
Environmental Factors Structuring Fish Communities in Floodplain Lakes of the Undisturbed System of the Biebrza River

 ,
, 
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
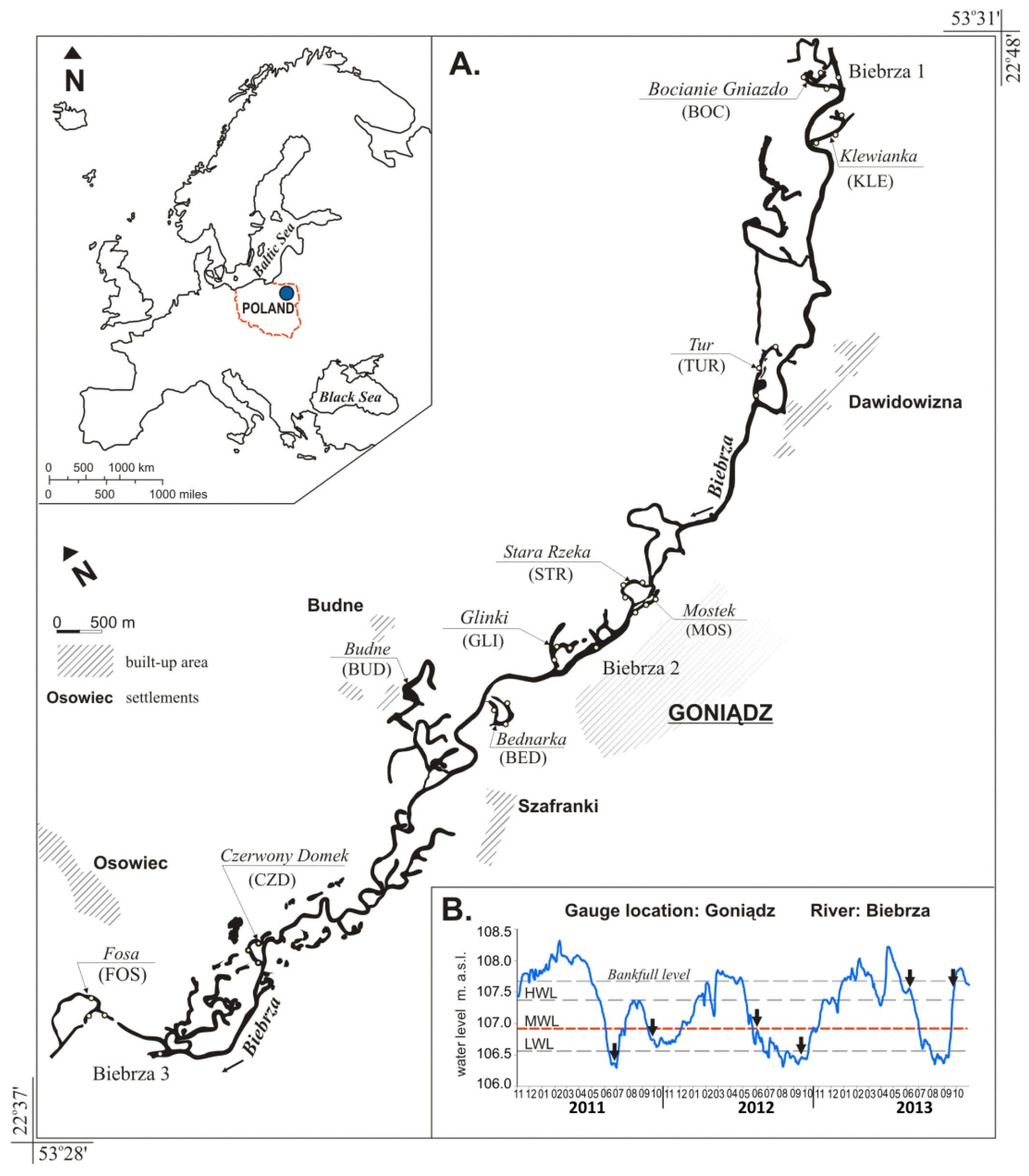
2.1. Study Area and Sampling Sites
2.2. Environmental Description of Study Sites
2.3. Environmental Data
2.4. Fish Sampling
2.5. Data Analysis
3. Results
3.1. Water Quality
3.2. Fish Assemblages
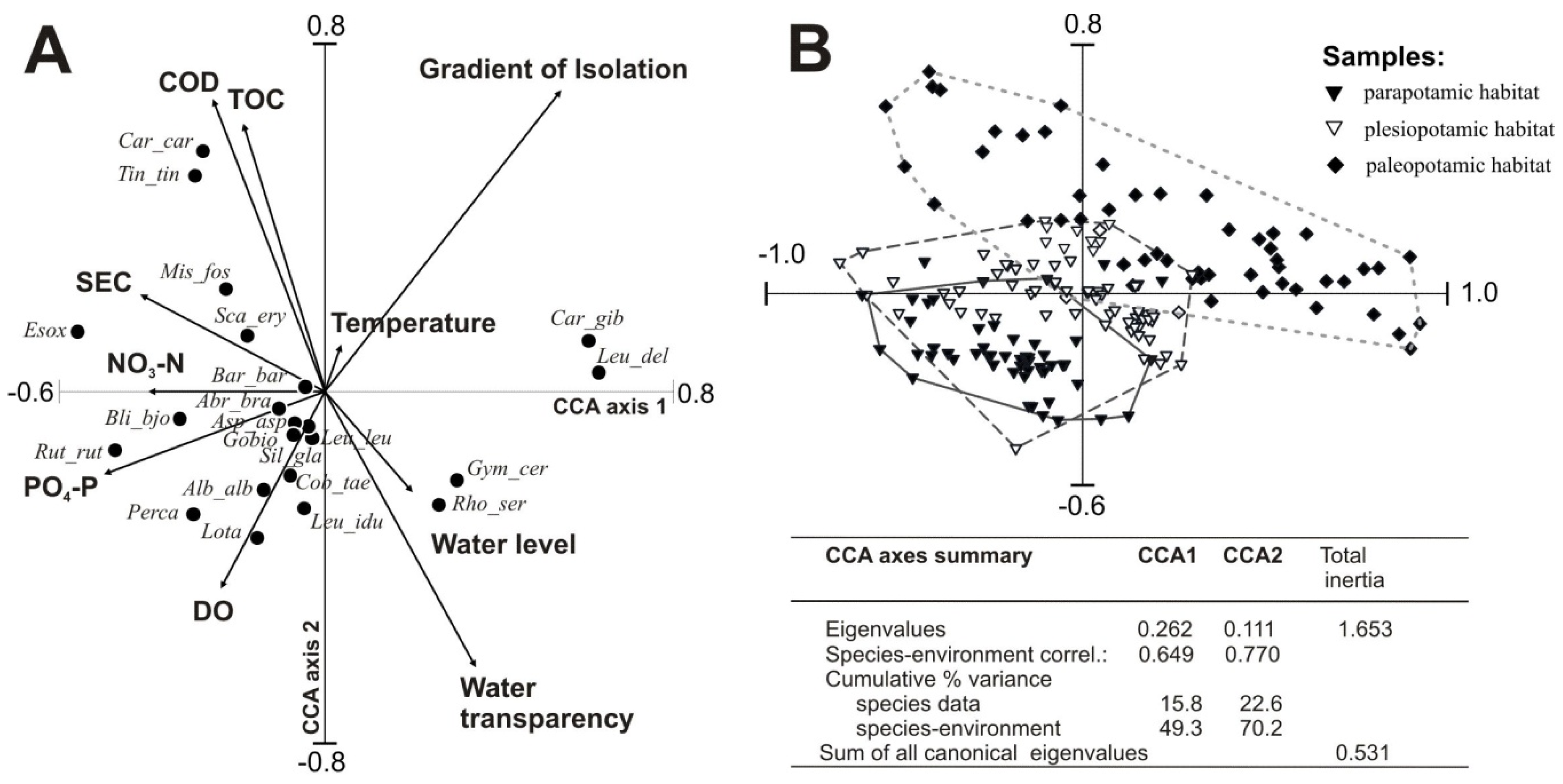
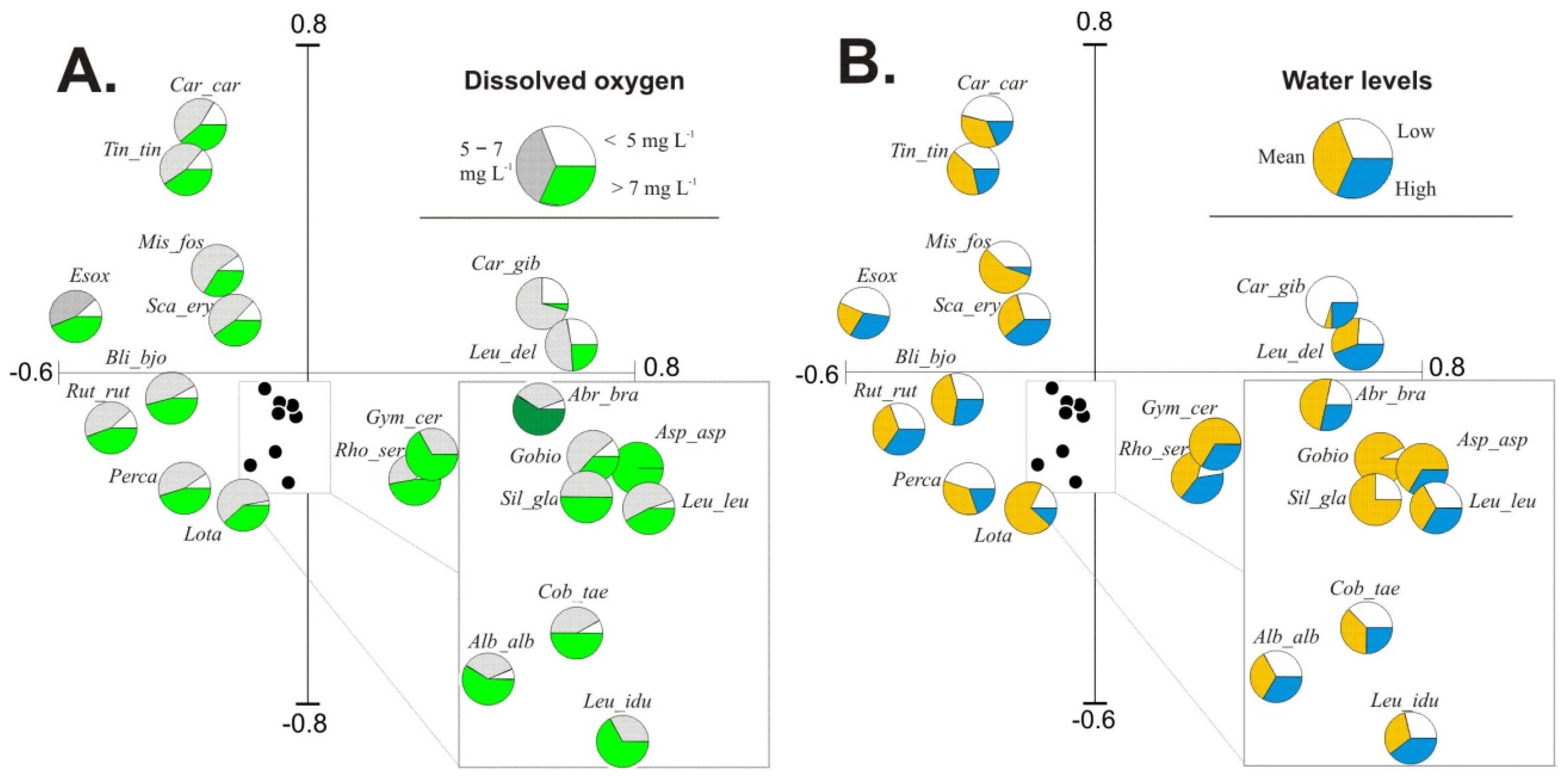
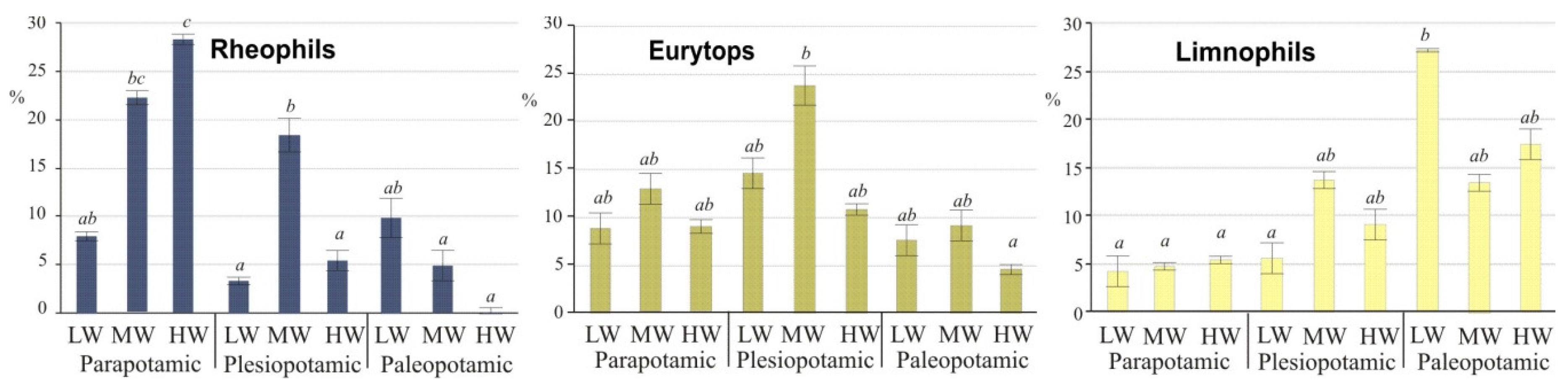
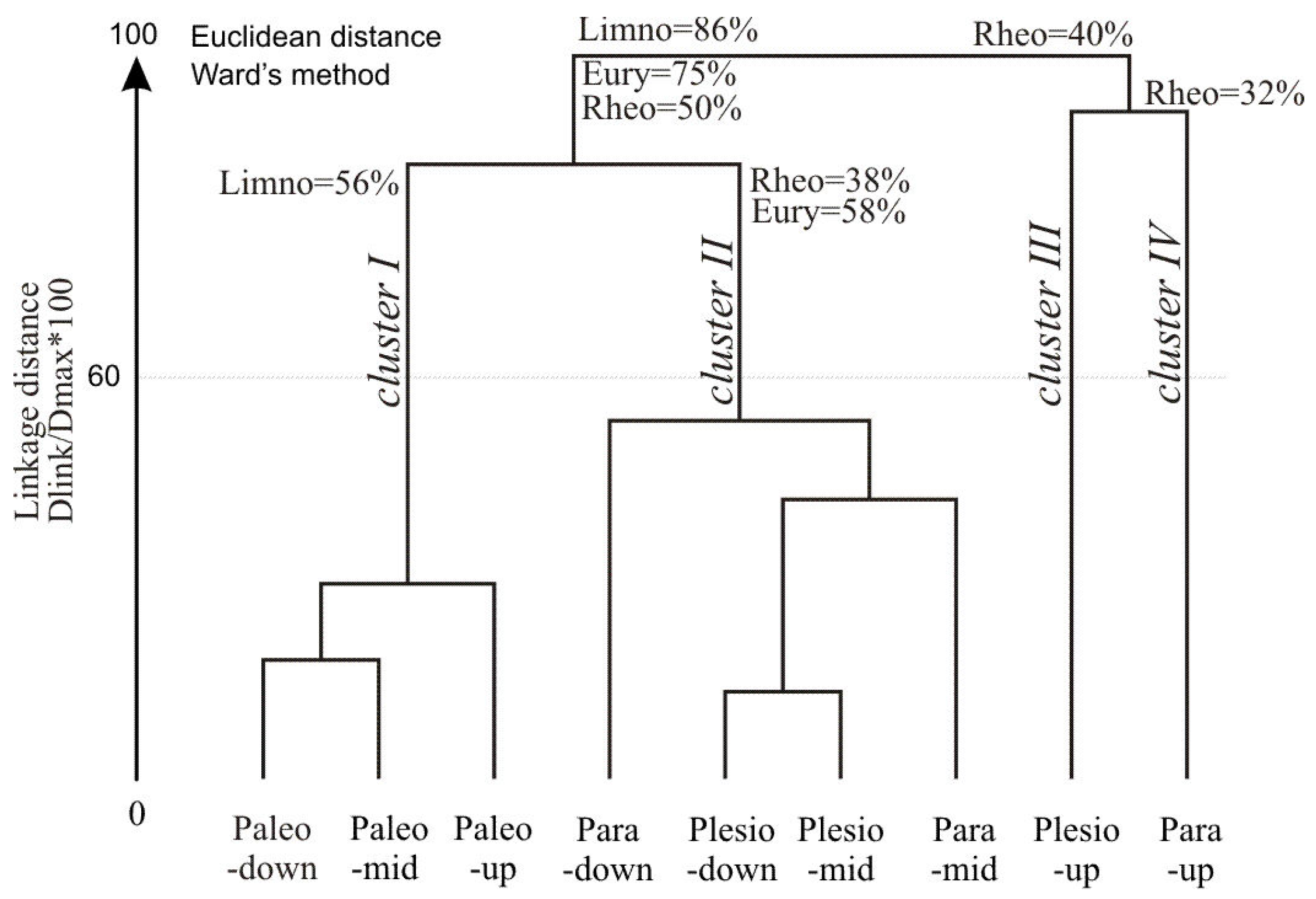
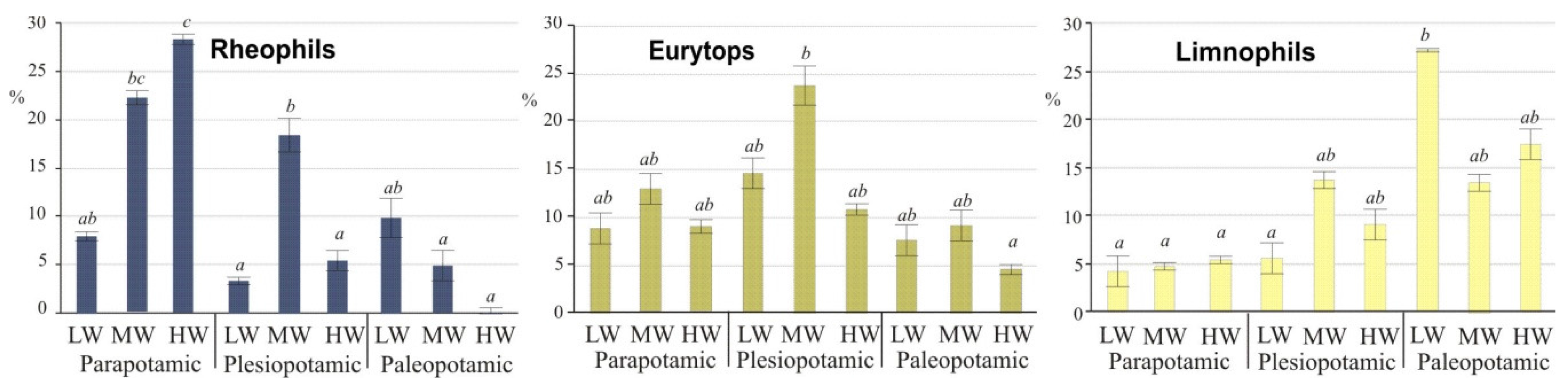
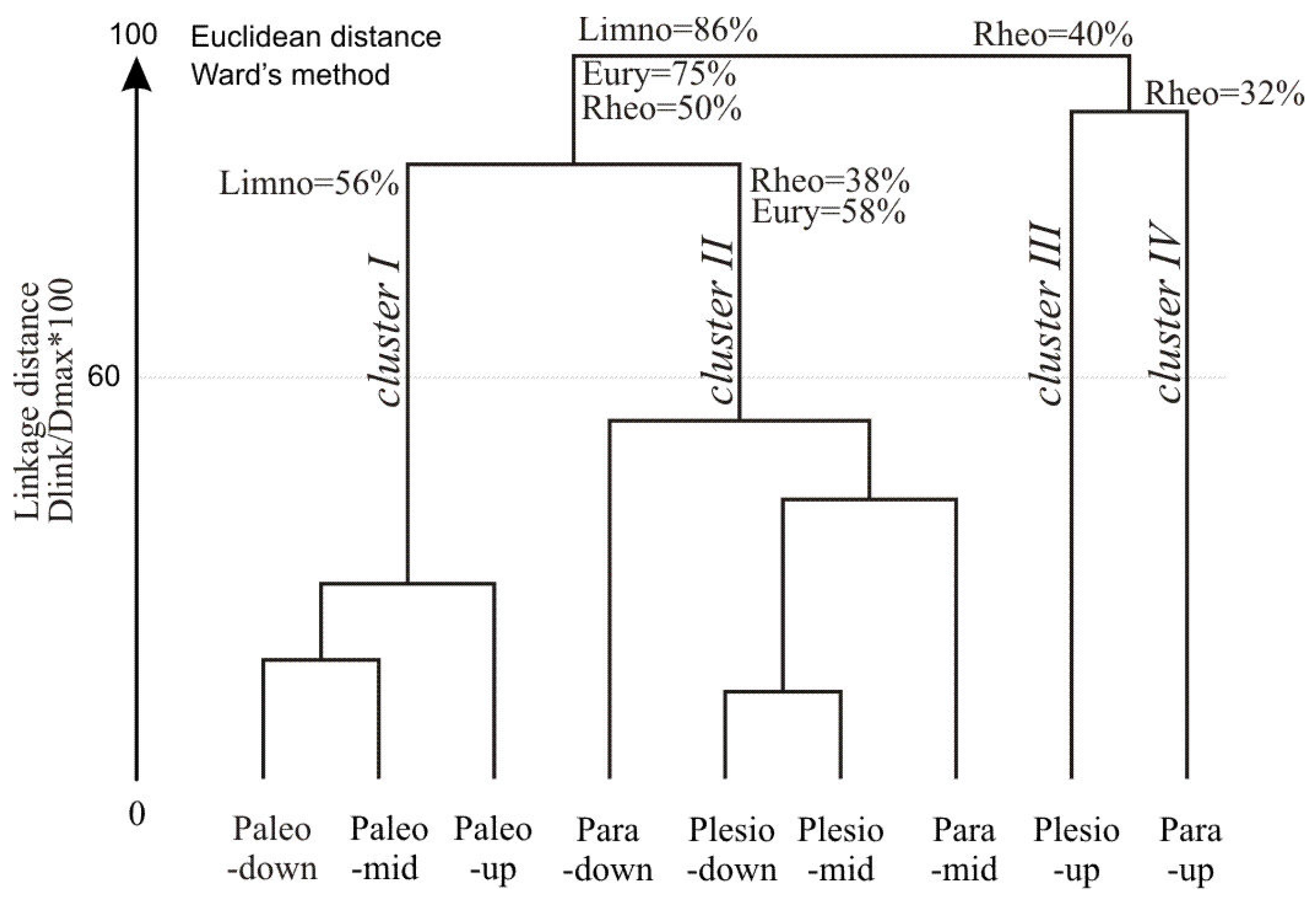
3.3. Fish Response to Habitats along the Connectivity Gradient
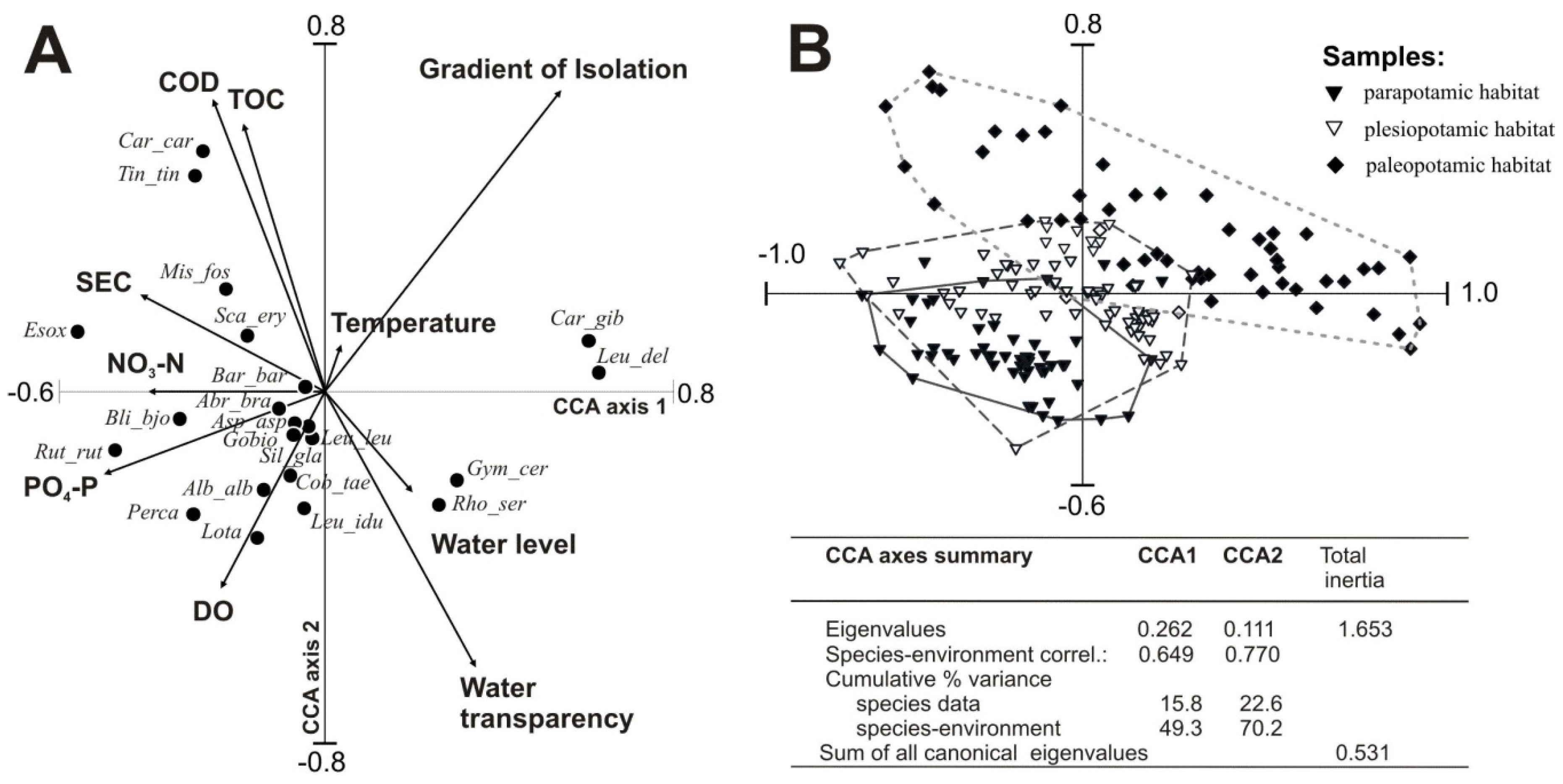
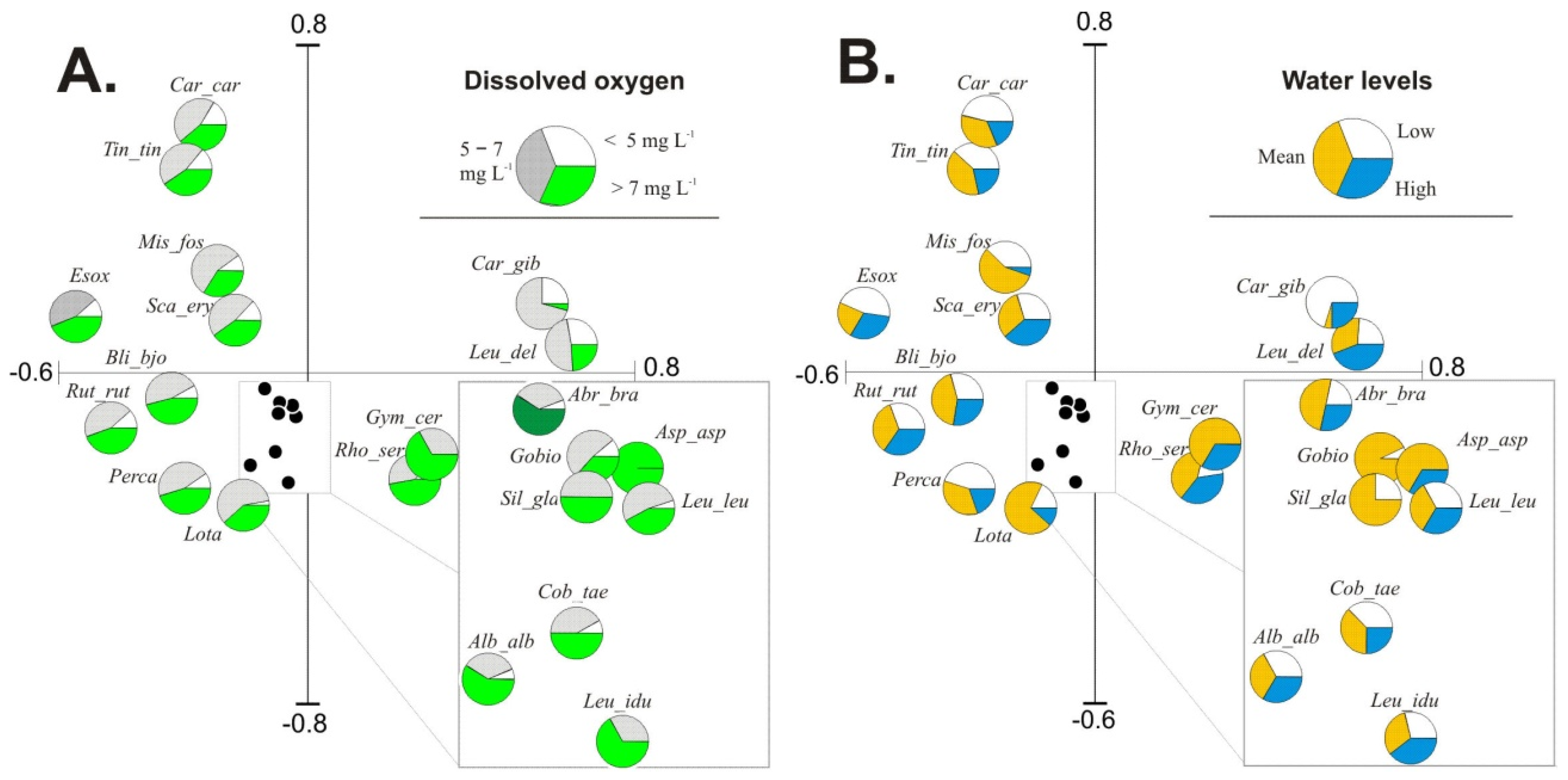
3.4. Influence of Environmental Factors on Fish Abundance and Diversity
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family and Species/Site | Bie-Brza River | Floodplain Lakes | Mean | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parapotamal | Plesiopotamal | Paleopotamal | ||||||||||||
| Up-Stream | Mid-Dle | Down-Stream | Up-Stream | Mid-Dle | Down-Stream | Up-Stream | Mid-Dle | Down-Stream | ||||||
| Esocidae | ||||||||||||||
| Esox lucius | 225● | 156 | 178 | 208 | 181● | 234 | 199 | 233 | 222● | 160 | 163 | 107 | 143□ | 182▼ |
| Cyprinidae | ||||||||||||||
| Abramis brama | 36□ | 46 | 74 | 48 | 56□ | 60 | 103 | 104 | 89□ | 14 | 79 | 29 | 41 | 62□ |
| Alburnus alburnus | 192▼ | 97 | 140 | 26 | 88□ | 111 | 101 | 82 | 98□ | 0 | 36 | 0 | 12 | 66□ |
| Aspius aspius | 3 | 3 | 3 | 0 | 2 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Blicca bjoerkna | 58□ | 120 | 139 | 86 | 115▼ | 302 | 268 | 298 | 289▼ | 115 | 65 | 158 | 113 | 173▼ |
| Carassius auratus gibelio | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 560 | 1076 | 363 | 666● | 222▼ |
| Carassius carassius | 0 | 0 | 14 | 2 | 6 | 23 | 15 | 18 | 19 | 101 | 60 | 47 | 69 | 31 |
| Gobio gobio | 33 | 40 | 3 | 8 | 17 | 40 | 26 | 36 | 34 | 38 | 0 | 0 | 13 | 21 |
| Leucaspius delineatus | 0 | 7 | 0 | 0 | 2 | 61 | 36 | 2 | 33□ | 568 | 649 | 568 | 595● | 210▼ |
| Squalius cephalus | 11 | 6 | 1 | 4 | 4 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 |
| Leuciscus idus | 183▼ | 80 | 32 | 137 | 83□ | 7 | 16 | 25 | 16 | 0 | 0 | 0 | 0 | 33 |
| Leuciscus euciscus | 13 | 33 | 0 | 0 | 11 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 4 |
| Rhodeus sericeus amarus | 126□ | 194 | 66 | 75 | 112□ | 212 | 222 | 187 | 207▼ | 200 | 243 | 243 | 229▼ | 182▼ |
| Rutilus rutilus | 676● | 995 | 936 | 599 | 843● | 1186 | 942 | 1164 | 1097● | 546 | 692 | 889 | 709● | 883● |
| Scardinius erythrophthalmus | 264▼ | 276 | 189 | 140 | 202▼ | 329 | 379 | 301 | 337● | 118 | 274 | 126 | 173▼ | 237● |
| Tinca tinca | 19 | 17 | 24 | 16 | 19 | 49 | 77 | 41 | 56□ | 76 | 93 | 76 | 82 | 52 |
| Cobitidae | ||||||||||||||
| Cobitis taenia | 17 | 6 | 17 | 8 | 10 | 0 | 3 | 7 | 3 | 3 | 0 | 3 | 2 | 5 |
| Misgurnus fossilis | 57□ | 16 | 117 | 86 | 73□ | 18 | 65 | 15 | 33 | 39 | 14 | 64 | 39 | 48 |
| Barbatula barbatula | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Siluridae | ||||||||||||||
| Silurus glanis | 4 | 0 | 1 | 3 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Gadidae | ||||||||||||||
| Lota lota | 94▼ | 43 | 28 | 41 | 37 | 3 | 5 | 8 | 5 | 0 | 1 | 0 | 0 | 14 |
| Percidae | ||||||||||||||
| Gymnocephalus cernua | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Perca fluviatilis | 133▼ | 140 | 121 | 138 | 133▼ | 142 | 160 | 133 | 145▼ | 64 | 58 | 29 | 50 | 110□ |
References
- Amoros, C.; Roux, A.L. Interactions between water bodies within floodplains of large rivers: Function and development of connectivity. Munst. Geogr. Arb. 1988, 29, 125–130. [Google Scholar]
- Penczak, T.; Galicka, W.; Glowacki, Ł.; Koszalinski, H.; Kruk, A.; Zięba, G.; Kostrzewa, J.; Marszal, L. Fish assemblage changes relative to environmental factors and time in the Warta River, Poland, and its oxbow lakes. J. Fish Biol. 2004, 64, 483–501. [Google Scholar]
- Bolland, J.D.; Nunn, A.D.; Lucas, M.C.; Cowx, I.G. The importance of variable lateral connectivity between artificial floodplain waterbodies and river channels. River Res. Appl. 2012, 28, 1189–1199. [Google Scholar] [CrossRef]
- Copp, G.H. The habitat diversity and fish reproductive function of floodplain ecosystems. Environ. Biol. Fish 1989, 26, 1–26. [Google Scholar] [CrossRef]
- Hohausová, E.; Jurajda, P. Restoration of a river backwater and its influence on fish assemblage. Czech J. Anim. Sci. 2005, 50, 473–482. [Google Scholar]
- Fernandes, C.C. Lateral migration of fishes in Amazon floodplains. Ecol. Freshw. Fish 1997, 6, 36–44. [Google Scholar] [CrossRef]
- Agostinho, A.A.; Gomes, L.C.; Zalewski, M. The importance of floodplains for the dynamics of fish communities of the upper River Paraná. Ecohydrol. Hydrobiol. 2001, 1, 209–217. [Google Scholar]
- Welcomme, R.L.; Halls, A. Dependence of tropical river fisheries on flow. In Proceedings of the Second International Symposium on the Management of Large Rivers for Fisheries, Phnom Penh, Kingdom of Cambodia, 11–14 February 2003; Welcomme, R., Petr, T., Eds.; FAO Regional Office for Asia and the Pacific, RAP Publ: Bangkok, Thailand, 2004; pp. 267–283. [Google Scholar]
- Guti, G. Significance of side-tributaries and floodplains for the Danubian fish populations. Large Rivers. Arch. Hydrobiol. Suppl. 2002, 13, 151–163. [Google Scholar]
- Schomaker, C.; Wolter, C. The contribution of long-term isolated water bodies to floodplain fish diversity. Freshw. Biol. 2011, 56, 1469–1480. [Google Scholar] [CrossRef]
- Tockner, K.; Pennetzdorfer, D.; Reiner, N.; Schiemer, F.; Ward, J.V. Hydrological connectivity and the exchange of organic matter and nutrients in a dynamic river-floodplain system (Danube, Austria). Freshw. Biol. 1999, 41, 521–535. [Google Scholar] [CrossRef]
- Miranda, L.E. Fish assemblages in oxbow lakes relative to connectivity with the Mississippi River. Trans. Am. Fish Soc. 2005, 134, 1480–1489. [Google Scholar] [CrossRef]
- Dembkowski, D.J.; Miranda, L.E. Hierarchy in factors affecting fish biodiversity in floodplain lakes of the Mississippi Alluvial Valley. Environ. Biol. Fish 2012, 93, 357–368. [Google Scholar] [CrossRef]
- Witkowski, A. An analysis of the ichthyofauna of the Biebrza River system. Part II. Materials to the knowledge of the ichthyofauna and review of species. Fragm Faun 1984, 28, 137–184. (In Polish) [Google Scholar] [CrossRef]
- Winemiller, K.O.; Terim, S.; Shormann, D.; Cotner, J.B. Fish assemblage structure in relation to environmental variation among Brazos River oxbow lakes. Trans. Am. Fish Soc. 2000, 129, 451–468. [Google Scholar] [CrossRef]
- Schiemer, F.; Spindler, T. Endangered fish species of the Danube River in Austria. Regul. River Res. Manag. 1989, 4, 397–407. [Google Scholar] [CrossRef]
- Lubinski, B.J.; Jackson, J.R.; Eggleton, M.A. Relationships between floodplain lake fish communities and environmental variables in a large river-floodplain ecosystem. Trans. Am. Fish Soc. 2008, 137, 895–908. [Google Scholar] [CrossRef]
- Rodriguez, M.A.; Lewis, W.M., Jr. Structure of fish assemblages along environmental gradients in floodplain lakes of the Orinoco River. Ecol. Monogr. 1997, 67, 109–128. [Google Scholar] [CrossRef]
- Schiemer, F.; Waidbacher, H. Strategies for Conservation of Danubian Fish Fauna. In River Conservation and Management; Boon, P.J., Calow, P., Petts, G.J., Eds.; Wiley-Blackwell: London, UK, 1992; pp. 363–382. [Google Scholar]
- Welcomme, R.L.; Winemiller, K.O.; Cowx, I.G. Fish environmental guilds as a tool for assessment of ecological condition of rivers. River Res. Appl. 2006, 22, 377–396. [Google Scholar] [CrossRef]
- Tockner, K.; Baumgartner, C.; Schiemer, F.; Ward, J.V. Biodiversity of a Danubian Floodplain: Structural, Functional and Compositional Aspects. In Biodiversity in Wetlands: Assessment, Function and Conservation; Gopal, B., Junk, W.J., Davis, J.A., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2000; pp. 141–159. [Google Scholar]
- Ward, J.V.; Tockner, K.; Schiemer, F. Biodiversity of floodplain river ecosystems: Ecotones and connectivity. Regul. Rivers Res. Manag. 1999, 15, 125–139. [Google Scholar] [CrossRef]
- Junk, W.J.; Wantzen, K.M. The flood pulse concept: New aspects, approaches and applications—An update. In Proceedings of the Second International Symposium on the Management of Large Rivers for Fisheries, Phnom Penh, Kingdom of Cambodia, 11–14 February 2003; Welcomme, R., Petr, T., Eds.; FAO Regional Office for Asia and the Pacific, RAP Publication: Bangkok, Thailand, 2004; pp. 117–140. [Google Scholar]
- Grabowska, M.; Glińska-Lewczuk, K.; Obolewski, K.; Burandt, P.; Kobus, S.; Dunalska, J.; Kujawa, R.; Goździejewska, A.; Skrzypczak, A. Effects of Hydrological and Physicochemical Factors on Phytoplankton Communities in Floodplain Lakes. Pol. J. Environ. Stud. 2014, 23, 713–725. [Google Scholar]
- Hein, T.; Baranyi, C.; Reckendorfer, W.; Schiemer, F. The impact of surface water exchange on the nutrient and particle dynamics in side-arms along the River Danube, Austria. Sci. Total Environ. 2004, 328, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Molls, F.; Neumann, D. Fish abundance and fish migration in gravel-pit lakes connected with the River Rhine. Water Sci. Technol. 1994, 29, 307–309. [Google Scholar]
- Allouche, S.; Thevenet, A.; Gaudin, P. Habitat use by chub (Leuciscus cephalus) in a large river, the French Upper Rhone, as determined by radiotelemetry. Arch. Hydrobiol. 1999, 145, 219–236. [Google Scholar]
- Schwartz, J.S.; Herricks, E.E. Fish use of stage-specific fluvial habitats as refuge patches during a flood in a low-gradient Illinois stream. Can. J. Fish. Aquat. Sci. 2005, 62, 1540–1552. [Google Scholar] [CrossRef]
- Schmutz, S.; Kaufmann, M.; Vogel, B.; Jungwirth, M.; Muhar, S. A Multi-Level Concept for Fish-Based, River-Type-Specific Assessment of Ecological Integrity; Springer: Berlin, Germany, 2000; pp. 279–289. [Google Scholar]
- WFD, EU. Directive 2000/60/EC of the European Parliament and of the Council Establishing a Framework for the Community Action in the Field of Water Policy; The European Parliament and the Council of the European Union: Brussels, Belgium, 2000. [Google Scholar]
- House, A. Council Directive 92/43/EEC on the Conservation of Natural Habitats and of Wild Fauna and Flora; European Commission: Brussels, Belgium, 2014. [Google Scholar]
- Witkowski, A. Structure of communities and biomass of ichthyofauna in the Biebrza River, its old river beds and affluents. Pol. Ecol. Stud. 1984, 10, 447–474. [Google Scholar]
- Wassen, M.J.; Peeters, W.H.M.; Veneterink, H.O. Patterns in vegetation, hydrology and nutrient availability in an undisturbed river floodplain in Poland. Plant Ecol. 2002, 165, 27–43. [Google Scholar] [CrossRef]
- Chormański, J.; Okruszko, T.; Ignar, S.; Batelaan, O.; Rebel, K.T.; Wassen, M.J. Flood mapping with remote sensing and hydrochemistry: A new method to distinguish the origin of flood water during floods. Ecol. Eng. 2011, 37, 1334–1349. [Google Scholar] [CrossRef]
- Standard Methods for the Examination of Water and Wastewater, 20th ed.; American Public Health Association: Washington, DC, USA, 1998.
- Dunalska, J.A.; Górniak, D.; Jaworska, B.; Evelyn, E.; Gaiser, E.E. Effect of temperature on organic matter transformation in a different ambient nutrient availability. Ecol. Eng. 2012, 49, 27–34. [Google Scholar] [CrossRef]
- Persat, H.; Copp, G.H. Electrofishing and Point Abundance Sampling for the Ichthyology of Large Rivers. In Developments in Electrofishing. Fishing New Books; Cowx, I.G., Ed.; Blackwell Scientific Publications Ltd: Oxford, UK, 1989; pp. 205–215. [Google Scholar]
- European Standard CSN-EN 14011-Water Quality-Sampling of Fish with Electricity; European Committee for Standardization: Brussels, Belgium, 2006.
- Obolewski, K. Epiphytic macrofauna on water soldiers (Stratiotes aloides L.) in Slupia River oxbows. Oceanol. Hydrobiol. Stud. 2005, 34, 37–54. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Ter Braak, C.J.F.; Šmilauer, P. Reference Manual and CanoDraw for Windows User's Guide; Version 4.5; Software for Canonical Community Ordination—Microcomputer Power; Biometrics: Ithaca, NY, USA, 2002. [Google Scholar]
- Damme, D.V.; Bogutskaya, N.; Hoffmann, R.C.; Smith, C. The introduction of the European bitterling (Rhodeus amarus) to west and central Europe. Fish Fish. 2007, 8, 79–106. [Google Scholar] [CrossRef]
- Kwak, T.J. Lateral movement and use of floodplain habitat by fishes of the Kankakee River Illinois. Am. Midl. Nat. 1988, 120, 241–249. [Google Scholar] [CrossRef]
- Amoros, C.; Bornette, G. Connectivity and biocomplexity in waterbodies of riverine floodplains. Freshw. Biol. 2002, 47, 761–776. [Google Scholar] [CrossRef]
- Miranda, L.E.; Andrews, C.S.; Kröger, R. Connectedness of land use, nutrients, primary production, and fish assemblages in oxbow lakes. Aquat. Sci. 2014, 76, 41–50. [Google Scholar] [CrossRef]
- Holopainen, I.J.; Tonn, W.M.; Paszkowski, C.A. Tales of two fish: The dichotomous biology of crucian carp (Carassius carassius (L.)) in northern Europe. Ann. Zool. Fenn. 1997, 34, 1–22. [Google Scholar]
- Galat, D.L.; Fredrickson, L.H.; Humburg, D.D.; Bataille, K.J.; Bodie, J.R.; Dohrenwend, J.; Gelwicks, G.T.; Havel, J.E.; Helmers, D.L.; Hooker, J.B.; et al. Flooding to restore connectivity of regulated, large-river wetlands. BioScience 1998, 48, 721–733. [Google Scholar] [CrossRef]
- Petry, A.C.; Agostinho, A.A.; Gomes, L.C. Fish assemblages of tropical floodplain lagoons: Exploring the role of connectivity in a dry year. Neotropical Ichthyol. 2003, 1, 111–119. [Google Scholar] [CrossRef]
- White, J.L.; Harvey, B.C. Effects of an introduced piscivorous fish on native benthic fishes in a Coastal River. Freshw. Biol. 2001, 46, 987–995. [Google Scholar] [CrossRef]
- Wiśniewolski, W.; Ligęza, J.; Prus, P.; Buras, P.; Szlakowski, J.; Borzęcka, I. Znaczenie łączności rzeki ze starorzeczami dla składu ichtiofauny na przykładzie środkowej i dolnej Wisły. Nauka Przyr. Technol. 2009, 3, 2–10. (In Polish) [Google Scholar]
- Wiśniewolski, W.; Szlakowski, J.; Buras, P.; Klein, M. Ichtiofauna (Ichthyofauna). In Kotlina Biebrzańska i Biebrzański Park Narodowy; Banaszuk, H., Ed.; WE&S: Białystok, Poland, 2004; pp. 455–489. (In Polish) [Google Scholar]
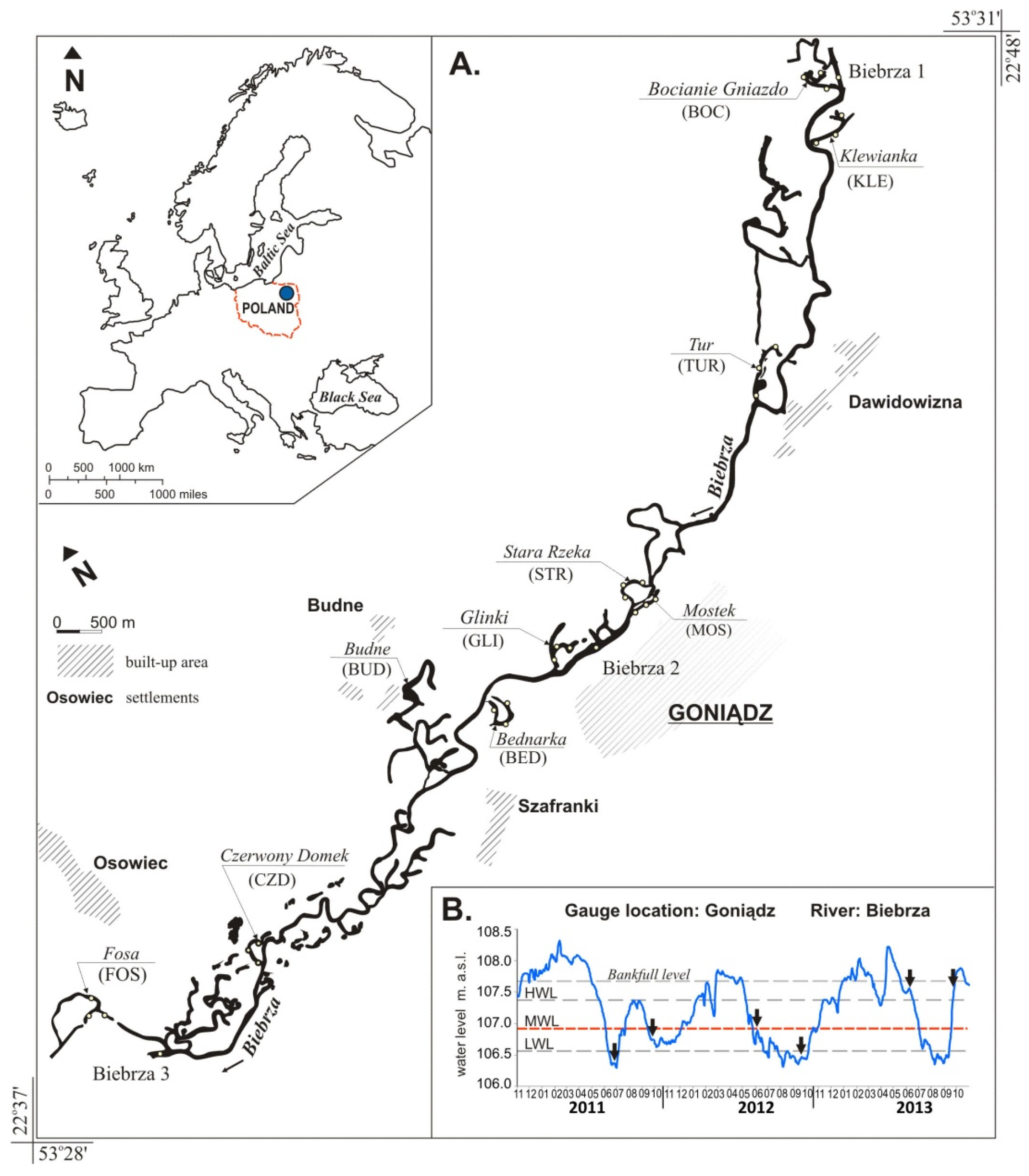
| Floodplain Lake (local Name) | Geographical Coordinates | Type of Connection | L (m) | W (m) | A (ha) | Distance from the River Channel | Depth | Sub-Strate | Macrophyte Cover | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Upstream Arm (m) | Downstream Arm (m) | Max. Distance (m) | hav (m) | hmax (m) | Emerged (%) | Submerged (%) | |||||||
| Stara Rzeka | N:53°30’0.26” E:22°44’37.2” | Parapotamic (lotic) | 1380 | 24 | 3.31 | 0 | 0 | 523 | 2.2 | 3.4 | mineral (gravel, pebbels) | 12 | 7 |
| Czerwony domek | N:53°29’21.53” E:22°39’54.54” | Parapotamic (lotic) | 548 | 17 | 1.38 | 0 | 0 | 136 | 1.8 | 3.3 | mineral (gravel) | 14 | 11 |
| Mostek | N:53°29’35.9” E:22°44’15.95” | Parapotamic (lotic) | 306 | 20 | 0.62 | 0 | 0 | 30 | 1.0 | 1.4 | mineral (sand, gravel) | 10 | 5 |
| Bocianie Gniazdo | N:53°31’15.47” E:22°47’55.62” | Plesio-potamic (semi-lotic) | 569 | 25 | 1.43 | 104 | 0 | 304 | 2.8 | 5.5 | mineral (sand, gravel), organic | 18 | 29 |
| Klewianka | N:53°30’57.34” E:22°47’55.87” | Plesio-potamic (semi-lotic) | 520 | 22 | 1.41 | 80 | 0 | 260 | 2.6 | 4.1 | mineral (sand, gravel), organic | 21 | 36 |
| Tur | N:53°30’15.68” E:22°46’17.54” | Plesio-potamic (semi-lotic) | 678 | 21 | 2.03 | 51 | 0 | 150 | 2.2 | 3.9 | organic, mineral (sand) | 18 | 31 |
| Glinki | N:53°29’40.29” E:22°43’28.94” | Plesio-potamic (semi-lotic) | 459 | 32 | 1.48 | 150 | 0 | 226 | 2.2 | 5.5 | organic, mineral (sand) | 24 | 59 |
| Bednarka | N:53°29’31.51” E:22°42’43.46” | Paleo-potamic (lentic) | 740 | 29 | 2.16 | 50 | 42 | 313 | 1.6 | 2.6 | organic | 38 | 59 |
| Budne | N:53°29’56.68” E:22°42”14.06” | Paleo-potamic (lentic) | 1652 | 26 | 4.41 | 407 | 380 | 680 | 1.2 | 1.8 | organic | 21 | 76 |
| Fosa | N:53°29’32.49” E:22°38’36.27” | Paleo-potamic (lentic) | 1360 | 31 | 4.30 | 547 | 792 | 817 | 2.9 | 5.6 | organic, mineral (sand) | 10 | 28 |
| Type of Hydrological Connectivity | Parapotamic Lotic | Plesiopotamic Semi-Lotic | Paleopotamic Lentic | All floodplain Lakes | Biebrza River | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameter | Unit | ±SD | ±SD | ±SD | ±SD | ±SD | |||||
| Temperature | (°C) | 17.13 | ±3.38 | 16.61 | ±3.06 | 17.40 | ±3.55 | 17.01 | ±3.31 | 17.4 | 3.42 |
| pH * | pH units | 7.83 b | 7.67 # 7.94 | 7.77 a,b | 7.56 # 7.95 | 7.69 a | 7.47 # 7.90 | 7.77 | 7.58 # 7.92 | 7.87 | 7.79 # 7.92 |
| DO | (mg·L−1) | 7.45 b | ±1.54 | 6.51 a | ±2.12 | 6.13 a | ±2.31 | 6.71 | ±2.08 | 7.83 | ±1.47 |
| SEC | (uS·sm−1) | 536 | ±90 | 553 | ±115 | 535 | ±122 | 542 | ±110 | 525 | ±84 |
| Chl_a | (μg·L−1) | 8.04 a | ±1.03 | 17.61 | ±12.85 | 17.47 | ±8.31 | 14.70 | ±10.25 | 7.31 | ±0.80 |
| CODCr | (mg·L−1) | 37.34 a | ±9.63 | 41.84 a,b | ±9.65 | 48.92 b | ±21.48 | 42.63 | ±14.96 | 36.51 | ±10.01 |
| TN | (mg·L−1) | 1.07 | ±0.38 | 1.27 | ±0.38 | 1.39 | ±0.57 | 1.24 | ±0.46 | 0.98 | ±0.32 |
| NO2-N | (mg·L−1) | 0.013 b | ±0.013 | 0.006 a | ±0.006 | 0.005 a | ±0.004 | 0.008 | ±0.009 | 0.010 | ±0.71 |
| NO3-N | (mg L−1) | 0.17 c | ±0.08 | 0.12 b | ±0.06 | 0.10 a | ±0.03 | 0.13 | ±0.07 | 0.17 | ±0.08 |
| NH4-N | (mg·L−1) | 0.14 a | ±0.07 | 0.19 b | ±0.08 | 0.27 c | ±0.22 | 0.20 | ±0.14 | 0.16 | ±0.09 |
| TP | (mg·L−1) | 0.33 | ±0.17 | 0.34 | ±0.16 | 0.38 | ±0.21 | 0.35 | ±0.18 | 0.33 | ±0.14 |
| PO4-P | (mg·L−1) | 0.12 b | ±0.05 | 0.08 a | ±0.05 | 0.07 a | ±0.04 | 0.09 | ±0.05 | 0.12 | ±0.05 |
| TOC | (mg·L−1) | 10.70 a | ±2.94 | 12.71 a,b | ±4.58 | 14.34 a | ±5.42 | 12.59 | ±4.64 | 10.30 | ±1.81 |
| DOC | (mg·L−1) | 9.26 | ±2.29 | 10.75 | ±3.38 | 11.43 | ±3.37 | 10.50 | ±3.18 | 9.10 | ±1.31 |
| Water transparency | (m) | 2.0 b | ±0.9 | 1.5 b | ±0.6 | 1.1 a | ±0.3 | 1.6 | ±0.8 | 2.5 | ±0.7 |
| Family Species | Abbreviation | Common Name | Feeding Guild | Reproductive Guild | Stream Velocity Preference | Constancy of Occurrence (C, %) | Conservation Status |
|---|---|---|---|---|---|---|---|
| Esocidae | |||||||
| Esox lucius | Esox | pike | P | phytophils | eurytopic | 88 | n.t. |
| Cyprinidae | |||||||
| Abramis brama | Abr_br | bream | O | phytophils | eurytopic | 36 | n.t. |
| Alburnus alburnus | Alb_al | bleak | I | phytophils | eurytopic | 26 | n.t. |
| Aspius aspius | Asp_as | asp | P | lithophils | rheophilic | 2 | HD-AII |
| Blicca bjoerkna | Bli_bj | silver bream | I | phyto-lithophils | eurytopic | 62 | n.t. |
| Carassius auratus gibelio | Car_gi | Prussian carp | O, E | phytophils | limnophilic | 14 | n.t. |
| Carassius carassius | Car_ca | crucian carp | O | phytophils | limnophilic | 31 | n.t. |
| Gobio gobio | Gobio | gudgeon | I | psammophils | rheophilic | 10 | n.t. |
| Leucaspius delineatus | Leu_del | sunbleak | I | phytophils | limnophilic | 14 | n.t. |
| Squalius cephalus | Squ_cep | chub | O/P | lithophils | rheophilic | 3 | n.t. |
| Leuciscus idus | Leu_idu | ide | 0 | phyto-lithophils | rheophilic | 21 | n.t. |
| Leuciscus leuciscus | Leu_le | dace | I | phyto-lithophils | rheophilic | 2 | n.t. |
| Rhodeus sericeus amarus | Rho_se | bitterling | H | ostracophils | limnophilic | 55 | HD-AII, IUCN |
| Rutilus rutilus | Rut_ru | roach | O | phyto-lithophils | eurytopic | 88 | n.t. |
| Scardinius erythrophthalmus | Sc_ery | rudd | O | phytophils | limnophilic | 73 | n.t. |
| Tinca tinca | Ti_tin | tench | I | phytophils | limnophilic | 60 | n.t. |
| Cobitidae | |||||||
| Cobitis taenia | Cob_tae | spined loach | I | phytophils | rheophilic | 14 | HD-AII, IUCN |
| Misgurnus fossilis | Misg_fo | weatherfish | I | phytophils | limnophilic | 33 | HD-AII, IUCN |
| Barbatula barbatula | Bar_ba | stone loach | I | psammophils | rheophilic | 1 | IUCN |
| Siluridae | |||||||
| Silurus glanis | Sil_gl | wels catfish | P | phytophils | eurytopic | 2 | n.t. |
| Gadidae | |||||||
| Lota lota | Lota_lo | burbot | I/P | litho-pelagophils | rheophilic | 19 | n.t. |
| Percidae | |||||||
| Gymnocephalus cernua | Gym_ce | ruffe | I | phyto-lithophils | eurytopic | 2 | n.t. |
| Perca fluviatilis | Perca | perch | I/P | phyto-lithophils | eurytopic | 69 | n.t. |
| Site Fish guilds or Metrics | Biebrza River | Floodplain Lakes | All Floodplain Lakes | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parapotamic | Plesiopotamic | Paleopotamic | ||||||||||||
| Upstream | Middle | Downstream | Upstream | Middle | Downstream | Upstream | Middle | Downstream | ||||||
| Rheophils, (%) | 16.3 | 9.3 | 4.0 | 12.2 | 8.2 | 1.9 | 2.0 | 2.9 | 2.3 | 1.5 | 0 | 0.1 | 0.5 | 3.1 |
| Eurytops, (%) | 65.7 | 68.3 | 76.3 | 68.2 | 71.1 | 73.2 | 67.7 | 75.8 | 72.3 | 34.6 | 31.2 | 44.9 | 36.4 | 59.4 |
| Limnophils, (%) | 18.0 | 22.4 | 19.7 | 19.6 | 20.7 | 24.9 | 30.3 | 21.3 | 25.5 | 63.9 | 68.8 | 55.0 | 63.1 | 37.5 |
| Total (specimens ha−1) | 2164 | 2275 | 2083 | 1628 | 1995 | 2782 | 2623 | 2655 | 2687 | 2603 | 3503 | 2703 | 2936 | 2552 |
| Spec. contribution (%)* | – | 9.1 | 8.3 | 6.5 | 23.9 | 14.2 | 13.4 | 13.5 | 41.0 | 10.4 | 14.0 | 10.8 | 35.1 | 100 |
| Biomass (kg·ha−1) | 94.3 | 70.9 | 79.2 | 85.1 | 78.4 | 82.0 | 74.8 | 98.3 | 85.0 | 62.3 | 92.7 | 53.1 | 69.4 | 77.6 |
| Biomass contribution (%) * | – | 10.2 | 11.3 | 12.2 | 33.7 | 11.7 | 10.7 | 14.1 | 36.5 | 8.9 | 13.3 | 7.6 | 29.8 | 100 |
| Species richness, S ** | 18 | 18 | 18 | 18 | 21 | 18 | 19 | 19 | 23 | 13 | 14 | 16 | 17 | 23 |
| Shannon’s biodiversity index, H’ ** | 2.11 | 1.49 | 1.44 | 1.41 | 2.45 | 1.47 | 1.59 | 1.51 | 2.51 | 1.20 | 1.29 | 1.18 | 2.23 | 2.41 |
| Pielou’s evenness index, J’ ** | 0.91 | 0.74 | 0.71 | 0.73 | 0.79 | 0.71 | 0.78 | 0.72 | 0.80 | 0.73 | 0.77 | 0.73 | 0.82 | 0.80 |
| Variable | Marginal Effects | Conditional Effects | ||
|---|---|---|---|---|
| λ1 * | λA ** | P | F-Value | |
| Isolation from the river | 0.15 | 0.13 | 0.002 | 14.92 |
| Water transparency | 0.06 | 0.07 | 0.002 | 8.86 |
| SEC | 0.05 | 0.05 | 0.002 | 6.06 |
| DO | 0.03 | 0.05 | 0.002 | 5.88 |
| CODCr | 0.05 | 0.03 | 0.006 | 3.91 |
| Temperature | 0.02 | 0.03 | 0.004 | 3.77 |
| Water level | 0.03 | 0.02 | 0.004 | 3.53 |
| PO4–P | 0.07 | 0.03 | 0.004 | 3.12 |
| NO3–N | 0.04 | 0.01 | 0.020 | 2.33 |
| TOC | 0.04 | 0.01 | 0.020 | 2.32 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glińska-Lewczuk, K.; Burandt, P.; Kujawa, R.; Kobus, S.; Obolewski, K.; Dunalska, J.; Grabowska, M.; Lew, S.; Chormański, J. Environmental Factors Structuring Fish Communities in Floodplain Lakes of the Undisturbed System of the Biebrza River. Water 2016, 8, 146. https://doi.org/10.3390/w8040146
Glińska-Lewczuk K, Burandt P, Kujawa R, Kobus S, Obolewski K, Dunalska J, Grabowska M, Lew S, Chormański J. Environmental Factors Structuring Fish Communities in Floodplain Lakes of the Undisturbed System of the Biebrza River. Water. 2016; 8(4):146. https://doi.org/10.3390/w8040146
Chicago/Turabian StyleGlińska-Lewczuk, Katarzyna, Paweł Burandt, Roman Kujawa, Szymon Kobus, Krystian Obolewski, Julita Dunalska, Magdalena Grabowska, Sylwia Lew, and Jarosław Chormański. 2016. "Environmental Factors Structuring Fish Communities in Floodplain Lakes of the Undisturbed System of the Biebrza River" Water 8, no. 4: 146. https://doi.org/10.3390/w8040146
APA StyleGlińska-Lewczuk, K., Burandt, P., Kujawa, R., Kobus, S., Obolewski, K., Dunalska, J., Grabowska, M., Lew, S., & Chormański, J. (2016). Environmental Factors Structuring Fish Communities in Floodplain Lakes of the Undisturbed System of the Biebrza River. Water, 8(4), 146. https://doi.org/10.3390/w8040146