
Timing of Tree Density Increases, Influence of Climate Change, and a Land Use Proxy for Tree Density Increases in the Eastern United States


Abstract
:1. Introduction
2. Materials and Methods
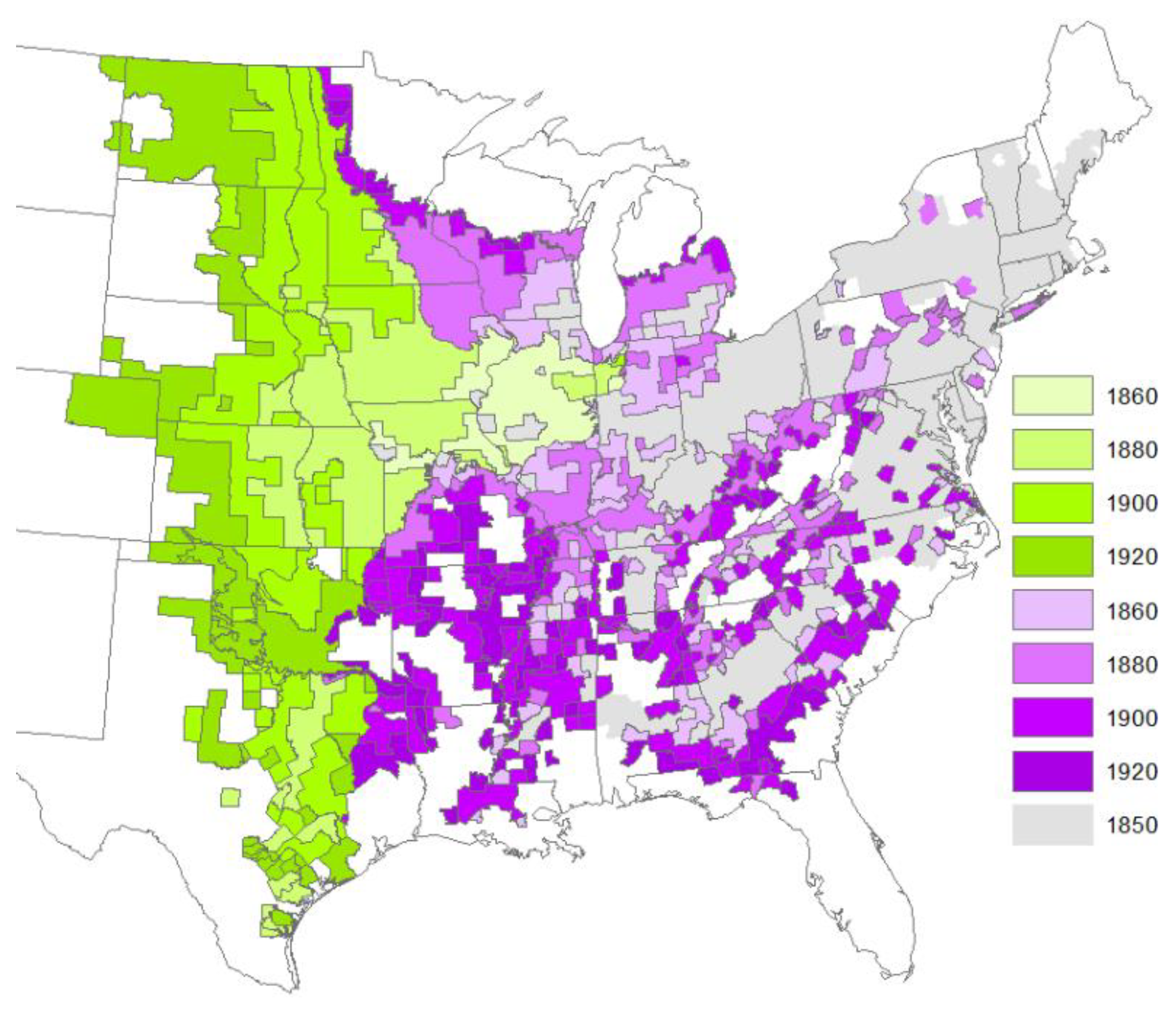
2.1. Timing of Tree Change from Historical Records
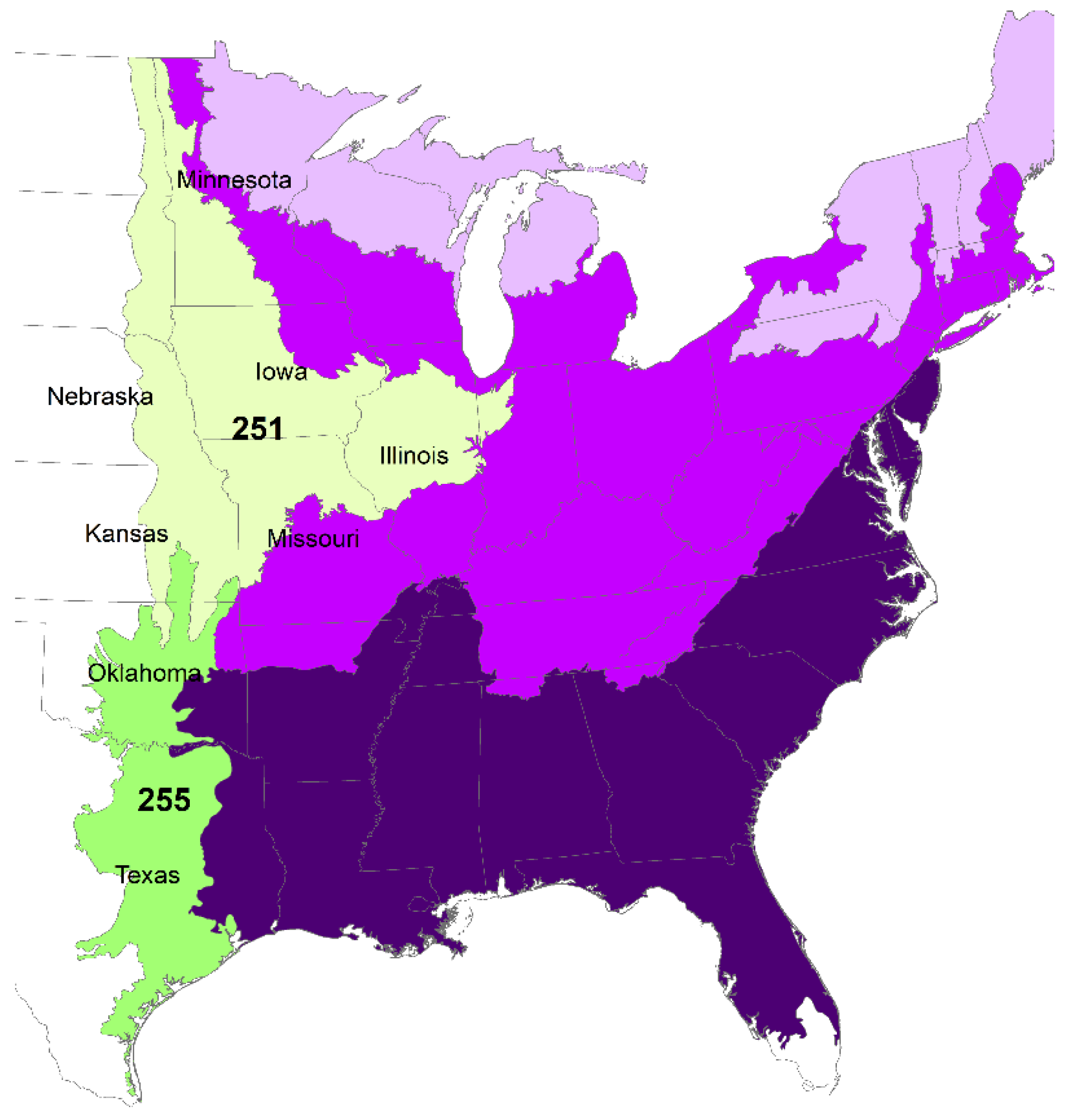
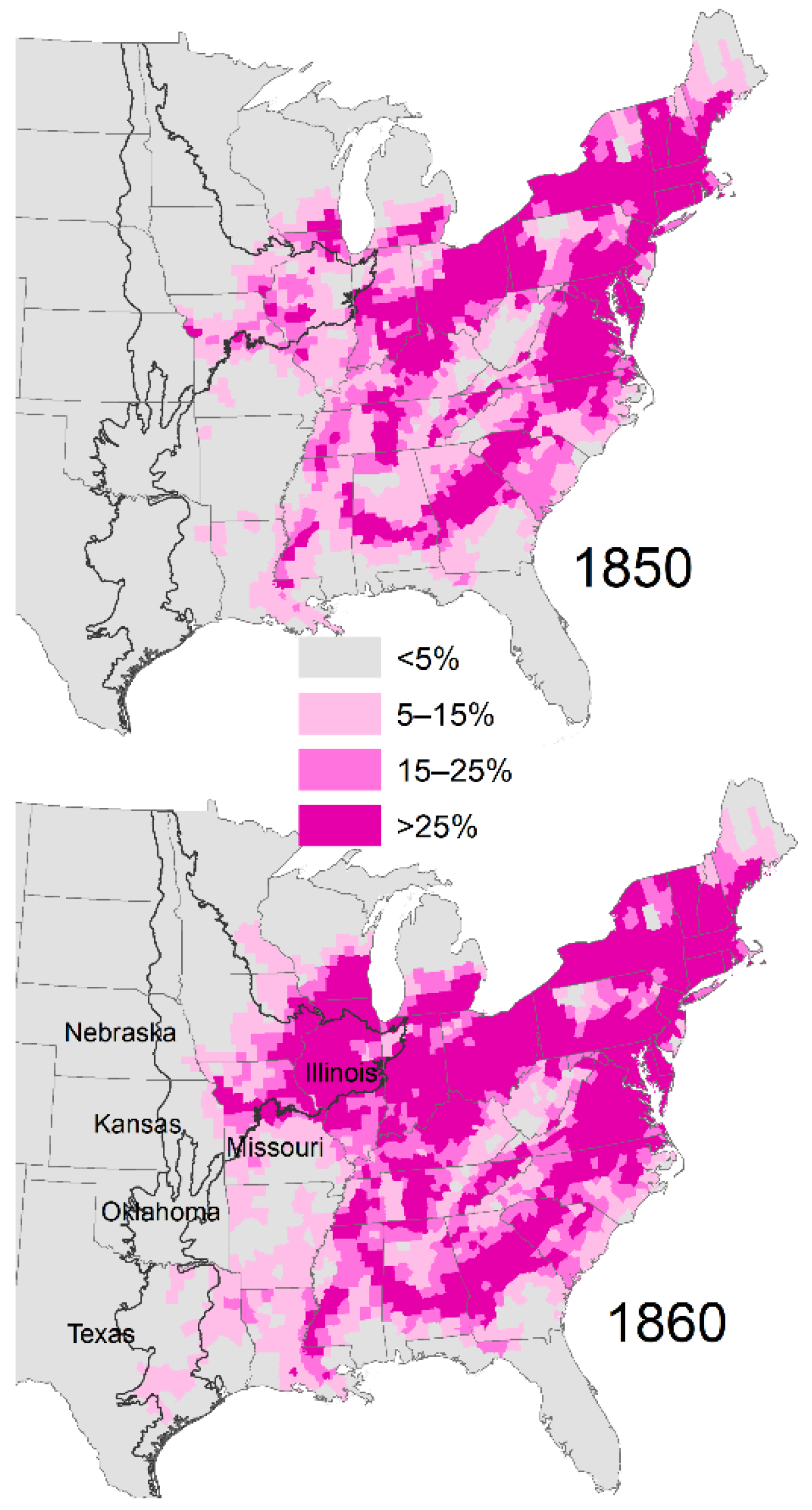
2.2. Decade That Agricultural Area Reached ≥25% by County as an Index of Land Use Change
2.3. The Palmer Modified Drought Index of Climate Change
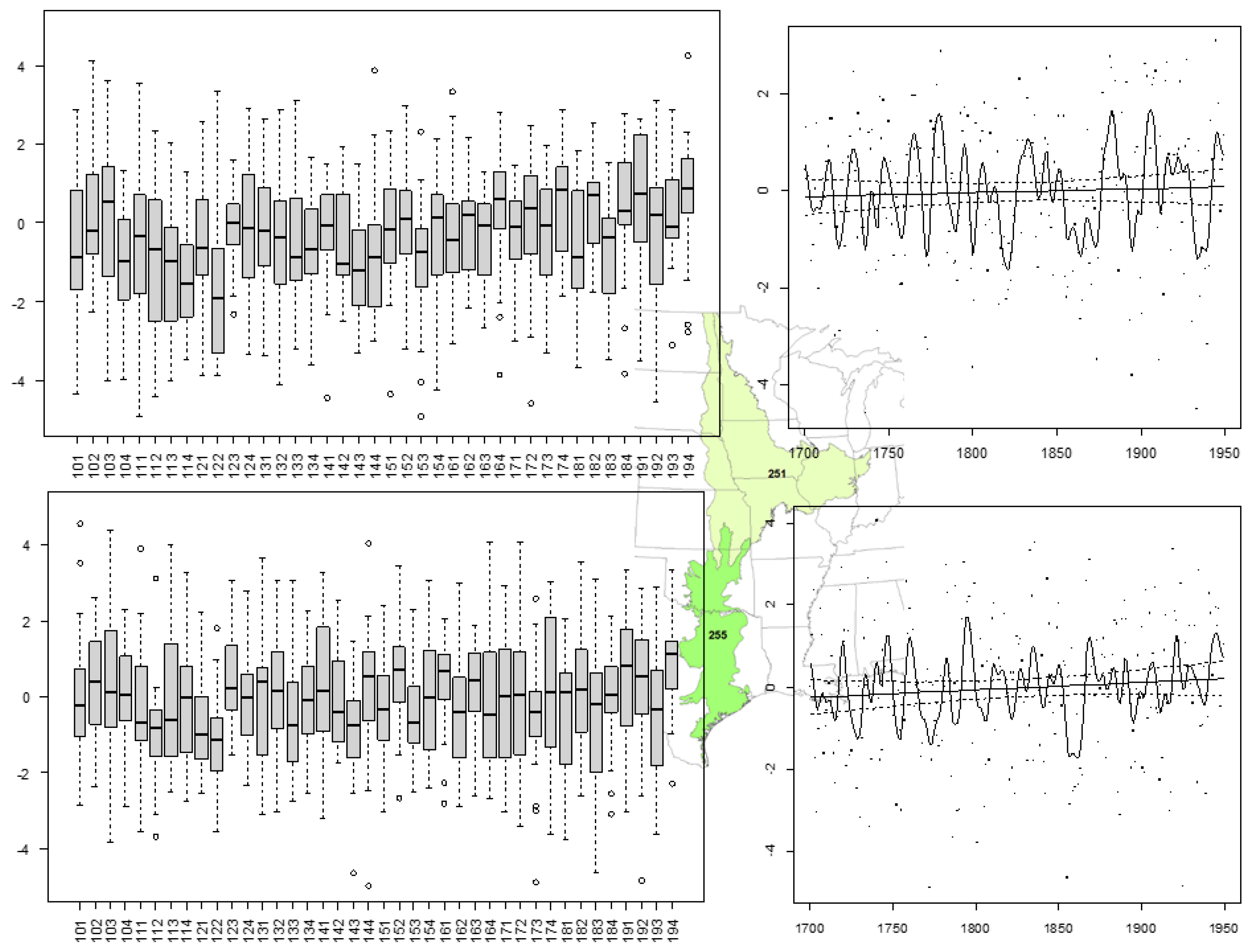
2.4. Change Point Detection
2.5. Change Point Detection for Historical Records of Tree Change in the Prairie Provinces
2.6. Change Point Detection for Grassland and Forest Landscapes Based on Land Use Change
3. Results
3.1. Timing of Tree Change from Historical Information
3.2. Climate Change Points Based on Historical Accounts of Timing
3.3. Climate Change Points Based on Timing of Land Use
4. Discussion
4.1. Influence of Climate Change
4.2. Influence of Land Use Change
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kangas, R.S.; Brown, T.J. Characteristics of US drought and pluvials from a high-resolution spatial dataset. Int. J. Climatol. 2007, 27, 1303–1325. [Google Scholar] [CrossRef]
- Fei, S.; Desprez, J.M.; Potter, K.M.; Jo, I.; Knott, J.A.; Oswalt, C.M. Divergence of species responses to climate change. Sci. Adv. 2017, 3, e1603055. [Google Scholar] [CrossRef] [Green Version]
- Hanberry, B.B.; Abrams, M.D.; White, J.D. Is increased precipitation during the 20th century statistically or ecologically significant in the eastern US? J. Land Use Sci. 2018, 13, 259–265. [Google Scholar] [CrossRef]
- Ziegler, S.S.; Larson, E.R.; Rauchfuss, J.; Elliott, G.P. Tree establishment during dry spells at an oak savanna in Minnesota. Tree-Ring Res. 2008, 64, 47–55. [Google Scholar] [CrossRef]
- Shuman, B.; Henderson, A.K.; Plank, C.; Stefanova, I.; Ziegler, S.S. Woodland-to-forest transition during prolonged drought in Minnesota after ca. AD 1300. Ecology 2009, 90, 2792–2807. [Google Scholar] [CrossRef] [Green Version]
- Rogers, T.R.; Russell, F.L. Historical patterns of oak population expansion in the Chautauqua Hills, Kansas. J. Biogeogr. 2014, 41, 2105–2114. [Google Scholar] [CrossRef]
- Delcourt, P.A.; Delcourt, H.R. Late-Quaternary dynamics of temperate forests: Applications of paleoecology to issues of global environmental change. Quat. Sci. Rev. 1987, 6, 129–146. [Google Scholar] [CrossRef]
- Hanberry, B.; Kabrick, J.M.; He, H.S. Changing tree composition by life history strategy in a grassland-forest landscape. Ecosphere 2014, 5, art34. [Google Scholar] [CrossRef] [Green Version]
- Lorimer, C.G. Historical and ecological roles of disturbance in eastern North American forest: 9,000 years of change. Wildl. Soc. Bull. 2001, 29, 425–439. [Google Scholar]
- Hanberry, B.B.; Coursey, K.; Kush, J.S. Structure and composition of historical longleaf pine ecosystems in Mississippi, USA. Hum. Ecol. 2018, 46, 241–248. [Google Scholar] [CrossRef]
- Hanberry, B.B. Baseline to novel ecosystems in Michigan, USA, with a quantitative and qualitative assessment. Écoscience 2020, 27, 255–268. [Google Scholar] [CrossRef]
- Pan, Y.; Chen, J.M.; Birdsey, R.; McCullough, K.; He, L.; Deng, F. Age structure and disturbance legacy of North American forests. Biogeosciences 2011, 8, 715–732. [Google Scholar] [CrossRef] [Green Version]
- Hanberry, B.B.; Hansen, M.H. Advancement of tree species across ecotonal borders into non-forested ecosystems. Acta Oecologica 2015, 68, 24–36. [Google Scholar] [CrossRef]
- Srinath, I.; Millington, A.C. Evaluating the Potential of the Original Texas Land Survey for Mapping Historical Land and Vegetation Cover. Land 2016, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Rothenberger, S.J. Extent of woody vegetation on the prairie in eastern Nebraska, 1855–1857. In Prairie Pioneers: Ecology, History and Culture: Proceedings of the Eleventh North American Prairie Conferences; Bragg, T.B., Stubbendieck, J., Eds.; University of Nebraska: Lincoln, NE, USA, 1989; pp. 15–18. [Google Scholar]
- McLachlan, J. Settlement Trees, Illinois Level 0 Ver 0. Environmental Data Initiative. 2020. Available online: https://portal.edirepository.org/nis/mapbrowse?packageid=msb-paleon.28.0 (accessed on 30 December 2020).
- Landers, R.Q. Native vegetation of Texas. Rangel. Arch. 1987, 9, 203–207. [Google Scholar]
- Hewes, F.W.; Gannett, H. Scribner’s Statistical Atlas of the United States. Scribner’s Sons: New York, NY, USA, 1883. Available online: https://www.loc.gov/item/a40001834/ (accessed on 7 January 2021).
- Cook, E.R.; Seager, R.; Heim, R.R.; Vose, R.S.; Herweijer, C.; Woodhouse, C. Megadroughts in North America: Placing IPCC projections of hydroclimatic change in a long-term paleoclimate context. J. Quat. Sci. 2010, 25, 48–61. Available online: https://www.ncdc.noaa.gov/paleo-search/study/19119 (accessed on 1 December 2020). [CrossRef] [Green Version]
- Keys, J.E.; Cleland, D.T.; McNab, W.H. Delineation, Peer Review, and Refinement of Subregions of the Conterminous United States; Gen. Tech. Report WO-76A; Department of Agriculture, Forest Service: Washington, DC, USA, 2007.
- Maizel, M.; White, R.D.; Root, R.; Gage, S.; Stitt, S.; Osborne, L.; Muehlbach, G. Historical Interrelationships between Population Settlement and Farmland in the Conterminous United States, 1790 to 1992. 1998. Available online: https://archive.usgs.gov/archive/sites/landcover.usgs.gov/luhna/chap2.html (accessed on 21 May 2020).
- Gleason, H.A. The vegetational history of the Middle West. Ann. Assoc. Am. Geogr. 1922, 12, 39–85. [Google Scholar] [CrossRef]
- Day, G.M. The Indian as an ecological factor in the Northeastern forest. Ecology 1953, 34, 329–346. [Google Scholar] [CrossRef]
- DeSantis, R.D.; Hallgren, S.W.; Lynch, T.B.; Burton, J.A.; Palmer, M.W. Long-term directional changes in upland Quercus forests throughout Oklahoma, USA. J. Veg. Sci. 2010, 21, 606–618. [Google Scholar] [CrossRef]
- Duncan, B.W.; Schmalzer, P.A. Anthropogenic influences on potential fire spread in a pyrogenic ecosystem of Florida, USA. Landsc. Ecol. 2004, 19, 153–165. [Google Scholar] [CrossRef]
- Marine Data Science. Comparison of Change Point Detection Methods. 2019. Available online: https://www.marinedatascience.co/blog/2019/09/28/comparison-of-change-point-detection-methods/ (accessed on 25 September 2021).
- Killick, R.; Eckley, I. Changepoint: An R package for changepoint analysis. J. Stat. Softw. 2014, 58, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Ross, G.J. Parametric and nonparametric sequential change detection in R: The cpm Package. J. Stat. Softw. 2015, 66, 1–19. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Davidson, R. An Excursion to the Mammoth Cave and the Barrens of Kentucky. A.T. Silliman and Son: Lexington, Kentucky, 1840. Available online: https://www.google.com/books/edition/An_Excursion_to_the_Mammoth_Cave_and_the/71EVAAAAYAAJ?hl=en&gbpv=1&printsec=frontcover (accessed on 1 July 2021).
- Shaler, N.S. Kentucky: A Pioneer Commonwealth. Houghton, Mifflin and Co.: Boston, MA, USA; New York, NY, USA, 1885; Available online: https://www.google.com/books/edition/Kentucky_a_Pioneer_Commonwealth/eIAxAQAAMAAJ?hl=en&gbpv=1&printsec=frontcover (accessed on 2 June 2021).
- Bragg, T.B.; Hulbert, L.C. Woody plant invasion of unburned Kansas bluestem prairie. J. Range Manag. 1976, 29, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Taylor, B. Colorado, a Summer Trip. Putman: New York, NY, USA, 1867. Available online: https://babel.hathitrust.org/cgi/pt?id=hvd.32044072260284&view=1up&seq=9 (accessed on 7 January 2021).
- Bruner, W.E. The vegetation of Oklahoma. Ecol. Monogr. 1931, 1, 100–188. [Google Scholar] [CrossRef]
- Gregg, J. Commerce of the Prairies. 1844. Available online: https://atlas.nmhum.org/pdfs/CommerceofthePrairies.pdf (accessed on 24 September 2021).
- Cook, O.F. Change of Vegetation on the South Texas Prairie; USDA, Bureau of Plant Industry Circular: Washington, DC, USA, 1908; Volume 14. Available online: https://archive.org/details/changeofvegetati14cook/page/n1/mode/2up (accessed on 7 January 2021).
- Wells, P.V. Scarp woodlands, transported grassland soils, and concept of grassland climate in the Great Plains region. Science 1965, 148, 246–249. [Google Scholar] [CrossRef]
- Hall, S.A. Deteriorated pollen grains and the interpretation of Quaternary pollen diagrams. Rev. Palaeobot. Palynol. 1981, 32, 193–206. [Google Scholar] [CrossRef]
- Fye, F.K.; Stahle, D.W.; Cook, E.R. Paleoclimatic analogs to twentieth-century moisture regimes across the United States. Bull. Am. Meteorol. Soc. 2003, 84, 901–910. [Google Scholar] [CrossRef]
- Stambaugh, M.C.; Guyette, R.P.; Marschall, J. Fire history in the Cherokee nation of Oklahoma. Hum. Ecol. 2013, 41, 749–758. [Google Scholar] [CrossRef]
- Gleason, H.A. Further views on the succession-concept. Ecology 1927, 8, 299–326. [Google Scholar] [CrossRef]
- Prism Climate Group. PRISM Climate Data. 2020. Available online: https://prism.oregonstate.edu/ (accessed on 30 December 2020).
- Loehle, C. Height growth rate tradeoffs determine northern and southern range limits for trees. J. Biogeogr. 1998, 25, 735–742. [Google Scholar] [CrossRef]
- McNulty, S.G.; Boggs, J.L.; Sun, G. The rise of the mediocre forest: Why chronically stressed trees may better survive extreme episodic climate variability. New For. 2014, 4, 403–415. [Google Scholar] [CrossRef]
- National Oceanic and Atmospheric Administration National Centers for Environmental Information (NCEI). U.S. Billion-Dollar Weather and Climate Disasters. 2020. Available online: https://www.ncdc.noaa.gov/billions/ (accessed on 25 February 2020).
- Niinemets, Ü.; Valladares, F. Tolerance to shade, drought, and waterlogging of temperate northern hemisphere trees and shrubs. Ecol. Monogr. 2006, 76, 521–547. [Google Scholar] [CrossRef]
- Hanberry, B.B. Recent shifts in shade tolerance and disturbance traits in forests of the eastern United States. Ecol. Process 2019, 8, 32. [Google Scholar] [CrossRef] [Green Version]
- Hanberry, B.B.; Bragg, D.C.; Hutchinson, T.F. A reconceptualization of open oak and pine ecosystems of eastern North America using a forest structure spectrum. Ecosphere 2018, 9, e02431. [Google Scholar] [CrossRef] [Green Version]
- Gresham, C.A.; Williams, T.M.; Lipscomb, D.J. Hurricane Hugo wind damage to southeastern US coastal forest tree species. Biotropica 1991, 23, 420–426. [Google Scholar] [CrossRef]
- Christy, M. Why are the prairies treeless? In Proceedings of the Royal Geographical Society and Monthly Record of Geography; Duke University Press: Durham, NC, USA, 1892; Volume 14, pp. 78–100. [Google Scholar]
- Gleason, H.A. The relation of forest distribution and prairie fires in the Middle West. Torreya 1913, 13, 173–181. [Google Scholar]
- Davis, A.M. The prairie-deciduous forest ecotone in the upper Middle West. Ann. Assoc. Am. Geogr. 1977, 67, 204–213. [Google Scholar] [CrossRef]
- Gerhard, F. Illinois as It Is. Keen and Lee: Chicago, IL, USA, 1857. Available online: https://catalog.hathitrust.org/Record/001268669 (accessed on 10 December 2020).
- Kettle, W.D.; Rich, P.M.; Kindscher, K.; Pittman, G.L.; Fu, P. Land-use history in ecosystem restoration: A 40-year study in the prairie-forest ecotone. Restor. Ecol. 2000, 8, 307–317. [Google Scholar] [CrossRef]
- Penfound, W.T. The savanna concept in Oklahoma. Ecology 1962, 43, 774–775. [Google Scholar] [CrossRef]
- Hanberry, B.B. Classifying large wildfires by land cover in the United States. Remote Sens. 2020, 12, 2966. [Google Scholar] [CrossRef]
- Hanberry, B.B. Reclassifying the wildland-urban interface using fire occurrences for the United States. Land 2020, 9, 225. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extent | Tree Change | PMDI Change |
|---|---|---|
| North prairie | 1860–1880 | 1840–1860 |
| South prairie | 1870–1890 | 1850–1870 |
| Grassland | 1860 (1850–1870) | 1830–1850 |
| 1880 (1870–1890) | 1850–1870 | |
| 1900 (1890–1910) | 1870–1890 | |
| 1920 (1910–1930) | 1890–1910 | |
| Forest | 1860 (1850–1870) | 1820–1850 |
| 1880 (1870–1890) | 1840–1870 | |
| 1900 (1890–1910) | 1860–1890 | |
| 1920 (1910–1930) | 1880–1910 |
| Extent | Location | Year | Reference |
|---|---|---|---|
| North prairie | |||
| Kansas | before 1867 | Taylor 1867 | |
| Illinois, Indiana, Iowa, Wisconsin | after 1860 | Gleason 1922 | |
| South prairie | |||
| Oklahoma | before 1931 | Bruner 1931 | |
| Oklahoma and south prairies | before 1844 | Grigg 1844 | |
| Texas | before 1878 to 1898 | Cook 1908 |
| Extent | Tree Change | PMDI Change | Change Point Years |
|---|---|---|---|
| North Prairie | 1860–1880 | 1840–1860 | |
| cpm | 1011 1036 1104 1108 1115 1233 1245 1276 1440 1490 1548 1619 1627 1636 1664 1672 1697 1700 1903 1931 1942 1953 1973 | ||
| changepoint | 1104 1108 1115 1233 1245 1619 1627 1993 | ||
| South prairie | 1870–1890 | 1850–1870 | |
| cpm | 1053 1061 1165 1177 1258 1455 1459 1494 1855 1865 1952 1957 | ||
| changepoint | 1053 1061 1168 1177 1257 1973 | ||
| Grassland | 1860 (1850–1870) | 1830–1850 | |
| cpm | 1104 1107 1245 1276 1736 1738 1799 1882 | ||
| changepoint | 1104 1107 1619 | ||
| 1880 (1870–1890) | 1850–1870 | ||
| cpm | 1011 1036 1104 1108 1115 1233 1245 1276 1440 1490 1492 1619 1627 1636 1664 1672 1697 1700 1736 1855 1876 1894 | ||
| changepoint | 1104 1108 1115 1233 1245 1440 1490 1492 1973 | ||
| 1900 (1890–1910) | 1870–1890 | ||
| cpm | 1011 1036 1076 1104 1233 1245 1276 1282 1455 1490 1576 1619 1626 1673 1697 1827 1838 1855 1877 1893 1902 1931 1942 1952 1965 1993 | ||
| changepoint | 1076 1233 1245 1992 | ||
| 1920 (1910–1930) | 1890–1910 | ||
| cpm | 1017 1040 1090 1146 1160 1233 1238 1265 1421 1425 1449 1498 1579 1592 1610 | ||
| changepoint | none | ||
| Forest | 1860 (1850–1870) | 1820–1850 | |
| cpm | 1021 1023 1104 1142 1154 1346 1351 1563 1574 1882 | ||
| changepoint | none | ||
| 1880 (1870–1890) | 1840–1870 | ||
| cpm | 1021 1023 1104 1265 1273 1346 1351 1555 1581 1736 1738 | ||
| changepoint | none | ||
| 1900 (1890–1910) | 1860–1890 | ||
| cpm | 1142 1150 1265 1269 1309 1346 1402 1450 1457 1556 1575 1903 | ||
| changepoint | none | ||
| 1920 (1910–1930) | 1880–1910 | ||
| cpm | 1035 1104 1142 1150 1179 1204 1207 1249 1309 1322 1450 1457 1488 1600 1608 1903 | ||
| changepoint | 1104 1600 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanberry, B.B. Timing of Tree Density Increases, Influence of Climate Change, and a Land Use Proxy for Tree Density Increases in the Eastern United States. Land 2021, 10, 1121. https://doi.org/10.3390/land10111121
Hanberry BB. Timing of Tree Density Increases, Influence of Climate Change, and a Land Use Proxy for Tree Density Increases in the Eastern United States. Land. 2021; 10(11):1121. https://doi.org/10.3390/land10111121
Chicago/Turabian StyleHanberry, Brice B. 2021. "Timing of Tree Density Increases, Influence of Climate Change, and a Land Use Proxy for Tree Density Increases in the Eastern United States" Land 10, no. 11: 1121. https://doi.org/10.3390/land10111121
APA StyleHanberry, B. B. (2021). Timing of Tree Density Increases, Influence of Climate Change, and a Land Use Proxy for Tree Density Increases in the Eastern United States. Land, 10(11), 1121. https://doi.org/10.3390/land10111121