

Archaeozoology Supports a Holistic View on Fish Assessments in Large Rivers—A Case Study from the Volga River: From Quantitative Data and Ancient DNA to Biodiversity Analysis

 , , , , , ,
, , , , , ,  ,
,  and
and 
Abstract
1. Introduction
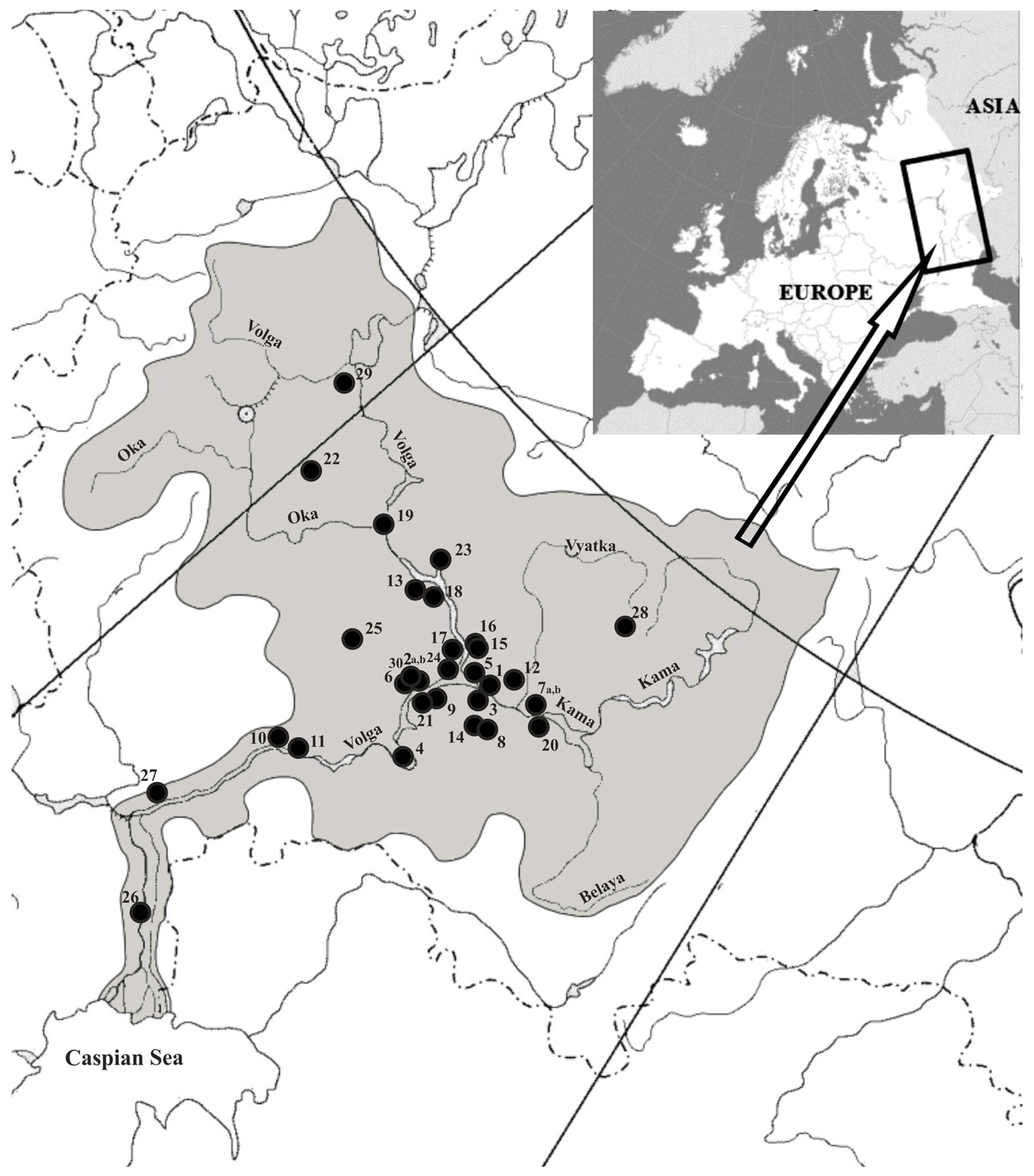
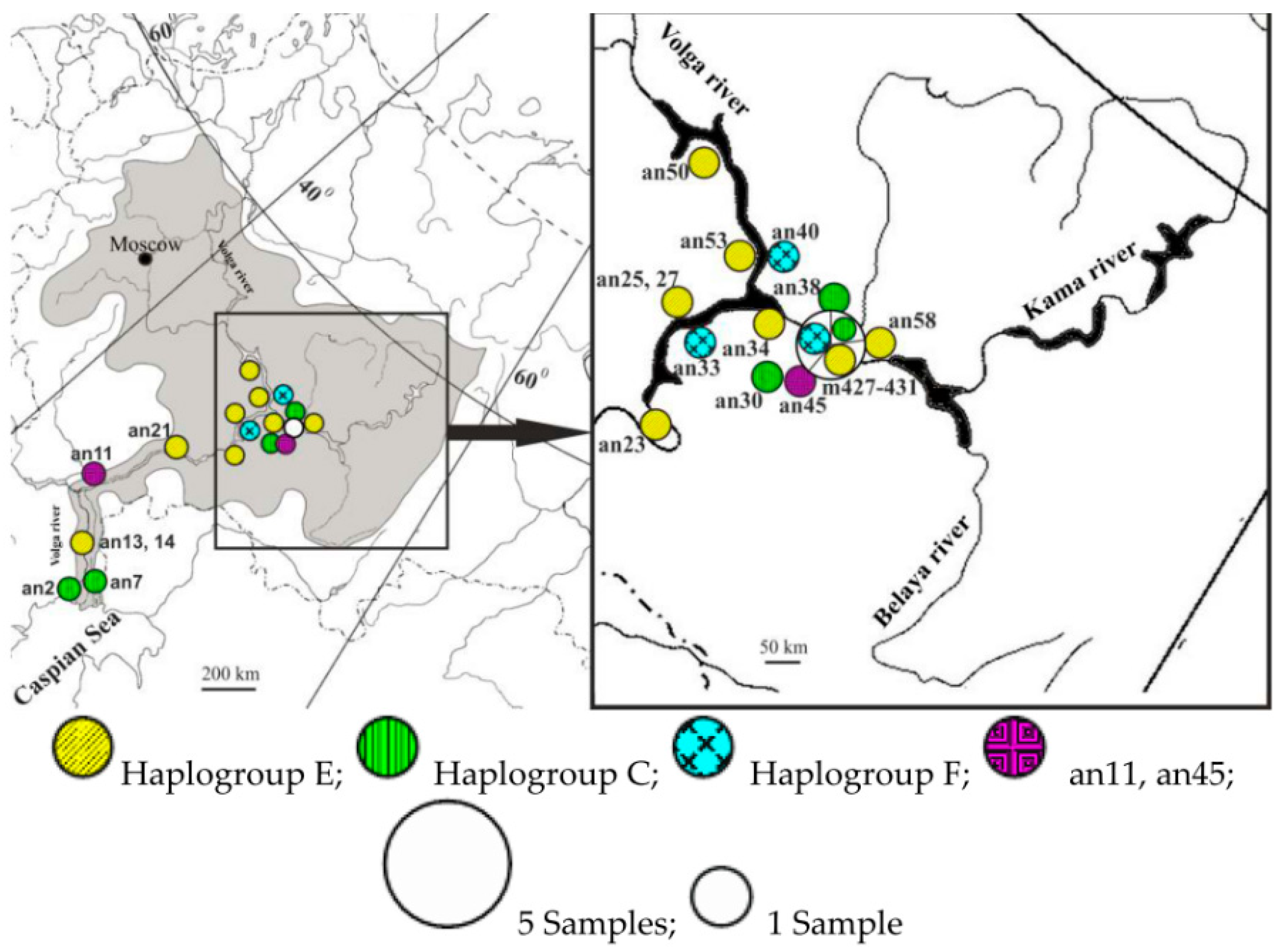
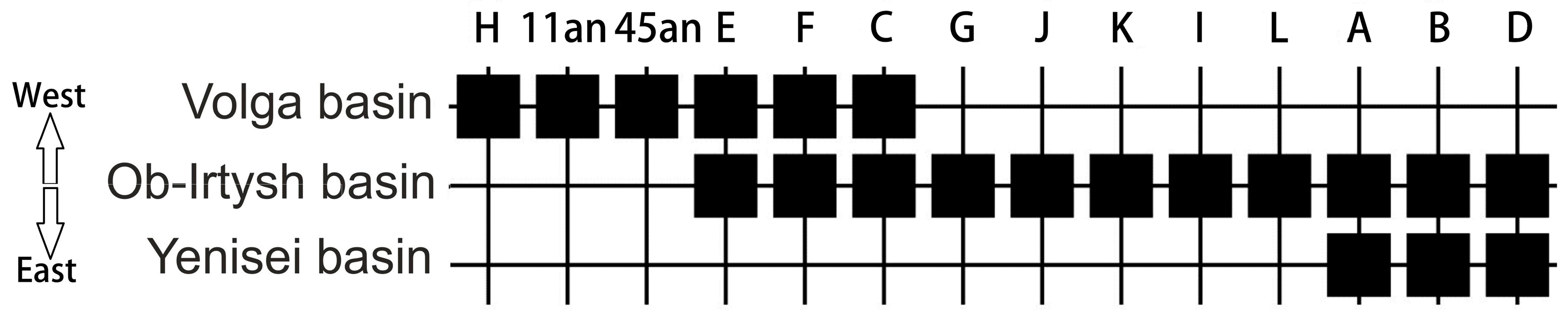
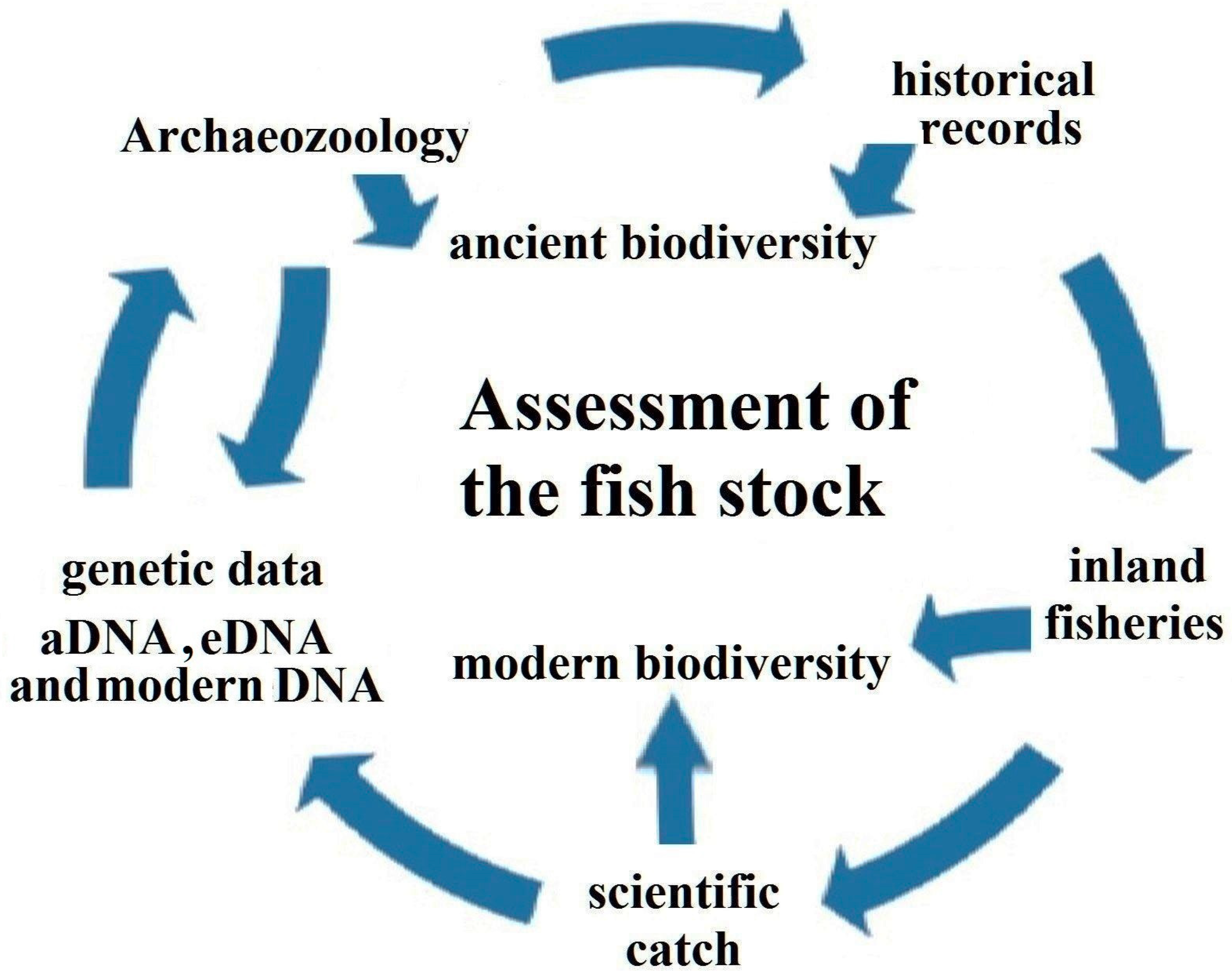
2. Materials and Methods
or
ΣMNI large fish >50 cm/ΣMNI large fish >50 cm + ΣMNI small fish < 50 cm
- To estimate the value of each of these species in ancient catches according to historical periods, we introduced a species-index (“index Salmo caspius”, “index Acipenser stellatus”, “index Stenodus leucichthys”, “index Acipenser ruthenus”) expressing the ratio of the total number of individuals (MNI) of the special species to the total number of individuals (MNI) of other fish species. Such indices are successfully used in the analysis of archaeozoological materials, in particular, fish remains [14,20].
- To assess trends in fish size over time, the LSI, “Logarithmic size Index”, used this index, which is often used in archaeozoology, and was assessed in comparison with the sizes of fish from modern populations: LSI = log (Mx/Ms) = log (Mx) − log (Ms) (Mx: average restored size (length) of fish (TL—sturgeons and Lsm—salmonids) in samples for each historical period (Ms: average size of fish (length) in samples for the first half of the 20th century from the Volga River as a standard sample). The average sizes of fish (length) (TL—Acipenser stellatus, Acipenser ruthenus, and Lsm—salmonids) in samples from the first half of the 20th century from the Volga River are: starry sturgeon—133.3 cm [40,41]; Acipenser ruthenus—42.9 cm [42]; Salmo caspius—88.4 cm [43]; and Stenodus leucichthys—89.0 cm [41].
- One of the key environmental factors affecting fish populations is climate change. The main climate component is temperature. In the past two decades, much data have been obtained on temperature changes over the last two millennia in the Northern Hemisphere, including the Russian Plain within the Volga basin [44]. Based on these data, we reconstructed the indicators of average annual air temperature for each of the four time periods within the entire Volga basin (Table 1). For this purpose, the perennial fields of average annual air temperature (average data for 1951–1980) of meteorological stations in 22 nodal geographical squares of 250 km × 250 km on the territory of the Volga basin were analyzed (data obtained from the database of the All-Russian Research Institute of Hydrometeorology and Information Center http://meteo.ru/, accessed on 2 February 2024). For the values of the deviation of the average annual temperature from the average data of 1951 to 1980 for each century, data from the work of V. Klimenko and O. Solomina [44] were used.
- Another environmental factor related to climate change and significantly affecting the number of anadromous fish is the hydrological regime. Over the past two thousand years, the level regime of the Caspian Sea has changed significantly. The water balance of the Caspian Sea is determined mainly by river runoff and precipitation (input part) and evaporation (expenditure part). In the input part, the river Volga plays a decisive role, the share of which is approximately 80% of the total water input into the sea. It is believed that the fluctuation in the sea level is determined by climate fluctuations in the entire vast Caspian basin. The average change in the level of the Caspian Sea for each of the four time periods (Table 1) is calculated according to the data from the monograph “The Caspian Sea: Extreme Hydrological Events” [45]. Over the past 2000 years, the range of changes in the level of the Caspian Sea (by decade) was 11.2 m: from −34.5 to −23.3 m. The minimum levels over the centuries in the last 2000 years were during the Derbent regression in the 6th century AD (on average for the century—32.7 m) and in the 12th century during the period of the “Medieval Temperature Maximum” (on average for the century—30.7 m). The greatest levels occurred in the 17th and 18th centuries, during the “Little Ice Age” (on average for each century:24.8 m and 24.5 m, respectively).
3. Results
Biodiversity Measures from Archaeoichthyological Collections
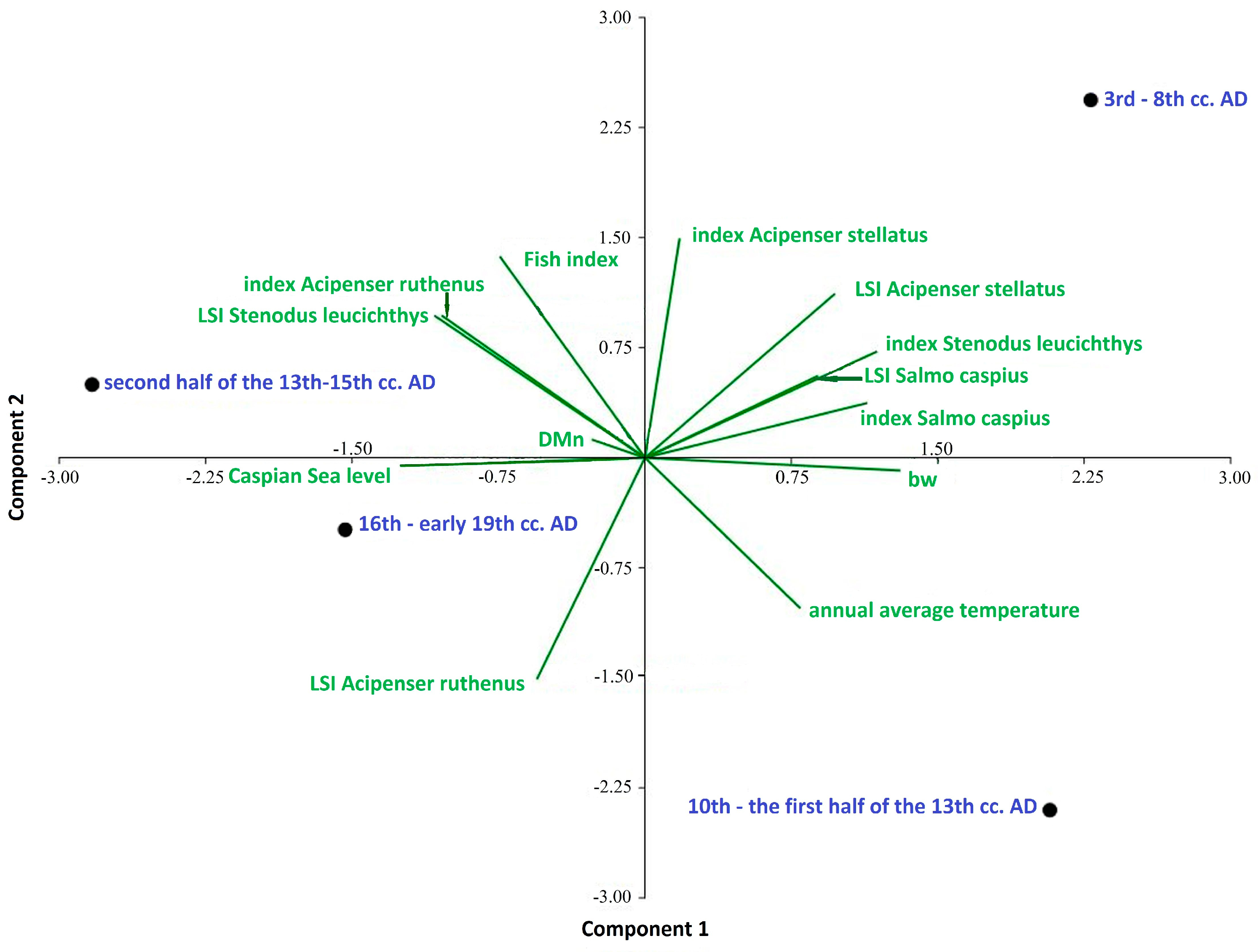
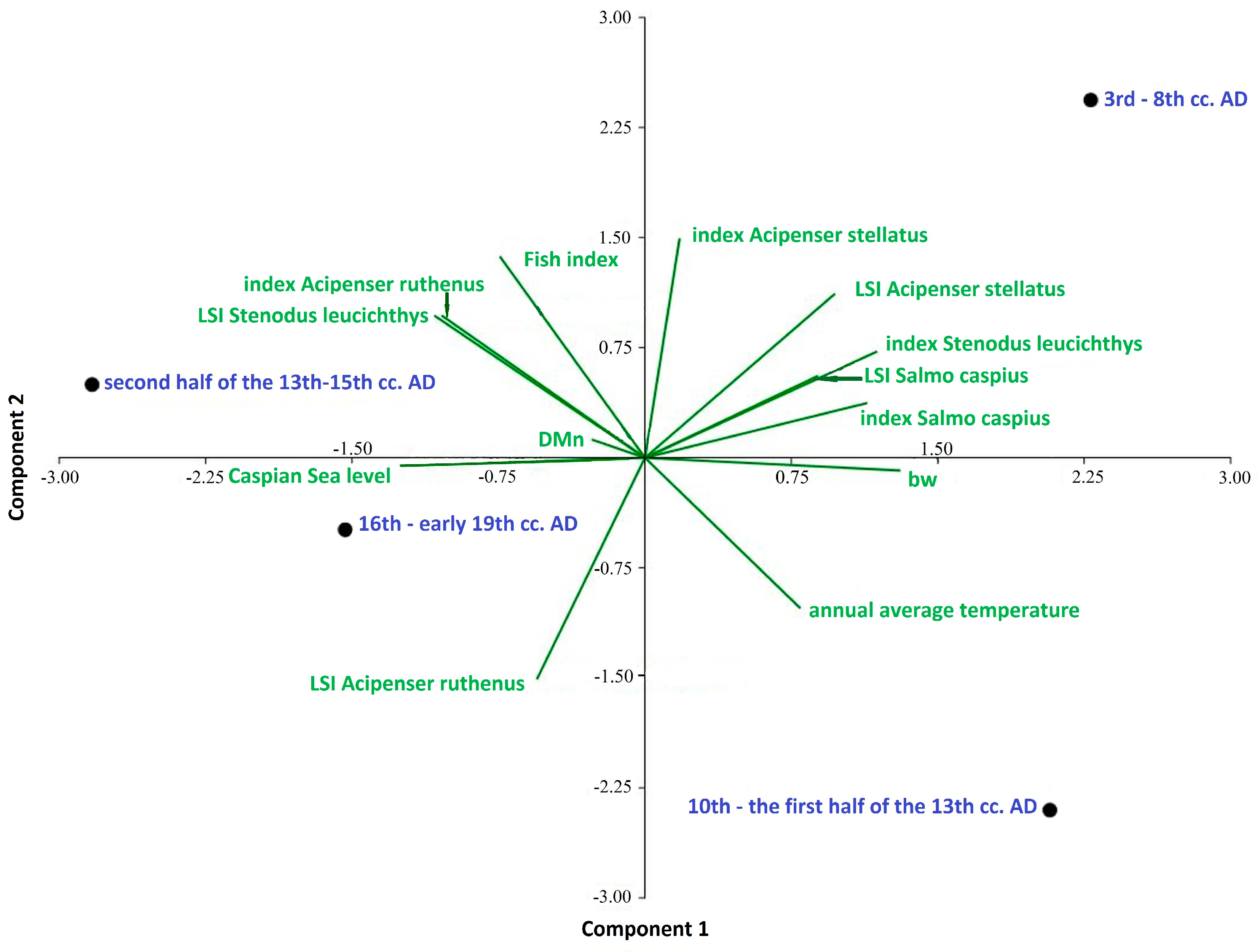
- Calculations of the Whittaker measure showed that the highest values were observed in the period of the 10th century to the first half of the 13th century, and the lowest in the second half of the 13th–15th centuries as well as in the 16th and early 19th centuries. This confirms that, where there are fewer common species in archaeoichthyological collections, their β-diversity is greater, and that there were quite large differences in the species composition of fish at archaeological sites within a certain period. This may indicate the fishing preferences in relation to species or groups of fish over a given period of time or a certain region.
- Menhínik species richness index values showed that, in the first three historical periods, commercial exploitation of fish stocks was approximately the same, but in the period of the 16th–early 19th century, it increased significantly due to an increase in the number of species and a change in the ratio of species in catches, as well as, in connection with this, an increase in the commercial exploitation of all species of fish.
- Historical stages in the development of society in the Volga region: 3rd–8th cc. AD, 10th–the first half of the 13th cc. AD, second half of the 13th–15th cc. AD, 16th–early 19th cc. AD. bw, Whittaker measure; DMn, index of species richness of Menhinik.
- Fish index, the ratio ΣMNI large fish >50 cm to ΣMNI large fish >50 cm + ΣMNI small fish < 50 cm.
- Index Acipenser stellatus, index Salmo caspius, index Stenodus leucichthys, index Acipenser ruthenus: species indices expressing the ratio of the number of remains of a species to the number of bone remains of other fish species.
- LSI Acipenser stellatus, LSI Salmo caspius, LSI Stenodus leucichthys, LSI Acipenser ruthenus: comparative fish size indices; annual average temperature and average annual temperature indicators; Caspian Sea level and changes in the level of the Caspian Sea.
4. Discussion
- The Volga river system was characterized by four species of sturgeon, wels catfish, zander, and common bream, with the inclusion of a large number of species of cyprinids, Northern pike, European perch, and Caspian inconnu. The Danube was characterized by common carp, northern pike, cyprinids, wels catfish, and sturgeons.
- A significant change in fishing ichthyofaunas in the Volga River system occurred in the Danube during the 19th century, at the end of the Middle Ages to the beginning of the modern period.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mineeva, N.; Lazareva, V.; Litvinov, A.; Stepanova, I.; Chuiko, G.; Papchenkov, V.; Korneva, L.; Shcherbina, G.; Pryanichnikova, E.; Perova, S.; et al. Chapter 2—The Volga River. In Rivers of Europe, 2nd ed.; Tockner, K., Zarfl, C., Robinson, C.T., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 27–79, 922. [Google Scholar]
- Schletterer, M.; Kuzovlev, V.V.; Zhenikov, Y.N.; Tuhtan, J.A.; Haidvogl, G.; Friedrich, T.; Górski, K.; Füreder, L. Fish fauna and fisheries of large European rivers: Examples from the Volga and the Danube. Hydrobiologia 2018, 814, 45–60. [Google Scholar] [CrossRef]
- Askeyev, I.V.; Askeyev, O.V.; Galimova, D.N. Natural environment and man in the Volga-Kama and Cis-Urals (Late Paleolithic—Middle Ages). In The Middle Volga and Southern Urals: Man and Nature in Antiquity. Collection of Scientific Articles Dedicated to the 75th Anniversary of Doctor of Historical Sciences Evgeny Petrovich Kazakov; Institute of History of the Academy of Sciences of the Republic of Tatarstan: Kazan, Russia, 2009; pp. 32–112. [Google Scholar]
- Askeyev, I.V.; Galimova, D.N.; Askeyev, O.V. Ichthyofauna of the middle Volga River basin in the Late Holocene (based on archaeological excavations). Zool. Zhurnal 2013, 92, 1014–1030. [Google Scholar] [CrossRef]
- Askeyev, I.V.; Askeyev, O.V.; Galimova, D.N. Archeo-ichthyological research on the territory of the Volga-Kama region. In Archeology and Natural Sciences of Tatarstan. Book 4; Institute of History of the Academy of Sciences of the Republic of Tatarstan: Kazan, Russia, 2011; pp. 44–156. [Google Scholar]
- Butler, V.L.; Delacorte, M.G. Doing Zooarchaeology as if it mattered: Use of faunal data to address current issues in fish conservation biology in Owens Valley, California. In Zooarchaeology and Conservation Biology; Lyman, R.L., Cannon, K.P., Eds.; University of Utah Press: Salt Lake City, UT, USA, 2004; pp. 25–44. [Google Scholar]
- Braje, T.J.; Rick, T.C.; Erlandson, J.M. Rockfish in the long view: Applied zooarchaeology and conservation of Pacific red snapper (genus Sebastes) in southern California. In Conservation Biology and Applied Zooarchaeology; Wolverton, S., Lyman, R.L., Eds.; University of Arizona Press: Tucson, AZ, USA, 2012; pp. 157–178. [Google Scholar]
- Speller, C.F.; Hauser, L.; Lepofsky, D.; Moore, J.; Rodrigues, A.T.; Moss, M.L.; McKechnie, I.; Yang, D.Y. High potential for using DNA from ancient herring bones to inform modern fisheries management and conservation. PLoS ONE 2012, 7, e51122. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.H. An environmental (pre)history of European fishing: Past and future archaeological contributions to sustainable fisheries. J. Fish Biol. 2019, 94, 1033–1044. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.; Gorin, S.; Fleming, N. The Archaeology Coursebook—An Introduction to Study Skills, Topics and Methods; Taylor & Francis Group: London, UK; New York, NY, USA, 2002; 323p. [Google Scholar]
- Piezonka, H.; Kostyleva, E.; Zhilin, M.G.; Dobrovolskaya, M.; Terberger, T. Flesh or fish? First results of archaeometric research of prehistoric burialsfrom Sakhtysh IIa, Upper Volga region, Russia. Doc. Praehist. 2013, 40, 57–73. [Google Scholar] [CrossRef]
- Jones, T.L.; Gobalet, K.W.; Codding, B.F. The archaeology of fish and fishing on the central coast of California: The case for an under-exploited resource. J. Anthr. Archaeol. 2016, 41, 88–108. [Google Scholar] [CrossRef]
- McKechnie, I.; Moss, M.L. Meta-analysis in zooarchaeology expands perspectives on Indigenous fisheries of the Northwest Coast of North America. J. Archaeol. Sci. Rep. 2016, 8, 470–485. [Google Scholar] [CrossRef]
- Shaymuratova, D.N.; Askeyev, I.V.; Askeyev, O.V.; Monachov, S.P.; Askeyev, A.O.; Smirnov, A.A. Sterlet Acipenser ruthenus (Acipenseriformes, Acipenseridae) of Middle Volga and Lower Kama in IV–XVIII centuries AD: Size and age composition, growth and value in the ancient fishing. Vopr. Rybolov. 2017, 18, 401–421. [Google Scholar]
- Guiry, E.J.; Kennedy, J.R.; O’connell, M.T.; Gray, D.R.; Grant, C.; Szpak, P. Early evidence for historical overfishing in the Gulf of Mexico. Sci. Adv. 2021, 7, eabh2525. [Google Scholar] [CrossRef]
- Lebedev, V.D. Freshwater Quaternary Ichthyofauna of the European Part of the USSR; Moscow State University Publishing: Moscow, Russia, 1960; 402p. [Google Scholar]
- Tsepkin, E.A. Changes in the exploited fish fauna of inland water-bodies of Eastern Europe and Northern Asia in the Quaternary period. Vopr. Ikhtiologii 1995, 35, 3–17. [Google Scholar]
- Makowiecki, D. History of Fishes and Fishing in Holocene on Polish Lowland in the Light of Archaeoichthyological Studies; Institute of Archaeology and Ethnology Polish Academy of Science: Poznan, Poland, 2003; 198p. [Google Scholar]
- Galik, A.; Haidvogl, G.; Bartosiewicz, L.; Guti, G.; Jungwirth, M. Fish remains as a source to reconstruct long-term changes of fish communities in the Austrian and Hungarian Danube. Aquat. Sci. 2015, 77, 337–354. [Google Scholar] [CrossRef] [PubMed]
- Broughton, J.M.; Martin, E.P.; McEneaney, B.; Wake, T.; Simons, D.D. Late Holocene anthropogenic depression of sturgeon in San Francisco Bay, California. J. Calif. Great Basin Anthropol. 2015, 35, 3–27. [Google Scholar]
- De Cupere, B.; Van Neer, W. Faunal Remains from Archaeological Sites Document Human Impact on the Terrestrial and Aquatic Environment: Examples from the last thousand years in Belgium. Internet Archaeol. 2023, 62. [Google Scholar] [CrossRef]
- Shaymuratova, D.N.; Askeyev, I.V.; Nedashkovsky, L.F. Archaeoichthyological research of settlements of the Golden Horde period of the Saratov Volga. Volga River Reg. Archaeol. 2021, 4, 191–204. [Google Scholar] [CrossRef]
- Casteel, R.W. Some archaeological uses of fish remains. Am. Antiq. 1972, 37, 404–419. [Google Scholar] [CrossRef]
- Morey, D.F. Archaeological assessment of seasonality from freshwater fish remains: Aquantitative procedure. J. Ethnobiol. 1983, 3, 75. [Google Scholar]
- Wheeler, A.; Jones, A.K.G. Fishes; Cambridge Manuals in Archaeology; Cambridge University Press: Cambridge, UK, 1989; 210p. [Google Scholar]
- Guillaud, E.; Elleboode, R.; Mahe, K.; Bearez, P. Estimation of age, growth and fishing season of a Paleolithic population of grayling (Thymallus thymallus) using scale analysis. Int. J. Osteoarchaeol. 2017, 27, 683–692. [Google Scholar] [CrossRef]
- Askeyev, I.V.; Tarasov, A.Y.; Askeyev, A.O.; Askeyev, O.V.; Shaymuratova, D.N.; Monakhov, S.P. Highly productive fishing in Lake Onega? New data on the subsistence basis of the Late Stone age populations in Russian Karelia. J. Archaeol. Sci. Rep. 2023, 47, 103771. [Google Scholar]
- Morales Muñiz, A.; Rosenlund, K. Fish Bone Measurements: An Attempt to Standardize the Measurement of Fish Bones from Archaeological Sites; Steenstrupia: Copenhagen, Denamrk, 1979; 48p. [Google Scholar]
- Radu, V. Exploitation des Ressources Aquatiques dans les Cultures Néolithiques et Calcolithiques de la Roumanie Méridionale. Ph.D. Thesis, l’Université Aix-Marseille I, Préhistoire, Archéologie, Histoire et Civilisation de l’Antiquité et du Moyen-Age, Aix-en-Provence, France, 2003; 432p. [Google Scholar]
- Živaljević, I.; Askeyev, I.V.; Shaymuratova (Galimova), D.N.; Askeyev, O.V.; Monakhov, S.P.; Borić, D.; Stefanović, S. Size estimations of sturgeons (Acipenseridae) from the Mesolithic-Neolithic Danube Gorges. In Foraging Assemblages, Volume 2; Borić, D., Antonović, D., Mihailović, B., Eds.; Serbian Archaeological Society: Belgrade, Serbia; The Italian Academy for Advanced Studies in America, Columbia University: New York, NY, USA, 2021; pp. 422–427. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentince-Hall: Englewood Cliffs, NJ, USA, 1999; 663p. [Google Scholar]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Chugunova, N.I. Guidebook for the Study of Fish Age and Growth; Izd. Akademii nauk SSSR: Moscow, Russia, 1959; 164p. [Google Scholar]
- Pravdin, I.F. Guide to the Study of Fish; Pischevaya Promyshlennost: Moscow, Russia, 1966; 376p. [Google Scholar]
- Panfili, J.; de Pontual, H.; Troadec, H.; Wright, P.J. (Eds.) Manual of Fish Sclerochronology; Ifremer-lRD Coedition: Brest, France, 2002; 464p. [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Science: Hoboken, NJ, USA, 2004; 256p. [Google Scholar]
- Whittaker, R.H. Evolution of species diversity in land communities. Evol. Biol. 1977, 10, 1–67. [Google Scholar]
- Butler, V.L. Resource depression on the Northwest Coast of North America. Antiquity 2000, 74, 649–661. [Google Scholar] [CrossRef]
- Sokolov, L.I.; Tsepkin, E.A. Sterlet Acipenser ruthenus L. in middle and late Holocene. Bull. Moscow Soc. Nat. Biol. Ser. 1971, 76, 137–145. [Google Scholar]
- Borzenko, M.P. Caspian stellate sturgeon (systematics, biology and fishing). Izv. Azerbaijan Sci. Res. Fish. Stn. 1942, 7, 3–114. [Google Scholar]
- Berg, L.S. Chapter 1. In Freshwater Fish of the USSR and Neighboring Countries; Izd. Akademii nauk SSSR: Moscow-Leningrad, Russia, 1948; 466p. [Google Scholar]
- Shmidtov, A.I. Sterlet (Acipenser ruthenus L.). Materials on the Biology and Fishing of Sterlet in the Lower Reaches of the Kama River; Scientific notes of Kazan University; Zoology; Volume 99; Book 4; Kazan University: Kazan, Russia, 1939; pp. 13–279. [Google Scholar]
- Derzhavin, A.N. Volga Salmon (Based on Historical Materials); Collection Articles in Honor of N.M. Knipovich; Pishchepromizdat: Moscow, Russia; Leningrad, Russia, 1939; pp. 187–206. [Google Scholar]
- Klimenko, V.; Solomina, O. Climatic variations in the East European Plain during the last Millennium: State of the art. In The Polish Climate in the European Context: An Historical Overview; Przybylak, R., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 71–101. [Google Scholar] [CrossRef]
- Bolgov, M.V.; Krasnozhon, G.F.; Lyubushin, A.A. Caspian Sea: Extreme Hydrological Events; Nauka: Moscow, Russia, 2007; 381p. [Google Scholar]
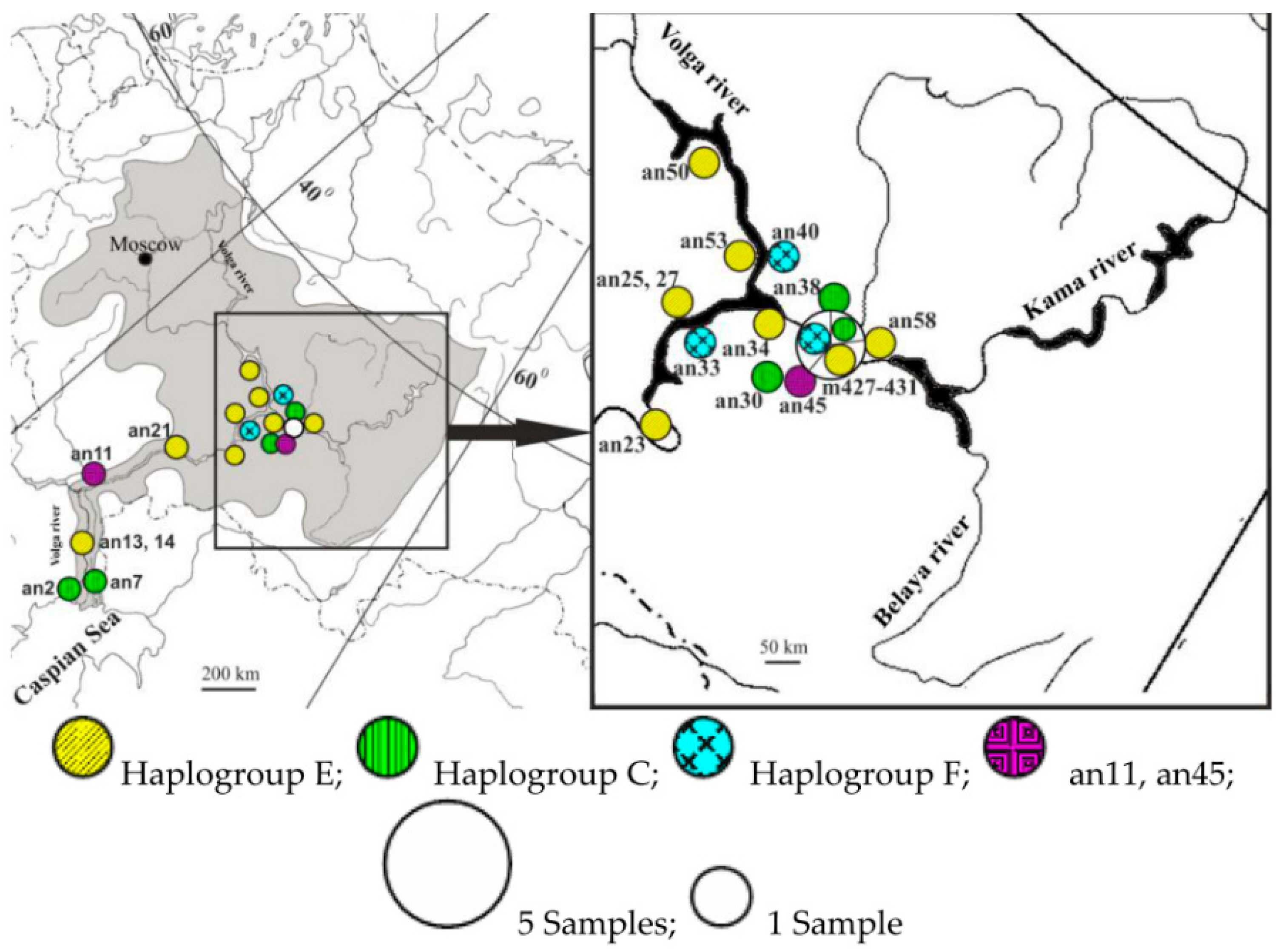
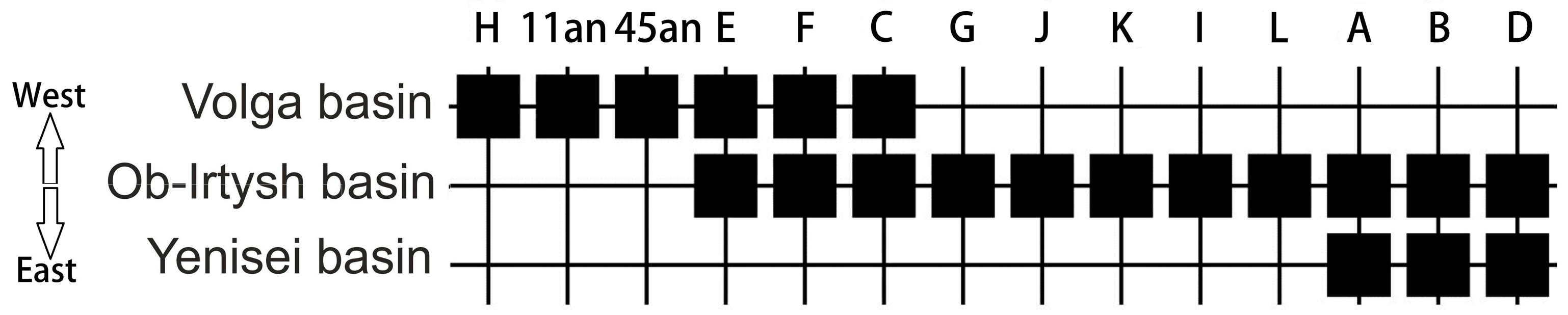
- Pobedintseva, M.A.; Makunin, A.I.; Kichigin, I.G.; Kulemzina, A.I.; Serdyukova, N.A.; Romanenko, S.A.; Vorobieva, N.V.; Interesova, E.A.; Korentovich, M.A.; Zaytsev, V.F.; et al. Population genetic structure and phylogeography of sterlet (Acipenser ruthenus, Acipenseridae) in the Ob and Yenisei river basins. Mitochondrial DNA Part A 2019, 30, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.Y.; Eng, B.; Waye, J.S.; Dudar, J.C.; Saunders, S.R. Technical Note: Improved DNA Extraction From Ancient Bones Using Silica-Based Spin Columns. Am. J. Phys. Anthropol. 1998, 105, 539–543. [Google Scholar] [CrossRef]
- Sanderson, C.; Radley, K.; Mayton, L. Ethylenediaminetetraacetic acid in ammonium hydroxide for reducing decalcification time. Biotech. Histochem. 1995, 70, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Cheng, L.; Li, J.-T.; Lu, C.Y.; Wang, Y.; Sun, X.W. Complete mitochondrial genome of sterlet (Acipenser ruthenus). Mitochondrial DNA 2013, 26, 259–260. [Google Scholar] [CrossRef]
- Dabney, J.; Meyer, M.; Pääbo, S. Ancient DNA damage. Cold Spring Harb Perspect Biol. 2013, 5, a012567. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 150–3152. [Google Scholar] [CrossRef] [PubMed]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; Ugene Team. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [PubMed]
- Clement, M.; Snell, Q.; Walker, P.; Posada, D.; Crandall, K. TCS: Estimating gene genealogies. Parallel Distributed Process. Symp. Int. 2002, 3, 184–190. [Google Scholar]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods. Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Kessler, K.F. Ichthyofauna of the Volga River. Proc. St. Petersburg Soc. Nat. 1870, 1, 236–311. [Google Scholar]
- Butorin, N.V. Volga and Its Life; Nauka: Leningrad, Russia, 1978; 348p. [Google Scholar]
- Schletterer, M.; Shaporenko, S.I.; Kuzovlev, V.V.; Minin, A.E.; Van Geest, G.J.; Middelkoop, H.; Górski, K. The Volga: Management issues in the largest river basin in Europe. River Res. Appl. 2019, 35, 510–519. [Google Scholar] [CrossRef]
- Pallas, P.S. Travels through Various Provinces of the Russian Empire, 3 Volumes (Vol. 1, 504 pp., Vol. 2, 744 pp., Vol. 3, 760 pp. + 118 plates) 1771–1776; Akademische Druck und Verlagsanstalt: Graz, Austria, 1967. [Google Scholar]
- Lepekhin, I.I. Day Notes of a Trip to Different Provinces of the Russian State. 1768 and 1769; Imperatorskaya Akademiya Nauk: St. Petersburg, Russia, 1771; 538p. [Google Scholar]
- van de Wolfshaar, K.E.; Middelkoop, H.; Addink, E.; Winter, H.V.; Nagelkerke, L.A.J. Linking Flow Regime, Floodplain Lake Connectivity and Fish Catch in a Large River-Floodplain System, the Volga–Akhtuba Floodplain (Russian Federation). Ecosystems 2011, 14, 920–934. [Google Scholar] [CrossRef]
- Górski, K.; van den Bosch, L.V.; van de Wolfshaar, K.E.; Middelkoop, H.; Nagelkerke, L.A.J.; Filippov, O.V.; Zolotarev, D.V.; Yakovlev, S.V.; Minin, A.E.; Winter, H.V.; et al. Post-damming flow regime development in a large lowland river (Volga, Russian Federation): Implications for floodplain inundation and fisheries. River Res. Appl. 2012, 28, 1121–1134. [Google Scholar] [CrossRef]
- Samoylenko, V.V. Dynamics, structure and drivers of fish catch in Russian Federation (2005–2017). Izv. TINRO 2019, 196, 204–218. [Google Scholar] [CrossRef]
- Ponomareva, E.N.; Balykin, P.A.; Startsev, A.V.; Korchunov, A.A.; Savitskaya, S.S. Current state of fisheries in the Volga-Caspian subarea. Vestnik Astrakhan State Tech. Univ. Series: Fish. Ind. 2022, 7–15. [Google Scholar] [CrossRef]
- Maltsev, S.A. Conservation of the Sturgeon Fish in Lower Volga. In Biology, Conservation and Sustainable Development of Sturgeons; Carmona, R., Domezain, A., García-Gallego, M., Hernando, J.A., Rodríguez, F., Ruiz-Rejón, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 265–273. [Google Scholar]
- Altukhov, Y.P.; Evsyukov, A.N. Juvenile Overproduction at Fish Hatcheries May Explain the Degradation of the Volga Stock of the Russian Sturgeon. Dokl. Biol. Sci. 2001, 380, 454–456. [Google Scholar] [CrossRef]
- Górski, K.; Winter, H.V.; DE Leeuw, J.J.; Minin, A.E.; Nagelkerke, L.A.J. Fish spawning in a large temperate floodplain: The role of flooding and temperature. Freshw. Biol. 2010, 55, 1509–1519. [Google Scholar] [CrossRef]
- Górski, K.; Buijse, A.D.; Winter, H.V.; De Leeuw, J.J.; Compton, T.J.; Vekhov, D.A.; Zolotarev, D.V.; Verreth, J.A.J.; Nagelkerke, L.A.J. Geomorphology and Flooding Shape Fish Distribution in A Large-Scale Temperate Floodplain. River Res. Appl. 2012, 29, 1226–1236. [Google Scholar] [CrossRef]
- Artaev, O.; Ruchin, A.; Ivanchev, V.; Ivancheva, E.; Sarychev, V.; Moreva, O.; Mikheev, V.; Medvedev, D.; Klevakin, A. Fish occurrence in the middle Volga and upper Don regions (Russia). Biodivers. Data J. 2020, 8, e54959. [Google Scholar] [CrossRef] [PubMed]
- Askeyev, O.V.; Monakhov, S.P.; Askeyev, I.V.; Askeyev, A.O.; Sparks, T.H. Fish assemblages in lakes along environmental gradients at the eastern edge of Europe. Environ. Biol. Fish. 2023, 106, 1265–1276. [Google Scholar] [CrossRef]
- Askeyev, O.; Askeyev, I.; Askeyev, A.; Monakhov, S.; Yanybaev, N. River fish assemblages in relation to environmental factors in the eastern extremity of Europe (Tatarstan Republic, Russia). Environ. Biol. Fishes 2014, 98, 1277–1293. [Google Scholar] [CrossRef]
- Askeyev, A.O.; Askeyev, O.V.; Yanybaev, N.M.; Askeyev, I.V.; Monakhov, S.P.; Marić, S.; Hulsman, K. River fish assemblages along an elevation gradient in the eastern extremity of Europe. Environ. Biol. Fishes 2017, 100, 585–596. [Google Scholar] [CrossRef]
- Askeyev, A.O.; Askeyev, O.V.; Askeyev, I.V.; Monakhov, S.P. Predatory fish species as indicators of biodiversity: Their distribution in environmental gradients in small and mid-sized rivers in Eastern Europe. Environ. Biol. Fishes 2021, 104, 767–778. [Google Scholar] [CrossRef]
- Marić, S.; Askeyev, O.V.; Askeyev, A.O.; Monakhov, S.P.; Yanybaev, N.M.; Askeyev, I.V.; Galimova, D.N.; Snoj, A. Lack of mtDNA variation among remote middle Volga and upper Ural brown trout suggests recent and rapid recolonization. J. Appl. Ichthyol. 2016, 32, 948–953. [Google Scholar] [CrossRef]
- Marić, S.; Alekseyev, S.; Snoj, A.; Askeyev, O.; Askeyev, I.; Weiss, S. First mtDNA sequencing of Volga and Ob basin taimen Hucho taimen: European populations stem from a late Pleistocene expansion of H. taimen out of western Siberia and are not intermediate to Hucho hucho. J. Fish Biol. 2014, 85, 530–539. [Google Scholar] [CrossRef]
- Lecaudey, L.A.; Schletterer, M.; Kuzovlev, V.V.; Hahn, C.; Weiss, S.J. Fish diversity in the headwaters of the Volga River using environmental DNA metabarcoding. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 1785–1800. [Google Scholar] [CrossRef]
- Schenekar, T.; Schletterer, M.; Lecaudey, L.A.; Weiss, S.J. Reference databases, primer choice, and assay sensitivity for environmental metabarcoding: Lessons learnt from a re-evaluation of an eDNA fish assessment in the Volga headwaters. River Res. Appl. 2020, 36, 1004–1013. [Google Scholar] [CrossRef]
- Marić, S.; Askeyev, I.V.; Askeyev, O.V.; Monakhov, S.P.; Bravničar, J.; Snoj, A. Phylogenetic and population genetic analysis of Thymallus thymallus (Actinopterygii, Salmonidae) from the middle Volga and upper Ural drainages. Hydrobiologia 2014, 740, 167–176. [Google Scholar] [CrossRef]
- Haponski, A.E.; Stepien, C.A. Phylogenetic and biogeographical relationships of the Sander pikeperches (Percidae: Perciformes): Pat terns across North America and Eurasia. Biol. J. Linn. Soc. 2013, 110, 156–179. [Google Scholar] [CrossRef]
- Triantafyllidis, A.; Krieg, F.; Cottin, C.; Abatzopoulos, T.J.; Triantaphyllidis, C.; Guyomard, R. Genetic structure and phylogeography of European catfish (Silurus glanis) populations. Mol. Ecol. 2002, 11, 1039–1055. [Google Scholar] [CrossRef] [PubMed]
- Van Houdt, J.K.J.; De Cleyn, L.; Perretti, A.; Volckaert, F.A.M. A mitogenic view on the evolutionary history of the Holarctic freshwater gadoid, burbot (Lota lota). Mol. Ecol. 2005, 14, 2445–2457. [Google Scholar] [CrossRef] [PubMed]
- Rastorguev, S.M.; Nedoluzhko, A.V.; Mazur, A.M.; Gruzdeva, N.M.; Volkov, A.A.; Barmintseva, A.E.; Mugue, N.S.; Prokhortchouk, E.B. High-throughput SNP-genotyping analysis of the relationships among Ponto-Caspian sturgeon species. Ecol. Evol. 2013, 3, 2612–2618. [Google Scholar] [CrossRef] [PubMed]
- Perdices, A.; Vasil’eva, E.; Vasil’ev, V. From Asia to Europe across Siberia: Phylogeography of the Siberian spined loach (Teleostei, Cobitidae). Zool. Scr. 2015, 44, 29–40. [Google Scholar] [CrossRef]
- Levin, B.A.; Simonov, E.P.; Ermakov, O.A.; Levina, M.A.; Interesova, E.A.; Kovalchuk, O.M.; Malinina, Y.A.; Mamilov, N.S.; Mustafayev, N.J.; Pilin, D.V.; et al. Phylogeny and phylogeography of the roaches, genus Rutilus (Cyprinidae), at the Eastern part of its range as inferred from mtDNA analysis. Hydrobiologia 2016, 788, 33–46. [Google Scholar] [CrossRef]
- Artaev, O.N.; Ermakov, O.A.; Vekhov, D.A.; Konovalov, A.F.; Levina, M.A.; Pozdeev, I.V.; Ruchin, A.B.; Alyushin, I.V.; Iljin, V.Y.; Levin, B.A. Genetic Screening of Distribution Pattern of Roaches Rutilus rutilus and R. lacustris (Cyprinidae) in Broad Range of Secondary Contact (Volga Basin). Inland Water Biol. 2021, 14, 205–214. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species (n = 41 + 3 Hybrids) | Number of Archaeozoological Collections (n = 32) | NISPbones | NISPscales | MNI |
|---|---|---|---|---|
| Russian sturgeon, Acipenser gueldenstaedtii | 26 | 2609 | 603 | |
| Fringebarbel sturgeon, Acipenser nudiventris | 8 | 13 | 11 | |
| Sterlet, Acipenser ruthenus | 26 | 3864 | 1524 | |
| Starry sturgeon, Acipenser stellatus | 26 | 2771 | 465 | |
| Beluga, Huso huso | 22 | 1180 | 361 | |
| Hybrid, Acipenser gueldenstaedtii × Acipenser ruthenus | 1 | 1 | 1 | |
| Hybrid, Acipenser ruthenus × Acipenser stellatus | 1 | 1 | 1 | |
| Caspian anadromous shad, Alosa kessleri | 13 | 155 | 368 | 84 |
| Volga shad, Alosa volgensis | 2 | 4 | 32 | 8 |
| Caspian shad, Alosa caspia | 1 | 2 | 1 | |
| Aral barbel, Luciobarbus brachycephalus caspius | 2 | 5 | 2 | |
| Bulatmai barbel, Luciobarbus capito capito | 2 | 4 | 3 | |
| Crucian carp, Carassius carassius | 14 | 196 | 207 | 82 |
| Common carp, Cyprinus carpio | 20 | 436 | 63 | 135 |
| Common bream, Abramis brama | 28 | 1531 | 4449 | 819 |
| Blue bream, Ballerus ballerus | 16 | 109 | 139 | 61 |
| White-eye bream, Ballerus sapa | 6 | 52 | 24 | 29 |
| White bream, Blicca bjoerkna | 16 | 304 | 340 | 99 |
| Caspian vimba, Vimba persa | 1 | 1 | 1 | |
| Common bleak, Alburnus alburnus | 5 | 369 | 76 | 64 |
| Danube bleak, Alburnus chalcoides | 2 | 8 | 4 | 5 |
| Asp, Aspius aspius | 16 | 103 | 26 | 57 |
| Common dace, Leuciscus leuciscus | 1 | 8 | 5 | |
| Ide, Leuciscus idus | 14 | 109 | 60 | 54 |
| Volga undermouth, Chondrostoma variabile | 7 | 27 | 12 | |
| Kutum, Rutilus kutum | 10 | 32 | 20 | |
| Roach, Rutilus rutilus | 18 | 536 | 1424 | 313 |
| Rudd, Scardinius erythrophthalmus | 6 | 26 | 15 | 10 |
| Chub, Squalis cephalus | 9 | 28 | 20 | |
| Sichel, Pelecus cultratus | 9 | 102 | 216 | 41 |
| Tench, Tinca tinca | 12 | 84 | 35 | |
| Hybrid, Abramis brama × Rutilus rutilus | 1 | 1 | 1 | |
| Wels catfish, Silurus glanis | 29 | 878 | 346 | |
| Northern pike, Esox lucius | 30 | 716 | 195 | 454 |
| Pereslavl lake vendace, Coregonus albula pereslavicus | 1 | 200 | 11 | 40 |
| Caspian Inconnu, Stenodus leucichthys leucichthys | 17 | 445 | 13 | 173 |
| Grayling, Thymallus thymallus | 1 | 9 | 2 | |
| Siberian taimen, Hucho taimen | 2 | 5 | 3 | |
| Caspian trout, Salmo caspius | 13 | 120 | 70 | |
| Burbot, Lota lota | 7 | 30 | 15 | |
| Ruffe, Gymnocephalus cernuus | 5 | 109 | 68 | 32 |
| European perch, Perca fluviatilis | 17 | 835 | 2685 | 352 |
| Zander, Sander lucioperca | 28 | 1667 | 192 | 799 |
| Volga zander, Sander volgensis | 7 | 14 | 6 | 13 |
| 3rd–8th cc. AD | 10th–1st Half of the 13th cc. AD | 2nd Half of the 13th–15th cc. AD | 16th–Early 19th cc. AD | |
|---|---|---|---|---|
| Index Stenodus leucichthys | 0.055 | 0.033 | 0.011 | 0.012 |
| Index Salmo caspius | 0.016 | 0.012 | 0.004 | 0.011 |
| Index Acipenser stellatus | 0.087 | 0.058 | 0.077 | 0.056 |
| Index Acipenser ruthenus | 0.271 | 0.137 | 0.406 | 0.276 |
| LSI Acipenser stellatus (n = 350) | 0.075 | 0.041 | 0.030 | 0.038 |
| LSI Acipenser ruthenus (n = 1268) | 0.086 | 0.149 | 0.132 | 0.150 |
| LSI Salmo caspius (n = 53) | 0.009 | −0.027 | −0.065 | −0.01 |
| LSI Stenodus leucichthys (n = 95) | −0.023 | −0.037 | −0.013 | −0.018 |
| Caspian Sea level | −29.224 | −29.217 | −26.316 | −25.796 |
| Annual average temperature | 3.16 | 3.54 | 3.15 | 3.0 |
| (bw) | 1.453 | 1.4823 | 1.000 | 1.0488 |
| (DMn) | 0.8214 | 0.7093 | 0.6918 | 1.095 |
| Fish index | 0.7485 | 0.5622 | 0.7972 | 0.6928 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Askeyev, I.V.; Askeyev, O.V.; Askeyev, A.O.; Shaymuratova, D.N.; Monakhov, S.P.; Pobedintseva, M.A.; Trifonov, V.A.; Górski, K.; Schletterer, M. Archaeozoology Supports a Holistic View on Fish Assessments in Large Rivers—A Case Study from the Volga River: From Quantitative Data and Ancient DNA to Biodiversity Analysis. Water 2024, 16, 1109. https://doi.org/10.3390/w16081109
Askeyev IV, Askeyev OV, Askeyev AO, Shaymuratova DN, Monakhov SP, Pobedintseva MA, Trifonov VA, Górski K, Schletterer M. Archaeozoology Supports a Holistic View on Fish Assessments in Large Rivers—A Case Study from the Volga River: From Quantitative Data and Ancient DNA to Biodiversity Analysis. Water. 2024; 16(8):1109. https://doi.org/10.3390/w16081109
Chicago/Turabian StyleAskeyev, Igor V., Oleg V. Askeyev, Arthur O. Askeyev, Dilyara N. Shaymuratova, Sergey P. Monakhov, Maria A. Pobedintseva, Vladimir A. Trifonov, Konrad Górski, and Martin Schletterer. 2024. "Archaeozoology Supports a Holistic View on Fish Assessments in Large Rivers—A Case Study from the Volga River: From Quantitative Data and Ancient DNA to Biodiversity Analysis" Water 16, no. 8: 1109. https://doi.org/10.3390/w16081109
APA StyleAskeyev, I. V., Askeyev, O. V., Askeyev, A. O., Shaymuratova, D. N., Monakhov, S. P., Pobedintseva, M. A., Trifonov, V. A., Górski, K., & Schletterer, M. (2024). Archaeozoology Supports a Holistic View on Fish Assessments in Large Rivers—A Case Study from the Volga River: From Quantitative Data and Ancient DNA to Biodiversity Analysis. Water, 16(8), 1109. https://doi.org/10.3390/w16081109