1. Introduction
The potential for wildfire ignition “based on flammability and exposure to ignition vectors” [
1] has been increasing in fire-prone regions globally throughout the past century [
2], and the world is seeing larger, more frequent wildfires as a result [
3], necessitating restoration plans that incorporate fire resiliency. There are multiple causes of the global increase in wildfires, including climate change [
4], changes in land use patterns [
1,
5], and shifting fuel dynamics within ecosystems, which occur when an ecosystem’s vegetation changes in a way that affects its fire potential [
6]. Shifts in fuel dynamics may be precipitated seasonally with senescence or over multiple years via structural changes as vegetation ages [
7,
8]. Shifts can also be caused by human alterations of an ecosystem’s structure such as logging, development, fire suppression [
1,
9,
10,
11], or when an ecosystem is invaded by a plant species such as non-native, invasive grasses [
6,
12,
13].
Invasive grasses can disrupt soil nutrient dynamics [
14,
15], plant water dynamics [
16], and soil carbon cycling [
17]. Many are suited to post-disturbance environments and propagate easily after wildfire, and some of the most successful invasive grasses have seeds that are stimulated to germinate after exposure to heat and smoke [
18,
19]. Additionally, many invasive grasses tend to have low moisture levels and high fuel biomass and flammability [
6,
12,
13]. Invasive species that are fire-tolerant can crowd out native species that are slower to establish in a post-fire landscape, creating a positive feedback loop in which an increase in the abundance of invasive species leads to more frequent wildfire, which in turn leads to a further increase in the abundance of invasive species [
20]. Such positive feedback increases the resilience of the ecosystem’s invaded state [
21], and makes the restoration of diverse, native landscapes without invasive grasses difficult [
22,
23].
The Hawaiian Islands offer an example of this phenomenon, as native forests there are being encroached upon by highly flammable invasive grasses such as
Ehrharta stipoides (meadow ricegrass)
, Megathyrsus maximus (Guinea grass), and
Andropogon glomeratus (bushy bluestem) [
12,
24,
25]. These grasses were introduced to the Hawaiian Islands to provide forage for cattle and quickly began to reproduce, becoming widespread by the 1960s [
24,
26]. In the absence of grass invasion, Hawaiian forests and woodlands have historically had subcanopies dominated by ferns, shrubs, and sub-trees, with little to no native grass present [
24,
26]. Because invading grasses change the fuel composition of Hawaiian forests, propagate quickly in burned areas, and prevent the reestablishment of less flammable native species, they increase fire potential and have led to wildfires that are more frequent and intense than historical fire patterns [
24,
25].
Paleontological evidence indicates that pre-settlement Hawaiʻi experienced occasional wildfires caused by volcanic activity and lightning strikes [
27]. While there is some debate over the degree to which Polynesian settlers used fire to alter landscapes in Hawaiʻi [
28], sedimentological evidence does indicate that their arrival caused a significant increase in fire frequency [
27,
29,
30]. Early Hawaiians employed fire as a tool to increase the abundance of
Heteropogon contortus (pili grass), which they used for thatching [
30]. European colonization further increased fire frequency as agriculture expanded and non-native grasses were introduced for cattle forage [
27,
30]. From 1904 to 1959, the total area burned per year statewide increased over fourfold, and it is experiencing unprecedented highs in the present day [
31]. From 2005 to 2011, there were on average 1007 fires per year across all the islands, burning an average of 8427 ha per year [
31]. Because many native plant species in Hawaiʻi are vulnerable to the effects of frequent [
32] or intense [
30] fire, the shifting Hawaiian wildfire regime has contributed to their decline, which in turn has negatively affected wildlife habitat quality and ecosystem function [
30].
Efforts to re-establish native species composition in grass-invaded sites in Hawaiʻi often include planting or otherwise facilitating the native koa tree
Acacia koa, which can reduce grass cover through shading [
33,
34].
A. koa is considered a good candidate for restoration of Hawaiian forests because it has high survival rates, grows quickly relative to other native canopy dominant trees, and holds economic, ecological, and cultural value [
33,
35]. Additionally, in the absence of invasive grass,
A. koa quickly regenerates after disturbance, including fire, through resprouting and seedling establishment [
36].
However, the overrepresentation of
A. koa at restoration sites may have negative ecological consequences.
A. koa is a nitrogen fixer, which leads to localized increase in invasive grass under its canopy relative to under the canopies of common non-fixing native trees such as
Metrosideros polymorpha (‘ōhi‘a) [
37]. This increased grass cover hinders the development of native understories, particularly in
A. koa monocultures [
37]. Because the National Park Service (NPS) and other management organizations aim to restore biodiversity in Hawaiʻi, suppression of native understory may be counterproductive, and restoration alternatives to
A. koa are being considered. Additionally, as invasive grass cover expands, grass-fueled fires will become more frequent and more likely to diminish
A. koa seed banks, because frequent fires can prevent
A. koa stands from reaching seeding age before being burned [
36], a phenomenon referred to as immaturity risk [
38]. It is unknown how a higher frequency of grass-fueled fires may affect post-fire
A. koa establishment in areas with increasing grass cover. Increased grass cover may suppress post-fire regeneration of
A. koa due to competition for resources other than nitrogen [
39].
If grass cover negatively affects post-fire
A. koa regeneration,
A. koa’s grass facilitation could be highly problematic for the long-term success of
A. koa restoration in areas that experience wildfires [
36]. Literature is sparse on the effects of high-severity fire on
A. koa regeneration, although there is evidence that
A. koa is able to establish naturally and aggressively after high-severity fire [
40]. It is not known whether there is a threshold at which grass cover has substantial negative effects on
A. koa regeneration through its potential influence on fire severity. We aimed to test this at sites with varying levels of pre-fire grass cover by determining the combined effects of grass cover and fire severity on
A. koa regeneration densities one year after fire.
Assessing grass cover is key to understanding its effects on ecosystems and can be done in person or remotely. However, it is difficult to assess grass cover from remote sensing data in forested ecosystems when the forest floor is obscured by the canopy, as is often the case in Hawaiʻi. This presents a challenge to determining the fire risk and management needs of forested landscapes without spending time and resources on in-person surveys. We aimed to test whether oblique-angle aerial images could fill this knowledge gap. Oblique aerial images are shot at an angle that allows the viewer to see further under the canopy than one can using traditional top-down images, and this could potentially allow for greater ability to assess grass cover.
The objectives of this study were to quantify how grass cover, fire severity, and the interaction of the two affect A. koa regeneration, and to discover whether one can obtain accurate grass cover estimations from oblique-angle aerial photography. The future resilience of A. koa to the increasing threat of grass-fueled wildfire depends on how it responds to various levels of grass cover and fire severity. Understanding this response and knowing whether oblique-angle aerial photography is a viable option for grass cover measurement will allow land managers to make informed choices about how they assess fire hazard and restoration priorities on their land.
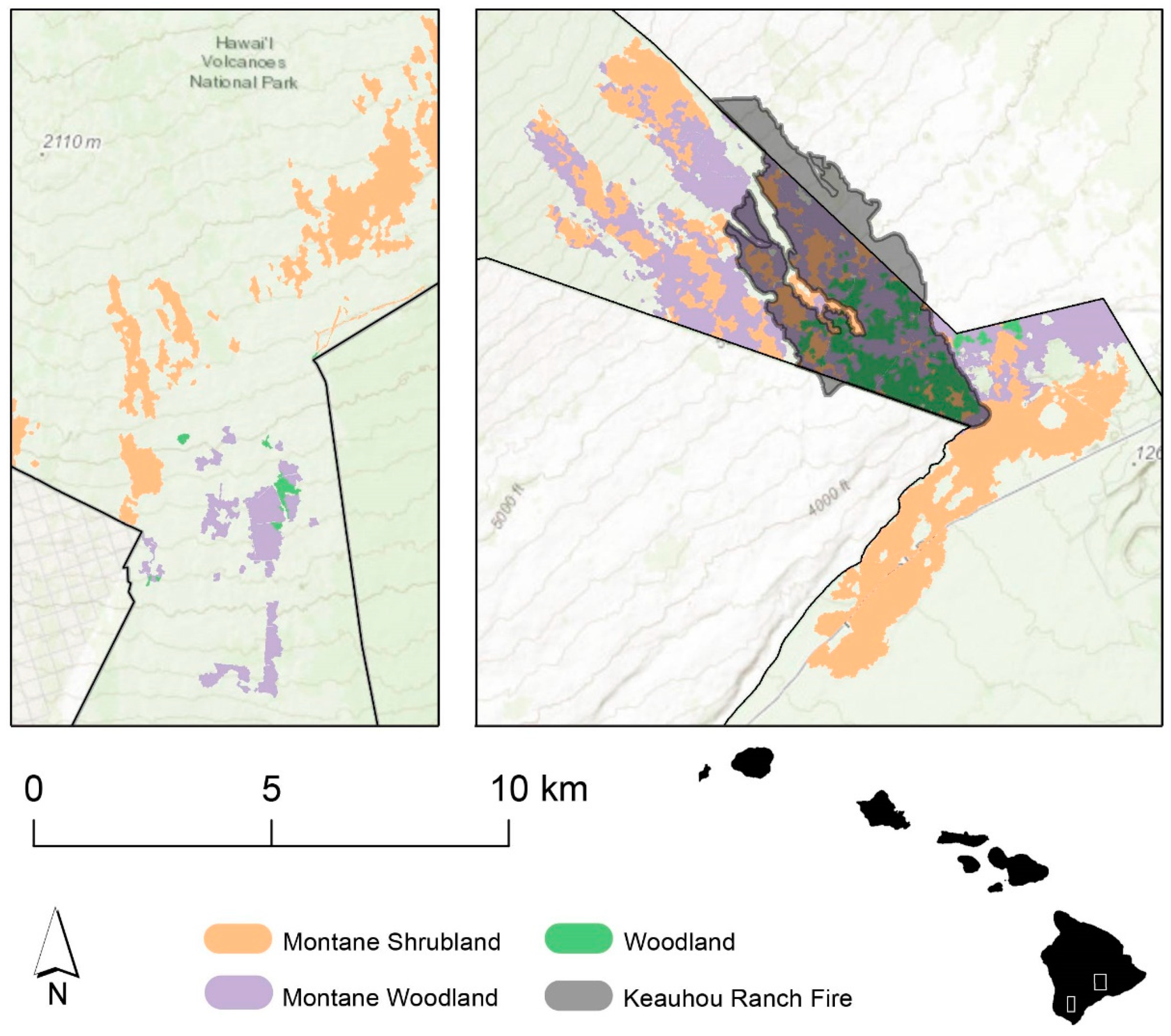
We chose to use the landscape burned in the 2018 Keauhou Ranch Fire as our study area. The Keauhou Ranch Fire burned an estimated 1203 hectares of land in Hawaiʻi Volcanoes National Park (HAVO) and 308 hectares of the surrounding state and public lands on the Island of Hawaiʻi [
41]. The fire affected numerous ecotypes, including
A. koa forests with varying levels of grass invasion. Ongoing restoration efforts in Hawaiʻi seek to restore forest habitat by planting native species and limiting invasive grass cover. However, for restoration efforts to be successful, land managers would benefit from knowledge of how invasive grass cover varies across the landscape and how it influences both fire effects and native plant response to fire. In an effort to achieve this knowledge, we investigated a method for estimating grass cover from oblique aerial photos, and analyzed the relationships between fire severity, grass cover, and the dominant native tree,
Acacia koa.
We asked the following:
At what level of precision can categories of pre-fire grass cover be accurately (≥60% accuracy) estimated from oblique aerial photos?
How do post-fire A. koa regeneration densities vary with fire severity?
How do post-fire A. koa regeneration densities vary with pre-fire grass cover and its interaction with fire severity?
5. Conclusions
In this study, we determined that grass cover can be accurately estimated from visual interpretation of oblique-angle aerial photos in Hawaiian montane shrubland and montane woodland ecosystems using four categories of precision. Other methods, such as lidar, may be helpful in areas with denser canopies.
We found that fire intensity, as indicated by minimum scorch height, has a positive effect on post-fire A. koa regeneration densities in mid-elevation woodlands when pre-fire tree density is low, though this trend reversed when pre-fire tree density was high (>70 trees per transect). Our findings imply that, when fire intervals are long enough to allow a sufficient soil seedbank to develop, A. koa is capable of regenerating at high density even after severe fire in low- to medium-density stands (<70 trees per transect). Further research would allow determination of the cause of the negative relationship between minimum scorch height and A. koa regeneration in high-density stands, and determination of how densely A. koa regenerates when fire intervals are less than 43 years, the shortest interval contained in our study area.
We did not detect a relationship between A. koa regeneration and the interaction between pre-fire grass cover and fire severity. This finding only applies to the montane woodland cover type because we could not use the grass cover estimations from the other two cover types in our analysis. If there truly is no relationship between grass cover, fire severity, and A. koa regeneration, then A. koa in grassy montane woodland areas are not at risk of grass suppressing their post-fire regeneration. Further research would be needed to determine whether this lack of a relationship holds true at larger sample sizes and across multiple cover types, and with other environmental variables considered.
Although
A. koa regenerates quickly after fire [
36,
40] and does not appear to be sensitive to potential changes in fire behavior caused by invasive grass, the establishment of
A. koa alone is not sufficient to maintain ecological diversity to the NPS’s standards, especially in areas where post-fire grass cover is high and suppresses native understory. Many threatened and endangered plant species of Hawaiʻi Volcanoes National Park are not fire adapted and cannot be planted into grassy areas; thus, creating landscapes that are low in grass cover and protected from wildfire may help with conservation efforts of these species. The planting of other native species alongside
A. koa, which is the NPS’s current practice in HAVO, would help achieve this goal in Hawaiʻi.




{kind=link}
{kind=link}
{kind=link}