
Seasonal and Spatial Variations of Dissolved Organic Matter Biodegradation along the Aquatic Continuum in the Southern Taiga Bog Complex, Western Siberia

 ,
,  , ,
, ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
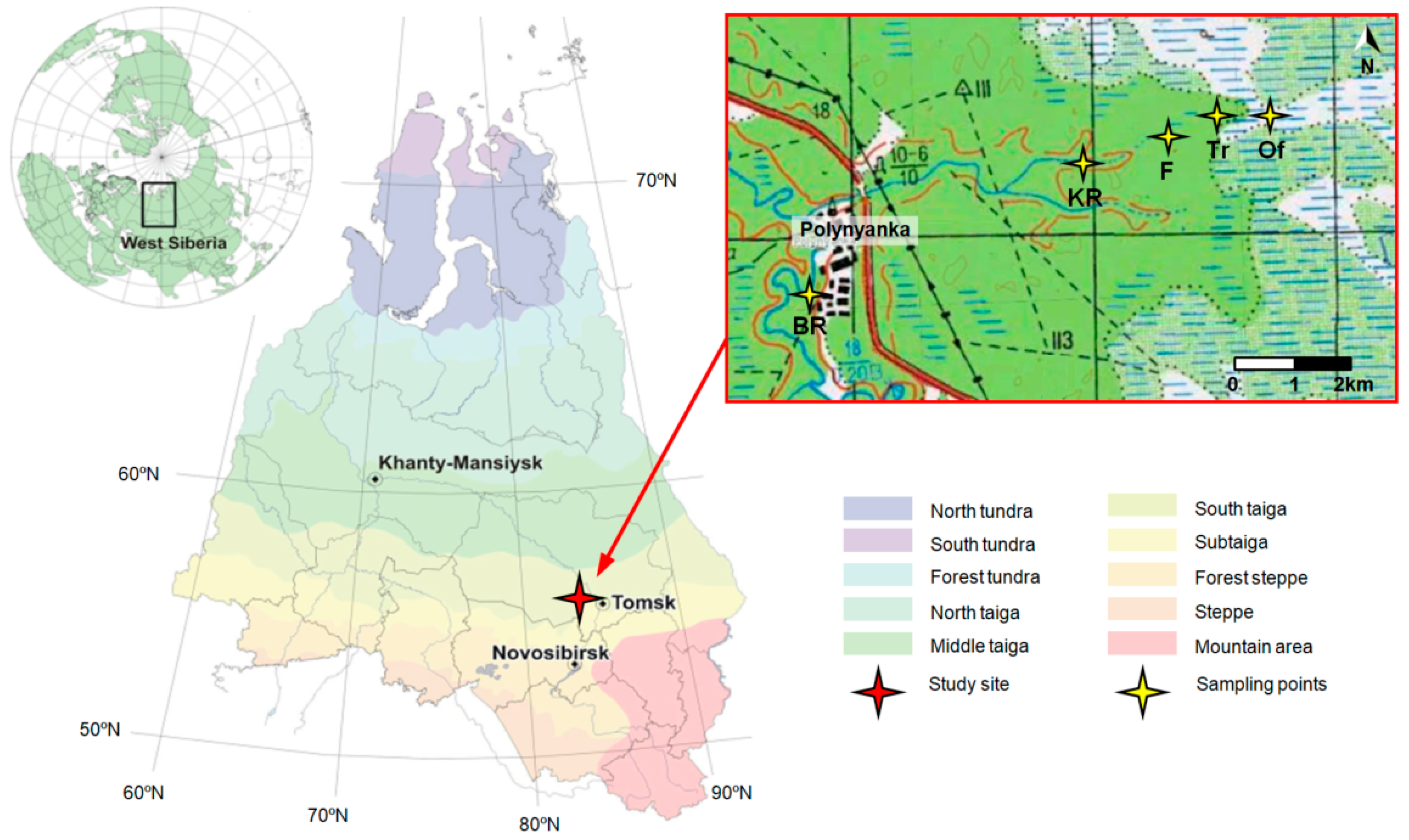
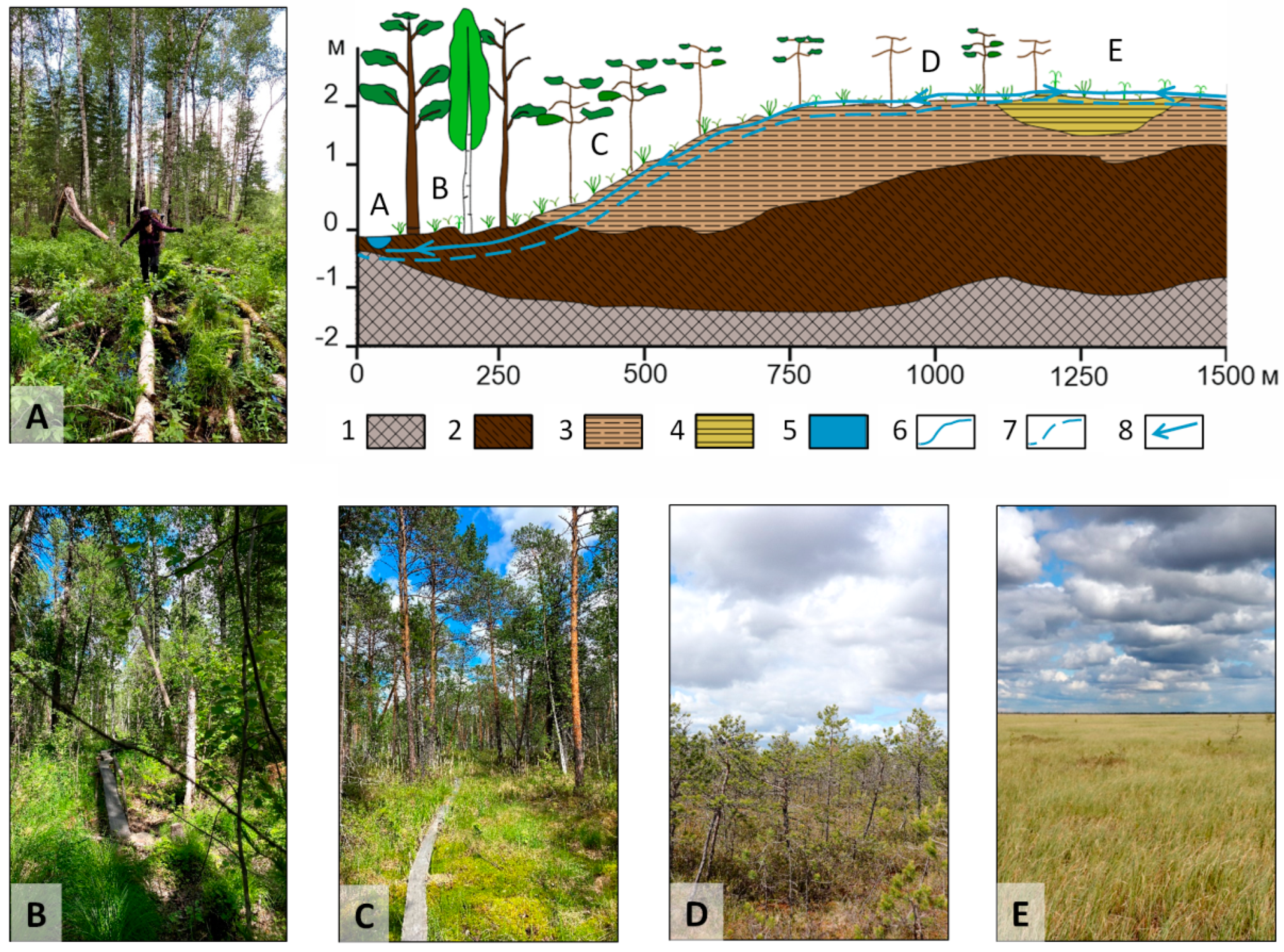
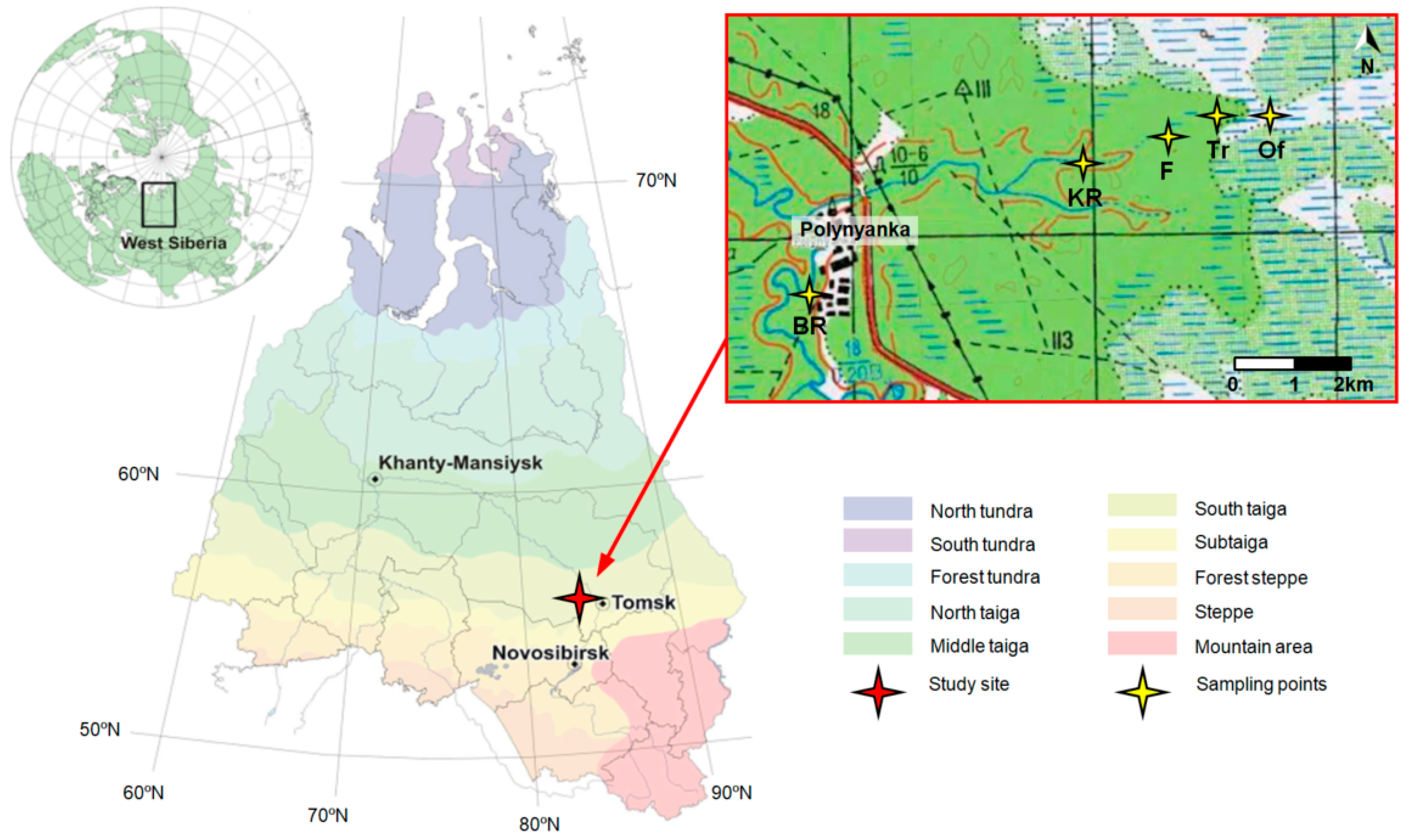
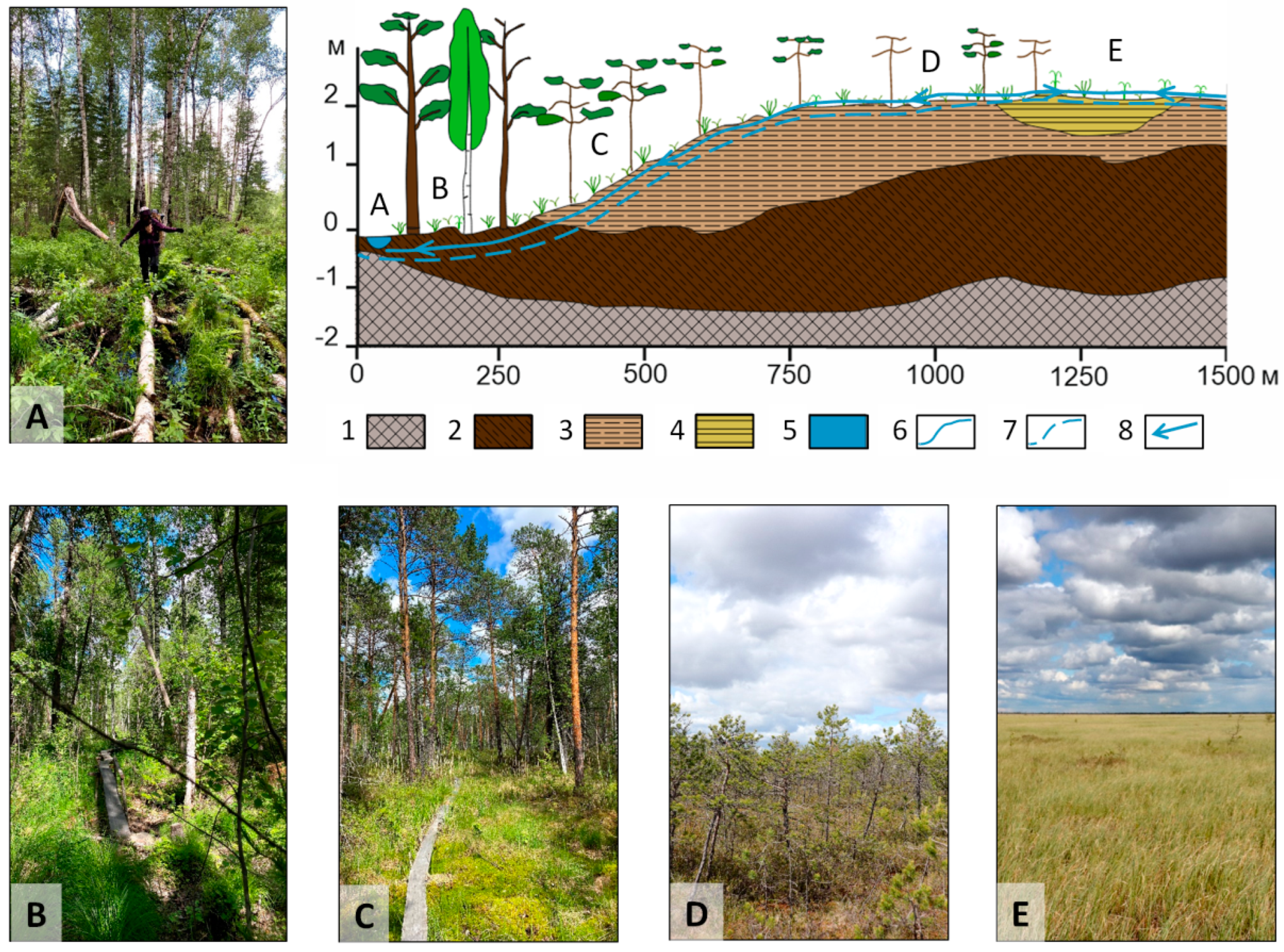
2.1. Study Area and Field Smpling
2.2. Experimental Setup and Data Treatment
3. Results and Discussion
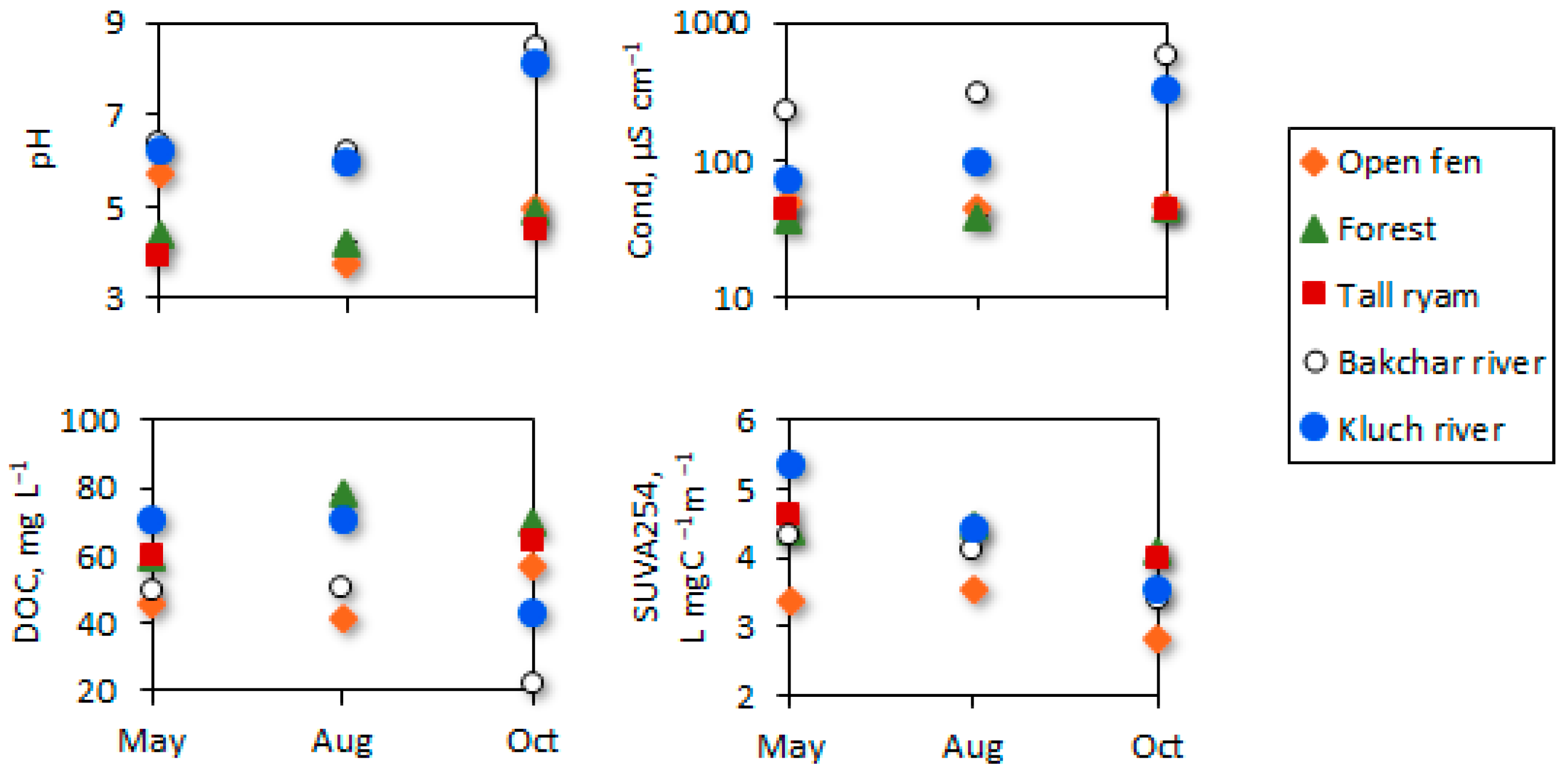
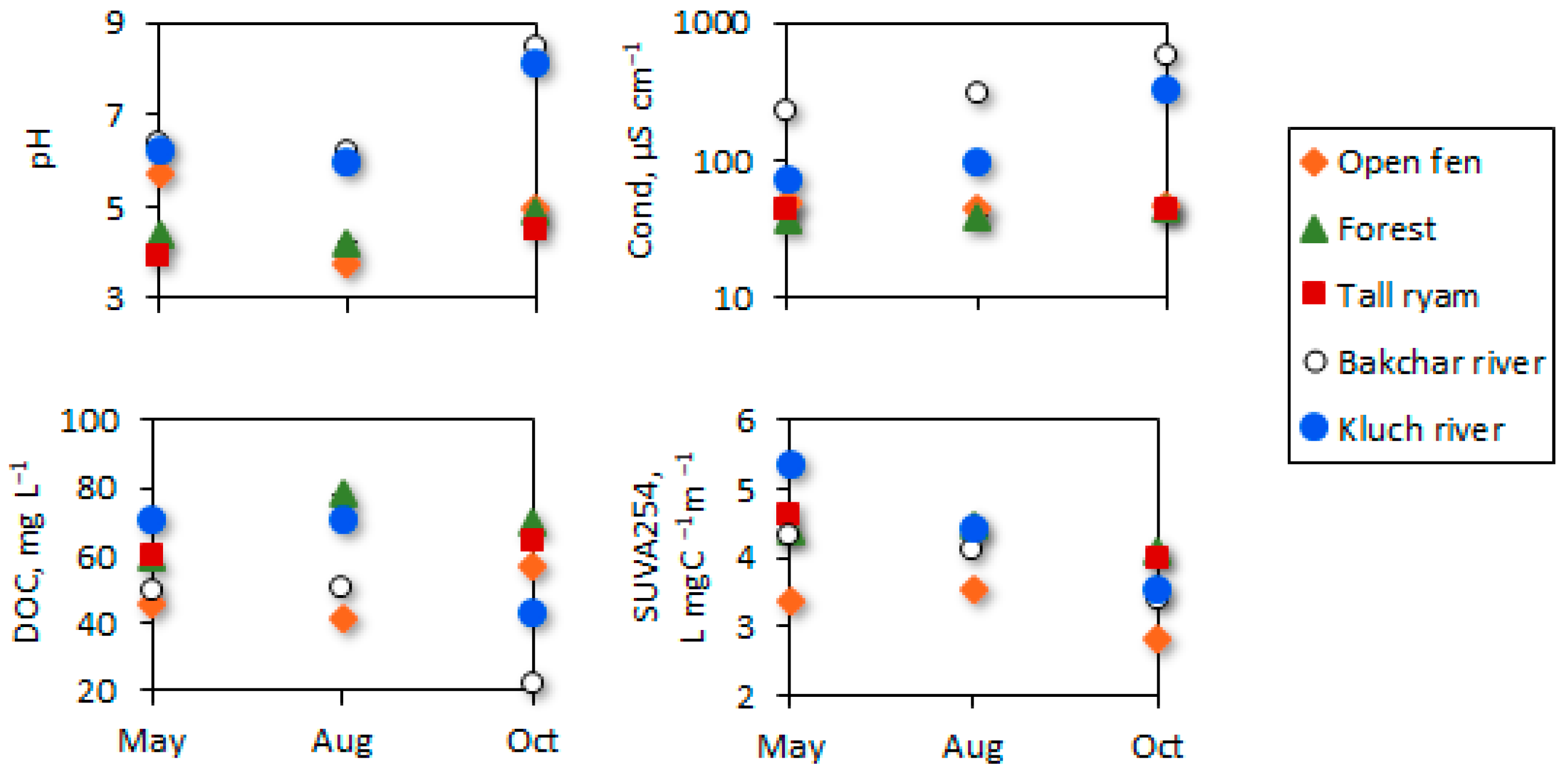
3.1. Initial Water Conditions across the Seasons
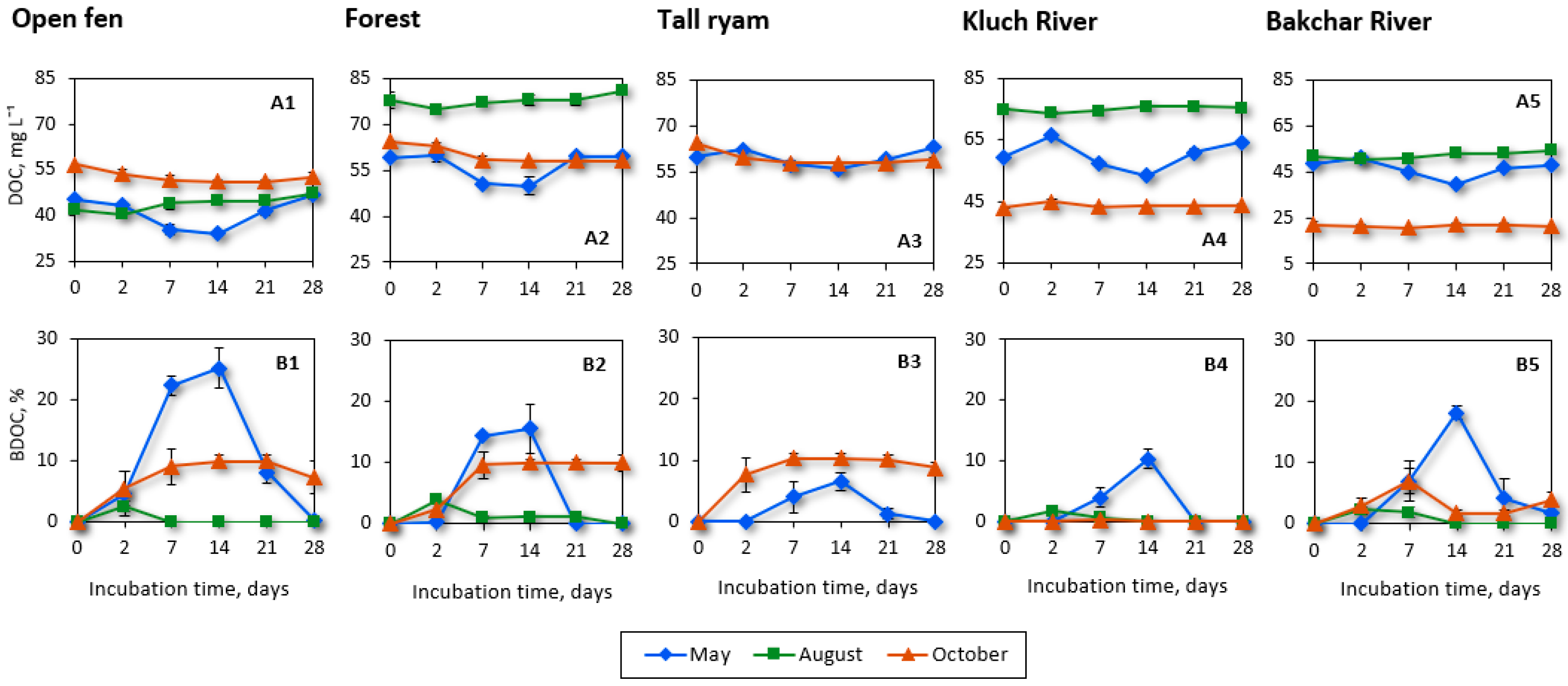
3.2. Variations in DOC Biodegradability
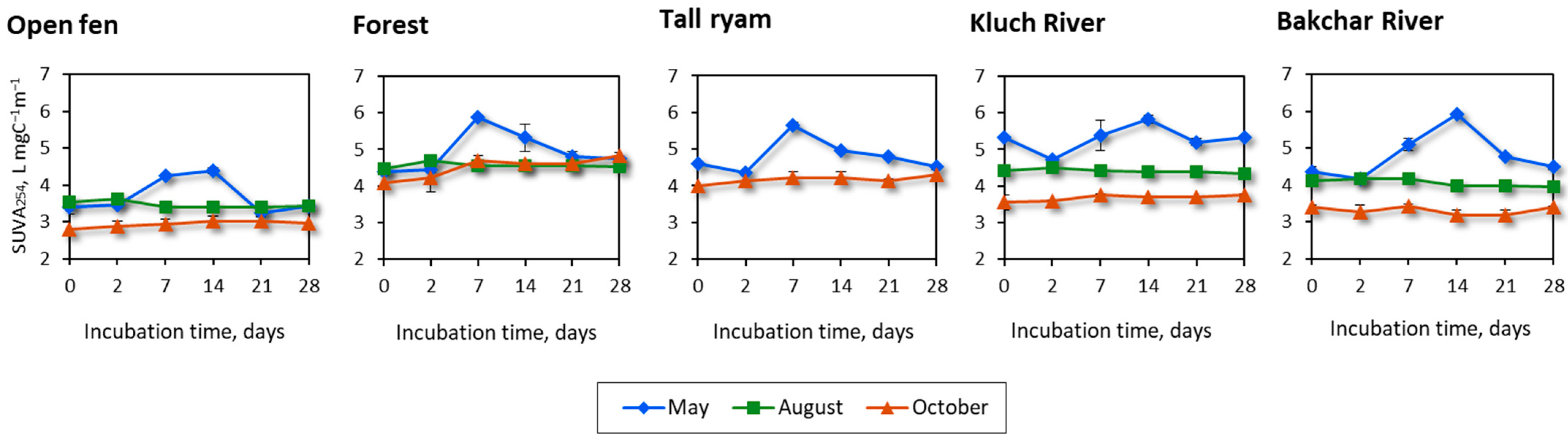
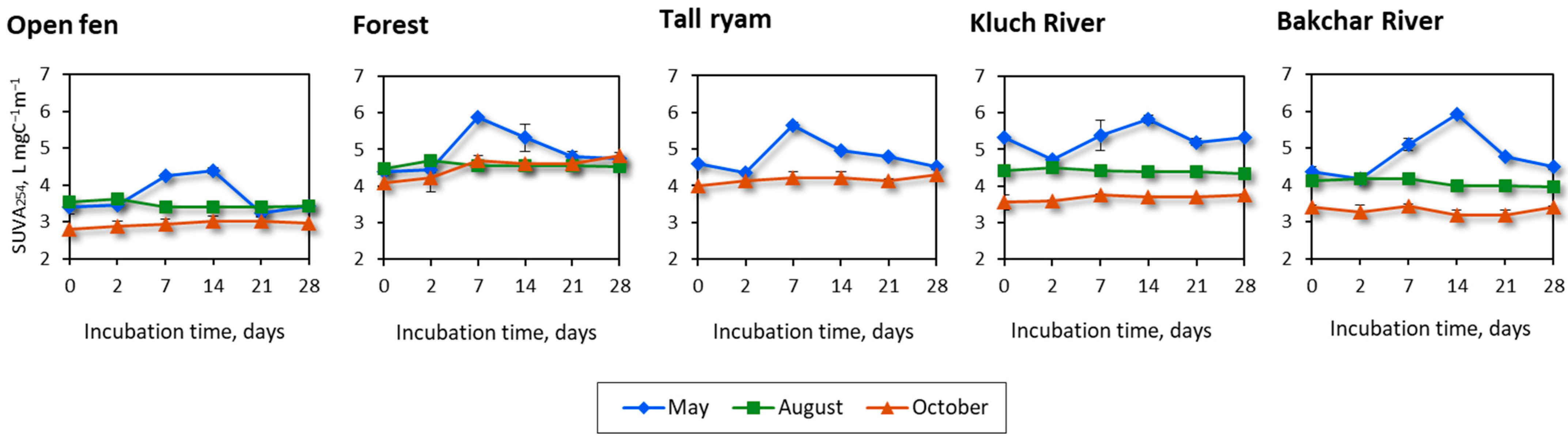
3.3. Optical Characteristics of CDOM during Incubation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tank, S.E.; Fellman, J.B.; Hood, E.; Kritzberg, E.S. Beyond respiration: Controls on lateral carbon fluxes across the terrestrial-aquatic interface. Limnol. Oceanogr. Lett. 2018, 3, 76–88. [Google Scholar] [CrossRef]
- Vonk, J.E.; Tank, S.E.; Mann, P.J.; Spencer, R.G.M.; Treat, C.C.; Striegl, R.G.; Abbott, B.W.; Wickland, K.P. Biodegradability of dissolved organic carbon in permafrost soils and aquatic systems: A meta-analysis. Biogeosciences 2015, 12, 6915–6930. [Google Scholar] [CrossRef] [Green Version]
- Cory, R.M.; Kling, G.W. Interactions between sunlight and microorganisms influence dissolved organic matter degradation along the aquatic continuum. Limnol. Oceanogr. Lett. 2018, 3, 102–116. [Google Scholar] [CrossRef]
- Holmes, R.M.; McClelland, J.W.; Raymond, P.A.; Frazer, B.B.; Peterson, B.J.; Stieglitz, M. Lability of DOC transported by Alaskan rivers to the Arctic Ocean. Geophys. Res. Lett. 2008, 35, L03402. [Google Scholar] [CrossRef] [Green Version]
- Marschner, B.; Kalbitz, K. Controls of bioavailability and biodegradability of dissolved organic matter in soils. Geoderma 2003, 113, 211–235. [Google Scholar] [CrossRef]
- Mann, P.J.; Davydova, A.; Zimov, N.; Spencer, R.G.M.; Davydov, S.; Bulygina, E.; Zimov, S.; Holmes, R.M. Controls on the composition and lability of dissolved organic matter in Siberia’s Kolyma River basin. J. Geophys. Res. Biogeosci. 2012, 117, G01028. [Google Scholar] [CrossRef] [Green Version]
- Wickland, K.P.; Aiken, G.R.; Butler, K.; Dornblaser, M.M.; Spencer, R.G.M.; Striegl, R.G. Biodegradability of dissolved organic carbon in the Yukon River and its tributaries: Seasonality and importance of inorganic nitrogen. Glob. Biogeochem. Cycles 2012, 26, GB0E03. [Google Scholar] [CrossRef]
- Abbott, B.W.; Larouche, J.R.; Jones, J.B.; Bowden, W.B.; Balser, A.W. Elevated dissolved organic carbon biodegradability from thawing and collapsing permafrost. J. Geophys. Res. 2014, 119, 2049–2063. [Google Scholar] [CrossRef]
- Vonk, J.E.; Tank, S.E.; Walvoord, M.A. Integrating hydrology and biogeochemistry across frozen landscapes. Nat. Commun. 2019, 10, 5377. [Google Scholar] [CrossRef] [Green Version]
- Schuur, E.A.G.; Bockheim, J.; Canadell, J.G.; Euskirchen, E.; Field, C.B.; Goryachkin, S.V.; Hagemann, S.; Kuhry, P.; Lafleur, P.M.; Lee, H.; et al. Vulnerability of permafrost carbon to climate change: Implications for the global carbon cycle. BioScience 2008, 58, 701–714. [Google Scholar] [CrossRef] [Green Version]
- Stutter, M.I.; Richards, S.; Dawson, J.J.C. Biodegradability of natural dissolved organic matter collected from a UK moorland stream. Water Res. 2013, 47, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Hulatt, C.J.; Kaartokallio, H.; Asmala, E.; Autio, R.; Stedmon, C.A.; Sonninen, E.; Oinonen, M.; Thomas, D.N. Bioavailability and radiocarbon age of fluvial dissolved organic matter (DOM) from a northern peatland-dominated catchment: Effect of land-use change. Aquat. Sci. 2014, 76, 393–404. [Google Scholar] [CrossRef]
- Oleinikova, O.; Drozdova, O.Y.; Lapitskiy, S.A.; Bychkov, A.Y.; Pokrovsky, O.S. Dissolved organic matter degradation by sunlight coagulates organo-mineral colloids and produces low molecular weight fraction of metals in boreal humic waters. Geochim. Cosmochim. Acta 2017, 211, 97–114. [Google Scholar] [CrossRef]
- Kaiser, K.; Canedo-Oropeza, M.; McMahon, R.; Amon, R.M.W. Origins and transformations of dissolved organic matter in large Arctic rivers. Sci. Rep. 2017, 7, 13064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirokova, L.S.; Chupakov, A.V.; Zabelina, S.A.; Neverova, N.V.; Payandi-Rolland, D.; Causserand, C.; Karlsson, J.; Pokrovsky, O.S. Humic surface waters of frozen peat bogs (permafrost zone) are highly resistant to bio- and photodegradation. Biogeosciences 2019, 16, 2511–2526. [Google Scholar] [CrossRef] [Green Version]
- Shakil, S.; Tank, S.E.; Vonk, J.E.; Zolkos, S. Low biodegradability of particulate organic carbon mobilized from thaw slumps on the Peel Plateau, NT, and possible chemosynthesis and sorption effects. Biogeosciences 2022, 19, 1871–1890. [Google Scholar] [CrossRef]
- Larouche, J.R.; Abbott, B.W.; Bowden, W.B.; Jones, J.B. The role of watershed characteristics, permafrost thaw, and wildfire on dissolved organic carbon biodegradability and water chemistry in Arctic headwater streams. Biogeosciences 2015, 12, 4221–4233. [Google Scholar] [CrossRef] [Green Version]
- Wickland, K.P.; Neff, J.C.; Aiken, G.R. Dissolved organic carbon in Alaskan boreal forest: Sources, chemical characteristics, and biodegradability. Ecosystems 2007, 8, 1323–1340. [Google Scholar] [CrossRef]
- Spencer, R.G.M.; Aiken, G.R.; Wickland, K.P.; Striegl, R.G.; Hernes, P.J. Seasonal and spatial variability in dissolved organic matter quantity and composition from the Yukon River basin, Alaska. Glob. Biogeochem. Cycles 2008, 22, GB4002. [Google Scholar] [CrossRef]
- Ward, C.P.; Nalven, S.G.; Crump, B.C.; Kling, G.W.; Cory, R.M. Photochemical alteration of organic carbon draining permafrost soils shifts microbial metabolic pathways and stimulates respiration. Nat. Commun. 2017, 8, 772. [Google Scholar] [CrossRef]
- Kawahigashi, M.; Kaiser, K.; Kalbitz, K.; Rodionov, A.; Guggenberger, G. Dissolved organic matter in small streams along a gradient from discontinuous to continuous permafrost. Glob. Chang. Biol. 2004, 10, 1576–1586. [Google Scholar] [CrossRef]
- Mann, P.J.; Eglinton, T.I.; McIntyre, C.P.; Zimov, N.; Davydova, A.; Vonk, J.E.; Holmes, R.M.; Spencer, R.G.M. Utilization of ancient permafrost carbon in headwaters of Arctic fluvial networks. Nat. Commun. 2015, 6, 7856. [Google Scholar] [CrossRef] [Green Version]
- Frey, K.E.; Sobczak, W.V.; Mann, P.J.; Holmes, R.M. Optical properties and bioavailability of dissolved organic matter along a flow-path continuum from soil pore waters to the Kolyma River mainstem, East Siberia. Biogeosciences 2016, 13, 2279–2290. [Google Scholar] [CrossRef] [Green Version]
- Keskitalo, K.H.; Bröder, L.; Jong, D.; Zimov, N.; Davydova, A.; Davydov, S.; Tesi, T.; Mann, P.J.; Haghipour, N.; Eglinton, T.I.; et al. Seasonal variability in particulate organic carbon degradation in the Kolyma River, Siberia. Environ. Res. Lett. 2022, 17, 034007. [Google Scholar] [CrossRef]
- Köhler, S.; Buffam, I.; Jonsson, A.; Bishop, K. Photochemical and microbial processing of stream and soil water dissolved organic matter in a boreal forested catchment in northern Sweden. Aquat. Sci. 2002, 64, 269–281. [Google Scholar] [CrossRef]
- Berggren, M.; Laudon, H.; Jansson, M. Hydrological control of organic carbon support for bacterial growth in boreal headwater streams. Microb. Ecol. 2009, 57, 170–178. [Google Scholar] [CrossRef]
- Attermeyer, K.; Catalán, N.; Einarsdottir, K.; Freixa, A.; Groeneveld, M.; Hawkes, J.A.; Bergquist, J.; Tranvik, L.J. Organic carbon processing during transport through boreal inland waters: Particles as important sites. J. Geophys. Res. Biogeosci. 2018, 123, 2412–2428. [Google Scholar] [CrossRef] [Green Version]
- Einarsdóttir, K.; Attermeyer, K.; Hawkes, J.A.; Kothawala, D.; Sponseller, R.A.; Tranvik, L.J. Particles and aeration at mire-stream interfaces cause selectiveremoval and modification of dissolved organic matter. J. Geophys. Res. Biogeosci. 2020, 125, e2020JG005654. [Google Scholar] [CrossRef]
- Roehm, C.L.; Giesler, R.; Karlsson, J. Bioavailability of terrestrial organic carbon to lake bacteria: The case of a degrading subarctic permafrost mire complex. J. Geophys. Res. Biogeosci. 2009, 114, G03006. [Google Scholar] [CrossRef]
- Payandi-Rolland, D.; Shirokova, L.S.; Tesfa, M.; Lim, A.G.; Kuzmina, D.; Benezeth, P.; Karlsson, J.; Giesler, R.; Pokrovsky, O.S. Dissolved organic matter biodegradation along a hydrological continuum in a discontinuous permafrost area: Case study of northern Siberia and Sweden. Sci. Total Environ. 2020, 749, 141463. [Google Scholar] [CrossRef]
- Winter, A.R.; Fish, T.A.E.; Playle, R.C.; Smith, D.S.; Curtis, P.J. Photodegradation of natural organic matter from diverse freshwater sources. Aquat. Toxicol. 2007, 84, 215–222. [Google Scholar] [CrossRef]
- Cory, R.M.; Ward, C.P.; Crump, B.C.; Kling, G.W. Sunlight controls water column processing of carbon in arctic fresh waters. Science 2014, 345, 925–928. [Google Scholar] [CrossRef] [PubMed]
- Porcal, P.; Dillon, P.J.; Molot, L.A. Photochemical production and decomposition of particulate organic carbon in a freshwater stream. Aquat. Sci. 2013, 75, 469–482. [Google Scholar] [CrossRef]
- Selvam, B.P.; Lapierre, J.-F.; Guillemette, F.; Voigt, C.; Lamprecht, R.E.; Biasi, C.; Christensen, T.R.; Martikainen, P.J.; Berggren, M. Degradation potentials of dissolved organic carbon (DOC) from thawed permafrost peat. Sci. Rep. 2017, 7, 45811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gareis, J.A.L.; Lesack, L.F.W. Photodegraded dissolved organic matter from peak freshet river discharge as a substrate for bacterial production in a lake-rich great Arctic delta. Arct. Sci. 2018, 4, 557–583. [Google Scholar] [CrossRef] [Green Version]
- Oleinikova, O.; Shirokova, L.S.; Drozdova, O.Y.; Lapitsky, S.A.; Pokrovsky, O.S. Low biodegradability of dissolved organic matter and trace metal from subarctic waters by culturable heterotrophic bacteria. Sci. Total Environ. 2018, 618, 174–187. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, J.; Serikova, S.; Vorobyev, S.N.; Rocher-Ros, G.; Denfeld, B.; Pokrovsky, O.S. Carbon emission from Western Siberian inland waters. Nat. Commun. 2021, 12, 825. [Google Scholar] [CrossRef]
- Bohn, T.J.; Lettenmaier, D.P.; Sathulur, K.; Bowling, L.C.; Podest, E.; McDonald, K.C.; Friborg, T. Methane emissions from western Siberian wetlands: Heterogeneity and sensitivity to climate change. Environ. Res. Lett. 2007, 2, 045015. [Google Scholar] [CrossRef]
- Pokrovsky, O.S.; Manasypov, R.M.; Loiko, S.V.; Krickov, I.A.; Kopysov, S.G.; Kolesnichenko, L.G.; Vorobyev, S.N.; Kirpotin, S.N. Trace element transport in western Siberian rivers across a permafrost gradient. Biogeosciences 2016, 13, 1877–1900. [Google Scholar] [CrossRef] [Green Version]
- Dyukarev, E.A.; Martynova, Y.V.; Golovatskaya, E.A. Assessment of the carbon balance of treed bogs under climate change with observation and modelling data. IOP Conf. Ser. Earth Environ. Sci. 2019, 386, 012028. [Google Scholar] [CrossRef]
- Kirpotin, S.N.; Berezin, A.; Bazanov, V.; Polishchuk, Y.; Vorobiov, S.; Mironycheva-Tokoreva, N.; Kosykh, N.; Volkova, I.; Dupre, B.; Pokrovsky, O.; et al. Western Siberia wetlands as indicator and regulator of climate change on the global scale. Int. J. Environ. Stud. 2009, 66, 409–421. [Google Scholar] [CrossRef]
- Inisheva, L.I.; Inishev, N.G. Elements of water balance and hydrochemical characteristic of oligotrophic bogs in the Southern Taiga subzone of Western Siberia. Water Resour. 2001, 28, 371. [Google Scholar] [CrossRef]
- Shvartsev, S.L.; Serebrennikova, O.V.; Zdvizhkov, M.A.; Savichev, O.G.; Naimushina, O.S. Geochemistry of wetland waters from the lower Tom Basin, southern Tomsk oblast. Geochem. Int. 2012, 50, 367–380. [Google Scholar] [CrossRef]
- Naymushina, O.; Shvartsev, S.; Serebrennikova, O.; Ses, K.; Matveenko, I. Spatial distribution of chemical and organic compounds in thewater of oligotrophic peatland of Tomsk region (Western Siberia). Procedia Chem. 2014, 10, 541–546. [Google Scholar] [CrossRef] [Green Version]
- Savichev, O.G. Geochemical parameters of bog waters in the taiga zone of the Western Siberia. Izv. Ross. Akad. Nauk. Seriya Geogr. 2015, 4, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Savichev, O.G.; Mazurov, A.K.; Semiletov, I.P.; Bazanov, V.A.; Guseva, N.V.; Khvashchevskaya, A.A.; Nalivajko, N.G. Hydrogeochemical conditions of formation oligotrophic bog ecosystems. Izv. Ross. Akad. Nauk. Seriya Geogr. 2016, 5, 60–69. [Google Scholar] [CrossRef]
- Kharanzhevskaya, Y.A.; Voistinova, E.S.; Sinyutkina, A.A. Spatial and temporal variations in mire surface water chemistry as a function of geology, atmospheric circulation and zonal features in the south-eastern part of Western Siberia. Sci. Total Environ. 2020, 733, 139343. [Google Scholar] [CrossRef]
- Savichev, O.G.; Skugarev, A.A.; Bazanov, V.A.; Kharanzhevskaya, Y.A. Water balance of the wetland catchment area in West Siberia (on the example of the small river Key, Tomsk region). Geoinformatika 2011, 3, 39–46. [Google Scholar]
- Seo, E.Y.; Ahn, T.S.; Zo, Y.G. Agreement, precision, and accuracy of epifluorescence microscopy methods for enumeration of total bacterial numbers. Appl. Environ. Microbiol. 2010, 76, 1981–1991. [Google Scholar] [CrossRef] [Green Version]
- Helms, J.R.; Stubbins, A.; Ritchie, J.D.; Minor, E.C.; Kieber, D.J.; Mopper, K. Absorption spectral slopes and slope ratios as indicators of molecular weight, source, and photobleaching of chromophoric dissolved organic matter. Limnol. Oceanogr. 2008, 53, 955–969. [Google Scholar] [CrossRef] [Green Version]
- Chin, Y.-P.; Aiken, G.; O’Loughlin, E. Molecular weight, polydispersity, and spectroscopic properties of aquatic humic substances. Environ. Sci. Technol. 1994, 28, 1853–1858. [Google Scholar] [CrossRef]
- Weishaar, J.L.; Aiken, G.R.; Bergamaschi, B.A.; Fram, M.S.; Fujii, R.; Mopper, K. Evaluation of specific ultraviolet absorbance as an indicator of the chemical composition and reactivity of dissolved organic carbon. Environ. Sci. Technol. 2003, 37, 4702–4708. [Google Scholar] [CrossRef] [PubMed]
- Peacock, M.; Evans, C.D.; Fenner, N.; Freeman, C.; Gough, R.; Jones, T.G.; Lebron, I. UV-visible absorbance spectroscopy as a proxy for peatland dissolved organic carbon (DOC) quantity and quality: Considerations on wavelength and absorbance degradation. Environ. Sci. Process. Impacts 2014, 16, 1445–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uyguner, C.; Bekbolet, M. Implementation of spectroscopic parameters for practical monitoring of natural organic matter. Desalination 2005, 176, 47–55. [Google Scholar] [CrossRef]
- O’Donnell, J.A.; Aiken, G.R.; Kane, E.S.; Jones, J.B. Source water controls on the character and origin of dissolved organic matter in streams of the Yukon River basin, Alaska. J. Geophys. Res. 2010, 115, G03025. [Google Scholar] [CrossRef] [Green Version]
- Spencer, R.G.M.; Stubbins, A.; Hernes, P.J.; Baker, A.; Mopper, K.; Aufdenkampe, A.K.; Dyda, R.Y.; Mwamba, V.L.; Mangangu, A.M.; Wabakanghanzi, J.N.; et al. Photochemical degradation of dissolved organic matter and dissolved lignin phenols from the Congo River. J. Geophys. Res. 2009, 114, G03010. [Google Scholar] [CrossRef] [Green Version]
- Kalbitz, K.; Schwesig, D.; Schmerwitz, J.; Kaiser, K.; Haumaier, L.; Glaser, B.; Ellerbrock, R.; Leinweber, P. Changes in properties of soil derived dissolved organic matter induced by biodegradation. Soil Biol. Biochem. 2003, 35, 1129–1142. [Google Scholar] [CrossRef]
- Krickov, I.V.; Lim, A.G.; Shirokova, L.S.; Korets, M.A.; Karlsson, J.; Pokrovsky, O.S. Environmental Carbon Emission and Concentration Patterns in Siberian Taiga Rivers across Seasons. Available online: https://ssrn.com/abstract=4221864 (accessed on 1 December 2022).
- Lapierre, J.-F.; Guillemette, F.; Berggren, M.; del Giorgio, P.A. Increases in terrestrially derived carbon stimulate organic carbon processing and CO2 emissions in boreal aquatic ecosystems. Nat. Commun. 2013, 4, 2972. [Google Scholar] [CrossRef] [Green Version]
- Moody, C.S.; Worrall, F. Modeling rates of DOC degradation using DOM composition and hydroclimatic variables. J. Geophys. Res. Biogeosci. 2017, 122, 1175–1191. [Google Scholar] [CrossRef] [Green Version]
- Hutchins, R.H.S.; Aukes, P.; Schiff, S.L.; Dittmar, T.; Prairie, Y.T.; del Giorgio, P.A. The optical, chemical, and molecular dissolved organic matter succession along a boreal soil-stream-river continuum. J. Geophys. Res. Biogeosci. 2017, 122, 2892–2908. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raudina, T.V.; Smirnov, S.V.; Lushchaeva, I.V.; Istigechev, G.I.; Kulizhskiy, S.P.; Golovatskaya, E.A.; Shirokova, L.S.; Pokrovsky, O.S. Seasonal and Spatial Variations of Dissolved Organic Matter Biodegradation along the Aquatic Continuum in the Southern Taiga Bog Complex, Western Siberia. Water 2022, 14, 3969. https://doi.org/10.3390/w14233969
Raudina TV, Smirnov SV, Lushchaeva IV, Istigechev GI, Kulizhskiy SP, Golovatskaya EA, Shirokova LS, Pokrovsky OS. Seasonal and Spatial Variations of Dissolved Organic Matter Biodegradation along the Aquatic Continuum in the Southern Taiga Bog Complex, Western Siberia. Water. 2022; 14(23):3969. https://doi.org/10.3390/w14233969
Chicago/Turabian StyleRaudina, Tatiana V., Sergei V. Smirnov, Inna V. Lushchaeva, Georgyi I. Istigechev, Sergey P. Kulizhskiy, Evgeniya A. Golovatskaya, Liudmila S. Shirokova, and Oleg S. Pokrovsky. 2022. "Seasonal and Spatial Variations of Dissolved Organic Matter Biodegradation along the Aquatic Continuum in the Southern Taiga Bog Complex, Western Siberia" Water 14, no. 23: 3969. https://doi.org/10.3390/w14233969
APA StyleRaudina, T. V., Smirnov, S. V., Lushchaeva, I. V., Istigechev, G. I., Kulizhskiy, S. P., Golovatskaya, E. A., Shirokova, L. S., & Pokrovsky, O. S. (2022). Seasonal and Spatial Variations of Dissolved Organic Matter Biodegradation along the Aquatic Continuum in the Southern Taiga Bog Complex, Western Siberia. Water, 14(23), 3969. https://doi.org/10.3390/w14233969