. [Google Scholar] [CrossRef]
- Beger, M.; Metaxas, A.; Balbar, A.C.; McGowan, J.A.; Daigle, R.; Kuempel, C.D.; Treml, E.A.; Possingham, H.P. Demystifying ecological connectivity for actionable spatial conservation planning. Trends Ecol. Evol. 2022, 37, 1079–1091. [Google Scholar] [CrossRef]
- Liu, B.; Gao, H.; Wang, J.; Zhu, Z.; Qian, C.; Fan, Z.; He, K. Migratory Connectivity of Zhejiang, with a Critical Stopover in East Asian-Australasian Flyway, Based on Recovery Data. Animals 2024, 14, 2404. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, J.; Zeng, G.; Xu, J.; Sang, L.; Liu, Q.; Dai, J.; Xiong, W.; Yuan, Z.; Wang, Y.; et al. Effects of Early Dry Season on Habitat Suitability for Migratory Birds in China’s Two Largest Freshwater Lake Wetlands after the Impoundment of Three Gorges Dam. J. Environ. Inform. 2019, 36, 82–92. [Google Scholar] [CrossRef]
- Modica, G.; Praticò, S.; Laudari, L.; Ledda, A.; Di Fazio, S.; De Montis, A. Implementation of multispecies ecological networks at the regional scale: Analysis and multi-temporal assessment. J. Environ. Manag. 2021, 289, 112494. [Google Scholar] [CrossRef]
- Salgueiro, P.A.; Valerio, F.; Silva, C.; Mira, A.; Rabaça, J.E.; Santos, S.M. Multispecies landscape functional connectivity enhances local bird species’ diversity in a highly fragmented landscape. J. Environ. Manag. 2021, 284, 112066. [Google Scholar] [CrossRef]
- Utama, A.; Nurvianto, S. The building size effect on bird community assemblages in tropical urban ecosystem. Biodiversitas J. Biol. Divers. 2022, 23, 5. [Google Scholar] [CrossRef]
- Villegas, M.; Garitano-Zavala, Á. Bird community responses to different urban conditions in La Paz, Bolivia. Urban Ecosyst. 2010, 13, 375–391. [Google Scholar] [CrossRef]
- Yang, Y.; Zhou, Y.; Feng, Z.; Wu, K. Making the Case for Parks: Construction of an Ecological Network of Urban Parks Based on Birds. Land 2022, 11, 1144. [Google Scholar] [CrossRef]
- Fei, F.; Yin, H.; Kong, F.; Chen, J.; Liu, J.; Song, X. Comparative analysis of ecological network pattern using 2D and 3D information in main urban area of Nanjing, China. Acta Ecol. Sin. 2020, 40, 5534–5545. [Google Scholar]
- Hou, W.; Zhou, W.; Li, J.; Li, C. Simulation of the potential impact of urban expansion on regional ecological corridors: A case study of Taiyuan, China. Sustain. Cities Soc. 2022, 83, 103933. [Google Scholar] [CrossRef]
- Li, P.; Cao, H.; Sun, W.; Chen, X. Quantitative evaluation of the rebuilding costs of ecological corridors in a highly urbanized city: The perspective of land use adjustment. Ecol. Indic. 2022, 141, 109130. [Google Scholar] [CrossRef]
- Sun, X.; Long, Z.; Jia, J. A multi-scale Maxent approach to model habitat suitability for the giant pandas in the Qionglai mountain, China. Glob. Ecol. Conserv. 2021, 30, e01766. [Google Scholar] [CrossRef]
- Kim, B.; Park, J. Random ecological networks that depend on ephemeral wetland complexes. Ecol. Eng. 2020, 156, 105972. [Google Scholar] [CrossRef]
- Chan, M.; Tsang, T.P.N.; Dingle, C.; Early, R.; Sorte, C.J.B.; Bonebrake, T.C. Microhabitat coverage influences avian species composition more than habitat heterogeneity in Hong Kong urban parks. Urban For. Urban Green. 2024, 101, 128519. [Google Scholar] [CrossRef]
- Kim, B.; Lee, J.; Park, J. Role of small wetlands on the regime shift of ecological network in a wetlandscape. Environ. Res. Commun. 2022, 4, 041006. [Google Scholar] [CrossRef]
- Li, Z.; Liu, Y.; Zeng, H. Application of the MaxEnt model in improving the accuracy of ecological red line identification: A case study of Zhanjiang, China. Ecol. Indic. 2022, 137, 108767. [Google Scholar] [CrossRef]
- Blazquez-Cabrera, S.; Gastón, A.; Beier, P.; Garrote, G.; Simón, M.Á.; Saura, S. Influence of separating home range and dispersal movements on characterizing corridors and effective distances. Landsc. Ecol. 2016, 31, 2355–2366. [Google Scholar] [CrossRef]
- Xing, Y.; Lu, J.; Chen, S.; Dissanayake, S. Vulnerability analysis of urban rail transit based on complex network theory: A case study of Shanghai Metro. Public Transp. 2017, 9, 501–525. [Google Scholar] [CrossRef]
- Li, M.; Liu, R.R.; Lü, L.Y.; Hu, M.B.; Xu, S.Q.; Zhang, Y.C. Percolation on complex networks: Theory and application. Phys. Rep. 2021, 907, 1–68. [Google Scholar] [CrossRef]
- Dang, H.; Bai, J.; Lü, Y.; Li, J. The cascade failure model under ecological network is effective for quantifying the resilience of urban regions. Sustain. Cities Soc. 2024, 114, 105749. [Google Scholar] [CrossRef]
- Xia, Y.; Wang, C.; Shen, H.-L.; Song, H. Cascading failures in spatial complex networks. Phys. A Stat. Mech. Appl. 2020, 559, 125071. [Google Scholar] [CrossRef]
- Gao, C.; Pan, H.; Wang, M.; Zhang, T.; He, Y.; Cheng, J.; Yao, C. Identifying priority areas for ecological conservation and restoration based on circuit theory and dynamic weighted complex network: A case study of the Sichuan Basin. Ecol. Indic. 2023, 155, 111064. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, Q.; Xie, M.; Shibata, S. Construction feasibility evaluation for potential ecological corridors under different widths: A case study of Chengdu in China. Landsc. Ecol. Eng. 2023, 19, 381–399. [Google Scholar] [CrossRef]
- Zhang, K.; Pan, J. Evaluation of ecological network resilience using OWA and attack scenario simulation in the Gansu section of the Yellow River Basin, NW China. Environ. Res. Commun. 2024, 6, 8. [Google Scholar] [CrossRef]
- Sasaki, T.; Furukawa, T.; Iwasaki, Y.; Seto, M.; Mori, A.S. Perspectives for ecosystem management based on ecosystem resilience and ecological thresholds against multiple and stochastic disturbances. Ecol. Indic. 2015, 57, 395–408. [Google Scholar] [CrossRef]
- Lu, Y.; Liu, Y.; Xing, L.; Liu, Y. Robustness test of multiple protection strategies for ecological networks from the perspective of complex networks: Evidence from Wuhan Metropolitan Area, China. Land Degrad. Dev. 2023, 34, 52–71. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, M.; Zhao, D.; Duo, L.; Lu, C. Optimizing the ecological network of resource-based cities to enhance the resilience of regional ecological networks. Environ. Sci. Pollut. Res. 2024, 31, 17182–17205. [Google Scholar] [CrossRef] [PubMed]
- Calderón-Contreras, R.; Quiroz-Rosas, L.E. Analysing scale, quality and diversity of green infrastructure and the provision of Urban Ecosystem Services: A case from Mexico City. Ecosyst. Serv. 2017, 23, 127–137. [Google Scholar] [CrossRef]
- Callaghan, C.T.; Bino, G.; Major, R.E.; Martin, J.M.; Lyons, M.B.; Kingsford, R.T. Heterogeneous urban green areas are bird diversity hotspots: Insights using continental-scale citizen science data. Landsc. Ecol. 2019, 34, 1231–1246. [Google Scholar] [CrossRef]
- Guo, F.; Buler, J.J.; Smolinsky, J.A.; Wilcove, D.S. Autumn stopover hotspots and multiscale habitat associations of migratory landbirds in the eastern United States. Proc. Natl. Acad. Sci. USA 2023, 120, e2203511120. [Google Scholar] [CrossRef]
- Walker, J.; Therrien, J.-F.; Bégin-Marchand, C.; Côté, P.; Terrigeol, A.; Gagnon, F.; Tremblay, J.A. Insights from a decade of using the Motus network to track boreal bird species from Observatoire d’oiseaux de Tadoussac, Québec to temperate and tropical wintering grounds. J. Field Ornithol. 2025, 96, 2. [Google Scholar] [CrossRef]
- Taylor, C.M.; Laughlin, A.J.; Hall, R.J. The response of migratory populations to phenological change: A Migratory Flow Network modelling approach. J. Anim. Ecol. 2016, 85, 648–659. [Google Scholar] [CrossRef] [PubMed]
- Kudari, M.; Nandeppanavar, A.S.; Thotad, P.; Jatti, S.; Koti, S. A Machine Learning based approach for Bird Migration Detection and feature analysis with SHAP. In Proceedings of the 2024 IEEE North Karnataka Subsection Flagship International Conference (NKCon), Bagalkote, India, 21–22 September 2024; pp. 1–7. [Google Scholar]
- Haase, P.; Tonkin, J.D.; Stoll, S.; Burkhard, B.; Frenzel, M.; Geijzendorffer, I.R.; Häuser, C.; Klotz, S.; Kühn, I.; McDowell, W.H.; et al. The next generation of site-based long-term ecological monitoring: Linking essential biodiversity variables and ecosystem integrity. Sci. Total Environ. 2018, 613–614, 1376–1384. [Google Scholar] [CrossRef] [PubMed]
- Marrotte, R.R.; Bowman, J.; Brown, M.G.C.; Cordes, C.; Morris, K.Y.; Prentice, M.B.; Wilson, P.J. Multi-species genetic connectivity in a terrestrial habitat network. Mov. Ecol. 2017, 5, 21. [Google Scholar] [CrossRef] [PubMed]
- Geng, J.; Cheng, C.; Shen, S.; Dai, K.; Zhang, T. STAPLE: A land use/-cover change model concerning spatiotemporal dependency and properties related to landscape evolution. Environ. Model. Softw. 2024, 178, 106059. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Description | Data Type | Resolution | Source |
|---|---|---|---|---|
| Bird point data | - | Excel | - | https://www.eBird.org; https://www.gbif.org |
| Land use | CLCD | Raster | 30 m | https://doi.org/10.5281/zenodo.12779975 |
| Normalized Difference Vegetation Index | NDVI | Raster | 30 m | Google Earth Engine (GEE) |
| Fractional Vegetation Cover | FVC | Raster | 30 m | https://doi.org/10.11888/Terre.tpdc.300590 |
| Digital Elevation Model | DEM | Raster | 30 m | https://www.gscloud.cn/ |
| Slope | - | Raster | 30 m | Calculate in ArcGIS Pro |
| Aspect | - | Raster | 30 m | Calculate in ArcGIS Pro |
| Road | - | Shape | - | http://www.openstreetmap.org/ |
| Building height | - | Shape | - | https://doi.org/10.5281/zenodo.11397015 |
| Resistance Factor | Classification | Resistance Value | Weight | Resistance Factor | Classification | Resistance Value | Weight |
|---|---|---|---|---|---|---|---|
| Distance to Road | >1500 m | 5 | 0.3523 | CLCD | Forest | 5 | 0.1449 |
| 900–1500 m | 10 | Grassland | 5 | ||||
| 600–900 m | 20 | water | 10 | ||||
| 300–600 m | 50 | Cropland | 20 | ||||
| <300 m | 100 | Impervious Surface | 100 | ||||
| NDVI | 0.9–1.0 | 5 | 0.2516 | Building height | <20 m | 20 | 0.2437 |
| 0.8–0.9 | 10 | 20–50 m | 40 | ||||
| 0.7–0.8 | 20 | 50–100 m | 60 | ||||
| 0.6–0.7 | 30 | 100–200 m | 80 | ||||
| 0.5–0.6 | 40 | >200 m | 100 | ||||
| 0.4–0.5 | 50 | DEM | <100 m | 5 | 0.0075 | ||
| 0.3–0.4 | 60 | 100–300 m | 20 | ||||
| 0.2–0.3 | 70 | >300 m | 50 | ||||
| 0.1–0.2 | 80 | ||||||
| 0.0–0.1 | 90 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tong, A.; Ouyang, H.; Zhou, Y.; Li, Z. Multidimensional Bird Habitat Network Resilience Assessment and Ecological Strategic Space Identification in International Wetland City. Land 2025, 14, 1166. https://doi.org/10.3390/land14061166
Tong A, Ouyang H, Zhou Y, Li Z. Multidimensional Bird Habitat Network Resilience Assessment and Ecological Strategic Space Identification in International Wetland City. Land. 2025; 14(6):1166. https://doi.org/10.3390/land14061166
Chicago/Turabian StyleTong, An, Huizi Ouyang, Yan Zhou, and Ziyan Li. 2025. "Multidimensional Bird Habitat Network Resilience Assessment and Ecological Strategic Space Identification in International Wetland City" Land 14, no. 6: 1166. https://doi.org/10.3390/land14061166
APA StyleTong, A., Ouyang, H., Zhou, Y., & Li, Z. (2025). Multidimensional Bird Habitat Network Resilience Assessment and Ecological Strategic Space Identification in International Wetland City. Land, 14(6), 1166. https://doi.org/10.3390/land14061166