
Genetic and Morphological Characterization of the Invasive Corbicula Lineages in European Russia
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Field Sampling
2.2. Morphological and Anatomical Studies
2.3. DNA Extraction, PCR Amplification, and DNA Sequencing
2.4. Sequence Alignment and Phylogeographic Analyses
3. Results
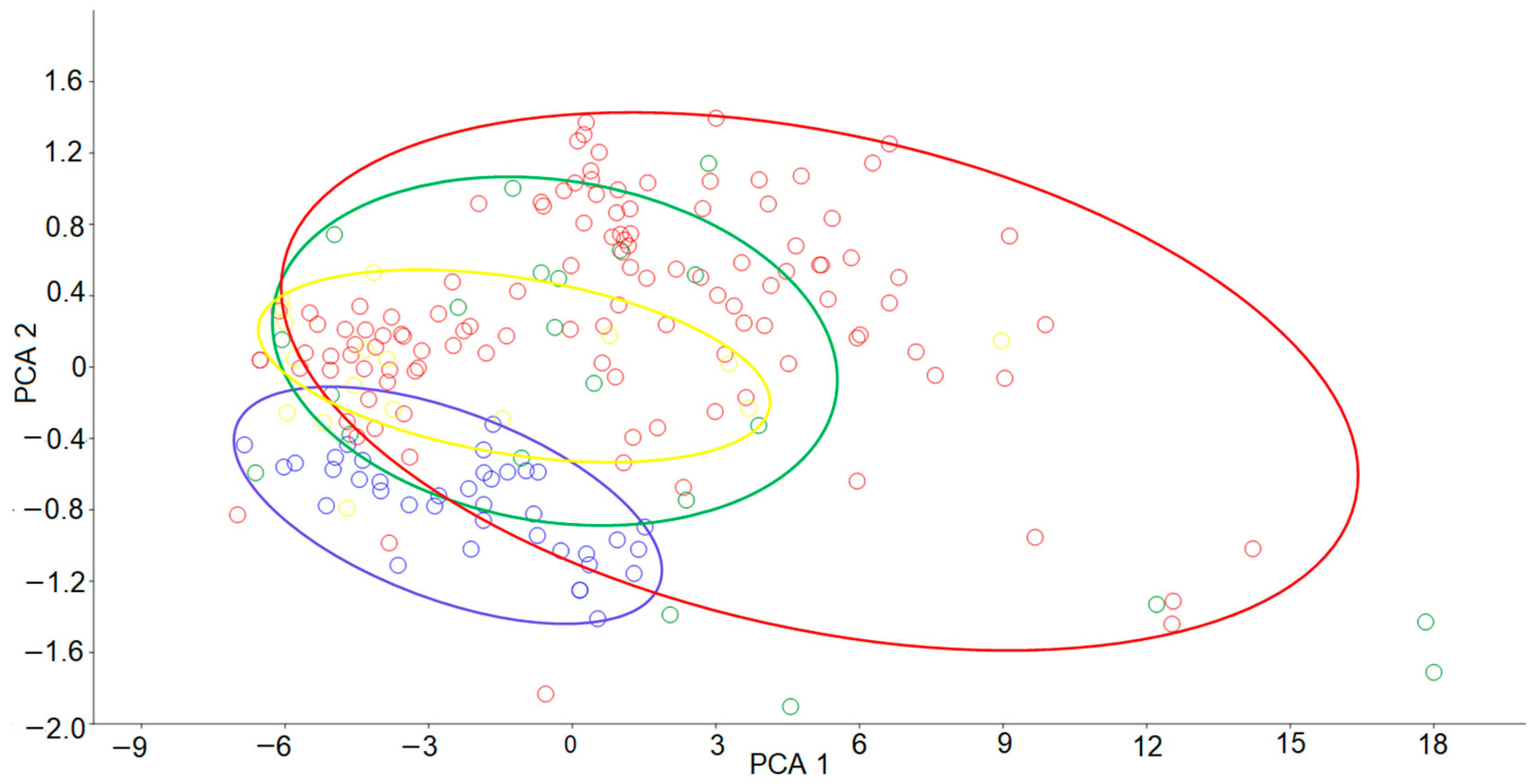
3.1. Shell Morphology and Anatomy of Clams
3.2. Mitochondrial DNA Sequences
3.3. Nuclear DNA Sequence Polymorphism
4. Discussion
4.1. Genetic and Morphological Analysis of Corbicula Lineages from European Russia
4.2. The 28S rRNA Polymorphism
4.3. Dispersal Routes of the Invasive Corbicula Lineages to Aquatic Ecosystems of European Russia
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Araujo, R.; Moreno, D.; Ramos, M.A. The Asiatic clam Corbicula fluminea (Meuller, 1774) (Bivalvia: Corbiculidae) in Europe. Am. Malacol. Bull. 1993, 10, 39–49. [Google Scholar]
- Pigneur, L.-M.; Etoundi, E.; Aldridge, D.C.; Marescaux, J.; Yasuda, N.; Van Doninck, K. Genetic uniformity and long-distance clonal dispersal in the invasive androgenetic Corbicula clams. Mol. Ecol. 2014, 23, 5102–5116. [Google Scholar] [CrossRef]
- Mouthon, J. Sur la pre’sence en France et au Portugal de Corbicula (Bivalvia, Corbiculidae) originaire d’Asie. Basteria 1981, 45, 109–116. [Google Scholar]
- Crespo, D.; Dolbeth, M.; Leston, S.; Sousa, R.; Pardal, A. Distribution of Corbicula fluminea (Müller, 1774) in the invaded range: A geographic approach with notes on species traits variability. Biol. Invasions 2015, 17, 2087–2101. [Google Scholar] [CrossRef]
- Gomes, C.; Sousa, R.; Mendes, T.; Borges, R.; Vilares, P.; Vasconcelos, V.; Guilhermino, L.; Antunes, A. Low genetic diversity and high invasion success of Corbicula fluminea (Bivalvia, Corbiculidae) (Müller, 1774) in Portugal. PLoS ONE 2016, 11, e0158108. [Google Scholar] [CrossRef] [PubMed]
- Penãrrubia, L.; Araguas, R.M.; Vidal, O.; Pla, C.; Vinãs, J.; Sanz, N. Genetic characterization of the Asian clam species complex (Corbicula) invasion in the Iberian Peninsula. Hydrobiologia 2017, 784, 349–365. [Google Scholar] [CrossRef]
- Bespalaya, Y.V.; Bolotov, I.N.; Aksenova, O.V.; Kondakov, A.V.; Gofarov, M.Y.; Laenko, E.; Sokolova, S.E.; Shevchenko, A.R.; Travina, O.V. Aliens are moving to the Arctic frontiers: An integrative approach reveals selective expansion of androgenic hybrid Corbicula lineages towards the North of Russia. Biol. Invasions 2018, 20, 2227–2243. [Google Scholar] [CrossRef]
- Bespalaya, Y.V.; Aksenova, O.V.; Kropotin, A.V.; Shevchenko, A.R.; Travina, O.V. Reproduction of the androgenetic population of the Asian Corbicula clam (Bivalvia: Cyrenidae) in the Northern Dvina River basin, Russia. Diversity 2021, 13, 316. [Google Scholar] [CrossRef]
- Nabozhenko, M.V.; Nabozhenko, S.V. Corbicula fluminalis (O.F. Müller, 1774), A new clam species for the Russian Sector of the Caspian Basin. Sci. South Russ. 2016, 12, 61–64. [Google Scholar]
- Khlopkova, M.V.; Guseynov, K.M.; Gasanova, A.S.; Barkhalov, R.M.; Zurkhaeva, U.D.; Guseynov, M.K. Russian first record of live clams of Corbicula fluminalis (Bivalvia: Corbiculidae) in the Dagestan Sector of the Caspian Sea Basin. Russ. J. Biol. Invasions 2019, 10, 79–82. [Google Scholar] [CrossRef]
- Khlopkova, M.V.; Barkhalov, R.M.; Guseynov, K.M.; Gasanova, A.S.; Zurkhaeva, U.D. Invasions of new morphotypes of Corbicula fluminalis and Corbicula fluminea (Bivalvia: Corbiculidae) to the basin of the Dagestan Region of the Caspian Sea. Russ. J. Biol. Invasions 2023, 14, 251–261. [Google Scholar] [CrossRef]
- Pryanichnikova, E.G.; Voroshilova, I.S.; Sabitova, R.Z. Introduction of Corbicula fluminea (Müller, 1774) (Mollusca: Bivalvia: Corbiculidae) in the Volga River Basin. Inland Water Biol. 2019, 12, 95–97. [Google Scholar] [CrossRef]
- Zhivoglyadova, L.A.; Revkov, N.K. First records of Corbicula fluminea (O.F. Müller, 1774) (Mollusca: Bivalvia) from the Lower Don. Ecol. Montenegrina 2018, 17, 46–52. [Google Scholar] [CrossRef]
- Zhivoglyadova, L.A.; Revkov, N.K. Invasion of the bivalve Corbicula fluminea (O.F. Müller, 1774) (Bivalvia: Cyrenidae) into the lower Don basin. Aquat. Bioresour. Environ. 2018, 1, 44–50. [Google Scholar] [CrossRef]
- Zhivoglyadova, L.A.; Revkov, N.K.; Kovalev, E.A. Extension of the bivalve Corbicula fluminea (O. F. MÜLLER, 1774) areal in the lower Don River system. Mar. Biol. J. 2018, 3, 73–75. [Google Scholar]
- Pigneur, L.M.; Hedtke, S.M.; Etoundi, E.; Van Doninck, K. Androgenesis: A review through the study of the selfish shellfish Corbicula spp. Heredity 2012, 108, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.; Siripattrawan, S.; Ituarte, C.F.; Foighil, D.O. Invasion of the clonal clams: Corbicula lineages in the New World. Am. Malacol. Bull. 2005, 20, 113–122. [Google Scholar]
- Hedtke, S.M.; Stanger-Hall, K.; Baker, R.J.; Hillis, D.M. All male asexuality: Origin and maintenance of androgenesis in the Asian clam Corbicula. Evolution 2008, 62, 1119–1136. [Google Scholar] [CrossRef]
- Hedtke, S.M.; Glaubrecht, M.; Hillis, D.M. Rare gene capture in predominantly androgenetic species. Proc. Natl. Acad. Sci. USA 2011, 108, 9520–9524. [Google Scholar] [CrossRef]
- Bespalaya, Y.V.; Aksenova, O.V.; Gofarov, M.Y.; Kondakov, A.V.; Kropotin, A.V.; Kononov, O.D.; Bolotov, I.N. Who inhabits the world’s deepest crater lake? A taxonomic review of Corbicula (Bivalvia: Cyrenidae) clams from Lake Toba, North Sumatra, Indonesia. J. Zool. Syst. Evol. Res. 2021, 59, 400–410. [Google Scholar] [CrossRef]
- Bespalaya, Y.V.; Kropotin, A.V.; Kondakov, A.V.; Aksenova, O.V.; Gofarov, M.Y.; Kim, S.K.; Lee, J.H.; Travina, O.V.; Vikhrev, I.V.; Vinarski, M.V.; et al. A taxonomic reassessment of native and invasive species of Corbicula clams (Bivalvia: Cyrenidae) from the Russian Far East and Korea. Zool. J. Linn. Soc. 2023, 197, 104–126. [Google Scholar] [CrossRef]
- Sheehan, R.; Etoundi, E.; Minchin, D.; Doninck, K.V.; Lucy, F. Identification of the invasive form of Corbicula clams in Ireland. Water 2019, 11, 1652. [Google Scholar] [CrossRef]
- Modesto, V.; Ilarri, M.; Labecka, A.M.; Rodríguez, N.F.; Coughlan, N.E.; Liu, X.; Sousa, R. What we know and do not know about the invasive Asian clam Corbicula fluminea. Hydrobiologia 2023. [Google Scholar] [CrossRef]
- Tiemann, J.S.; Haponski, A.E.; Douglass, S.A.; Lee, T.; Cummings, K.S.; Davis, M.A.; Ó Foighil, D. First record of a putative novel invasive Corbicula lineage discovered in the Illinois River, Illinois, USA. BioInvasions Rec. 2017, 6, 159–166. [Google Scholar] [CrossRef]
- Korniushin, A.V.; Glaubrecht, M. Novel reproductive modes in freshwater clams: Brooding and larval morphology in Southeast Asian taxa of Corbicula (Mollusca, Bivalvia, Corbiculidae). Acta Zool. 2003, 84, 293–315. [Google Scholar] [CrossRef]
- Kropotin, A.V.; Bespalaya, Y.V.; Aksenova, O.V.; Bolotov, I.N. Reproductive Mode of Corbicula tobae (Martens, 1900): Brooding and larval morphology in lake Toba (Indonesia). Diversity 2022, 14, 700. [Google Scholar] [CrossRef]
- Voroshilova, I.S.; Pryanichnikova, E.G.; Prokin, A.A.; Sabitova, R.Z.; Karabanow, D.P.; Pavlov, D.D.; Kurina, E.M. Morphological and genetic traits of the first invasive population of the Asiatic clam Corbicula fluminea (O.F. Müller, 1774) naturalized in the Volga River basin. Russ. J. Biol. Invasions 2021, 12, 36–43. [Google Scholar] [CrossRef]
- Pfenninger, M.; Reinhardt, F.; Streit, B. Evidence for cryptic hybridization between different evolutionary lineages of the invasive clam genus Corbicula (Veneroida, Bivalvia). J. Evol. Biol. 2002, 15, 818–829. [Google Scholar] [CrossRef]
- Korniushin, A.V. A revision of some Asian and African freshwater clams assigned to Corbicula fluminalis (Müller, 1774) (Mollusca: Bivalvia: Corbiculidae), with a review of anatomical characters and reproductive features based on museum collections. Hydrobiologia 2004, 529, 251–270. [Google Scholar] [CrossRef]
- Marescaux, J.; Pigneur, L.-M.; Van Doninck, K. New records of Corbicula clams in French rivers. Aquat. Invasions 2010, 5, 35–39. [Google Scholar] [CrossRef]
- Pigneur, L.M.; Marescaux, J.; Roland, K.; Etoundi, E.; Descy, J.P.; Van Doninck, K. Phylogeny and androgenesis in the invasive Corbicula clams (Bivalvia, Corbiculidae) in Western-Europe. BMC Evol. Biol. 2011, 11, 147. [Google Scholar] [CrossRef] [PubMed]
- Morhun, H.; Vinarski, M.V.; Labecka, A.M.; Velde, G.; van der Son, M.O. Differentiation of European invasive clams of the genus Corbicula (Cyrenidae) using shell shape analysis. J. Molluscan Stud. 2022, 88, eyab045. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Bolotov, I.N.; Makhrov, A.A.; Gofarov, M.Y.; Aksenova, O.V.; Aspholm, P.E.; Bespalaya, Y.V.; Kabakov, M.B.; Kolosova, Y.S.; Kondakov, A.V.; Ofenböck, T.; et al. Climate warming as a possible trigger of keystone mussel population decline in oligotrophic rivers at the continental scale. Sci. Rep. 2018, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotech. 1994, 3, 294–299. [Google Scholar]
- Mollaret, I.; Jamieson, B.G.; Adlard, R.D.; Hugall, A.; Lecointre, G.; Chombard, C.; Justine, J.L. Phylogenetic analysis of the Monogenea and their relationships with Digenea and Eucestoda inferred from 28S rDNA sequences. Mol. Biochem. Parasitol. 1997, 90, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Vastrade, M.; Etoundi, E.; Bournonville, T.; Colinet, M.; Debortoli, N.; Hedtke, S.M.; Nicolas, E.; Pigneur, L.-M.; Virgo, J.; Flot, J.-F.; et al. Substantial genetic mixing among sexual and androgenetic lineages within the clam genus Corbicula. Peer Community J. Evol. Biol. 2022, 2, e73. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.P.; Zardus, J.D.; Boyle, E.E.; Gonzalez, V.L.; Jennings, R.M.; McIntyre, E.; Wheeler, W.C.; Etter, R.J.; Giribet, G. Into the deep: A phylogenetic approach to the bivalve subclass Protobranchia. Mol. Phylogenet. Evol. 2013, 69, 188–204. [Google Scholar] [CrossRef] [PubMed]
- Siripattrawan, S.; Park, J.K.; Foighil, D.O. Two lineages of the introduced Asian freshwater clam Corbicula occur in North America. J. Molluscan Stud. 2000, 66, 423–429. [Google Scholar] [CrossRef]
- Haponski, A.E.; Ó Foighil, D. Phylogenomic analyses confirm a novel invasive North American Corbicula (Bivalvia: Cyrenidae) lineage. PeerJ 2019, 7, e7484. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | Species (Lineages, Forms) | Morphotype | Shell Shape | External Shell Coloration | Internal Shell Coloration | Shell Sculpture | Umbo Position | Anatomy of Siphons | COI Haplotype | Hybridizing Lineages ** |
|---|---|---|---|---|---|---|---|---|---|---|
| Volga River basin | Corbicula fluminea (=Corbicula clade FW5; =Corbicula R form) | R | Round and broad, triangular | Dark brown | Matte, with purple marks | Distinctly pronounced deep concentric ridges | Slightly shifted | Light yellow color with black spots in the siphonal papilla and dark pigmentation around | FW5 | R × Rlc |
| Don River basin | Corbicula fluminea (=Corbicula clade FW5) | Rlc | Oval triangular | Yellow or olive green | Yellow white, with small purple marks | Raised, regular concentric ribs | Centrally located | The papillae are long, have different lengths, and orange coloration with black spots and dark coloration around | FW5 | R × Rlc |
| C. fluminea (=R form) * | R | Oval triangular | Dark brown | White, with purple marks | Distinctly pronounced deep concentric ridges | Slightly shifted | N/A | N/A | N/A | |
| Shura-Ozen Sulak, and Samur River basin (Republic of Dagestan) | C. fluminalis (=Corbicula clade FW17; Corbicula form S) | S | Narrow, saddle-like | Yellow or olive green | Deep purple | Tight spaced ridges | Significantly prominent | The bases of each siphon and tentacle have a dark pigmentation | FW17 | R × S |
| Kura River (Stavropol Krai) | C. fluminalis (=Corbicula clade FW17; Corbicula form S) | “Intermediate form” Rlc + S | Oval triangular | Yellow or olive green | Deep purple | Raised, regular concentric ribs | Centrally located | Rather broad pigmented ring is not present. Siphonal papillae are pigmented | FW17 | Rlc × R × S |
| Species | 432 | 433 | 434 | 438 | 446 | 485 | 533 | 549 | 550 | 570 | 655 | 726 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C. fluminea (USA) | - | A | G | C | A | C | A | G | G | T | T | T | |
| C. fluminea (Arkhangelsk Oblast) ** (N = 5) | - | A/C | A/G | A/C | · | · | A/C | · | · | C/T | · | · | |
| C. fluminea (Kostroma Oblast) (N = 5) | - | · | · | · | · | · | · | · | · | · | · | · | |
| C. fluminea (Kostroma Oblast) ** (N = 5) | - | A/C | A/G | A/C | · | · | A/C | · | · | C/T | · | · | |
| C. fluminea (Rostov Oblast) (N = 5) | - | · | · | · | · | · | · | · | · | · | · | · | |
| C. fluminea (Rostov Oblast) ** (N = 5) | - | A/C | A/G | A/C | · | · | A/C | · | · | C/T | · | · | |
| C. fluminalis (Turkey) ** (N = 8) | - | C | A | A | · | · | C | A/G | G/T | C | G/T | · | |
| C. fluminalis (Dagestan) ** (N = 4) | - | · | · | · | · | A | · | · | · | · | · | · | |
| C. fluminalis (Dagestan)** (N = 1) | - | C | A | A | · | · | C | · | · | C | · | · | |
| C. fluminalis (Dagestan) *** (N = 5) | Var1 | - | C | A | A | · | · | C | · | · | C | · | · |
| Var2 | G | C | A | A | · | · | C | A | T | C | G | · | |
| C. fluminalis (Stavropol Krai) ** (N = 4) | - | · | · | · | · | A | · | · | · | · | · | · | |
| C. fluminalis (Stavropol Krai) *** (N = 5) | Var1 | - | C | A | A | · | · | C | · | · | C | · | · |
| Var2 | G | C | A | A | · | · | C | A | T | C | G | · | |
| C. fluminalis (Stavropol Krai) *** (N = 1) | Var1 | - | · | · | · | · | A | · | · | · | · | · | · |
| Var2 | G | C | A | A | · | · | C | A | T | C | G | · | |
| C. africana (South Africa) | G | C | A | A | G | · | C | · | · | C | G | · | |
| C. africana (South Africa) | G | C | A | A | · | · | C | · | · | C | · | C | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kropotin, A.V.; Bespalaya, Y.V.; Aksenova, O.V.; Kondakov, A.V.; Aksenov, A.S.; Khrebtova, I.S.; Palatov, D.M.; Travina, O.V.; Bolotov, I.N. Genetic and Morphological Characterization of the Invasive Corbicula Lineages in European Russia. Water 2023, 15, 3226. https://doi.org/10.3390/w15183226
Kropotin AV, Bespalaya YV, Aksenova OV, Kondakov AV, Aksenov AS, Khrebtova IS, Palatov DM, Travina OV, Bolotov IN. Genetic and Morphological Characterization of the Invasive Corbicula Lineages in European Russia. Water. 2023; 15(18):3226. https://doi.org/10.3390/w15183226
Chicago/Turabian StyleKropotin, Alexander V., Yulia V. Bespalaya, Olga V. Aksenova, Alexander V. Kondakov, Andrey S. Aksenov, Irina S. Khrebtova, Dmitry M. Palatov, Oksana V. Travina, and Ivan N. Bolotov. 2023. "Genetic and Morphological Characterization of the Invasive Corbicula Lineages in European Russia" Water 15, no. 18: 3226. https://doi.org/10.3390/w15183226