
Zooplankton Community Structure in Shallow Saline Steppe Inland Waters


Abstract
1. Introduction
- -
- to assess the less known zooplankton assemblages of representative shallow inland saline waters of North Kazakhstan on a large spatial scale (1000 km) in the arid steppe region;
- -
- to survey and identify dominant and indicator species among rotifer and microcrustacean taxa, and contribute to the available literature;
- -
- to analyze the change in abundance and taxonomic diversity along salinity gradient;
- -
- to determine the salt concentration ranges where significant quantitative and qualitative changes occur in the zooplankton community structure of shallow waters.
2. Materials and Methods
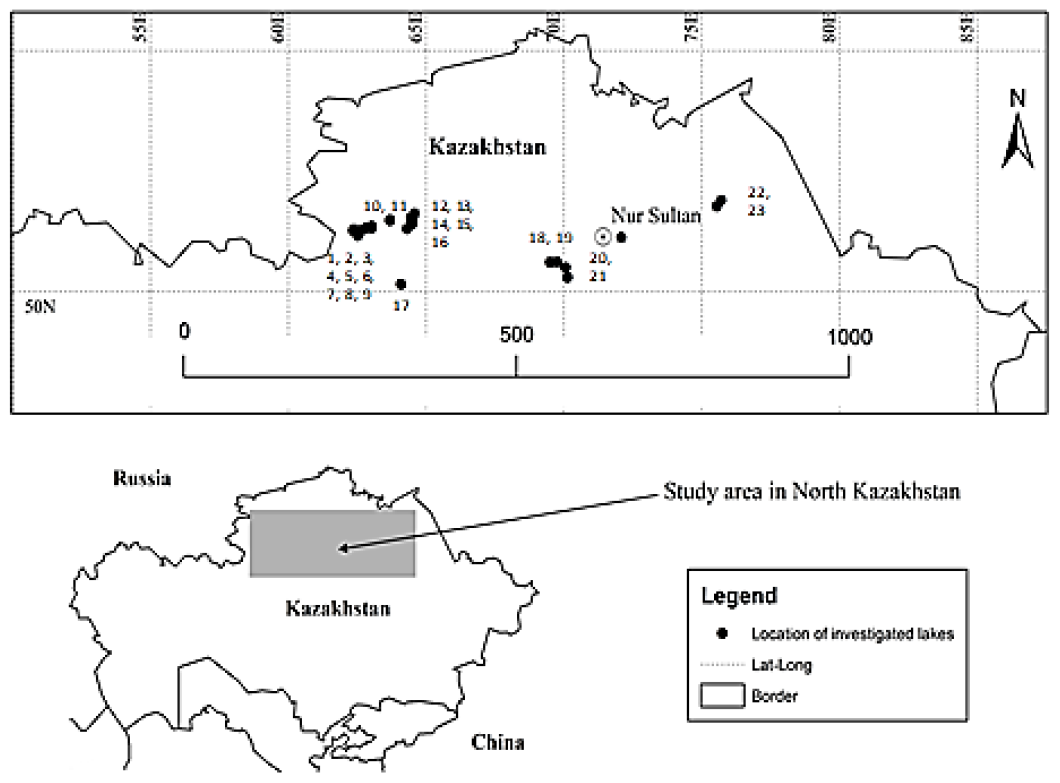
2.1. Study Area
2.2. Water Sampling and Zooplankton Identification
2.3. Data Analysis
3. Results and Discussion
3.1. Relationship among the Zooplankton Community, Physical, Chemical, and Trophic Properties
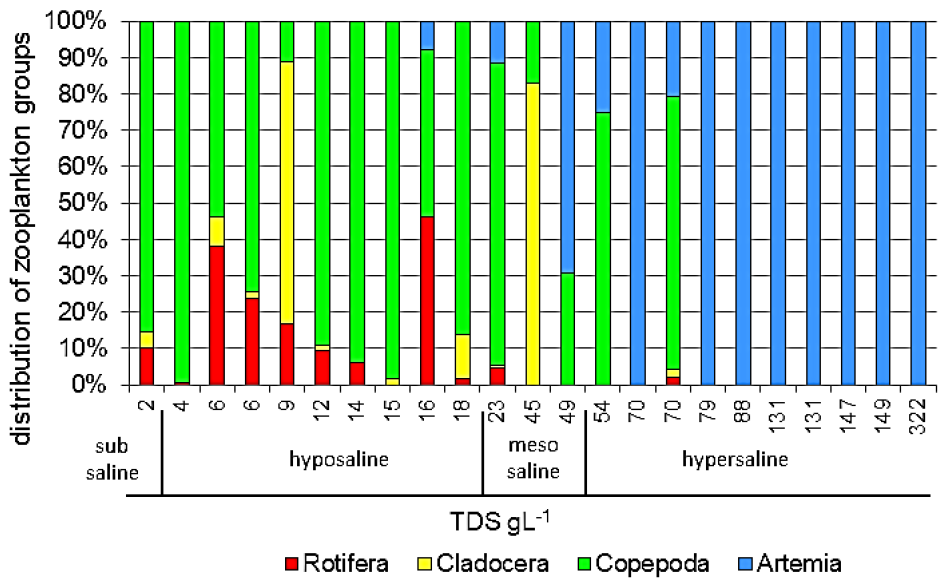
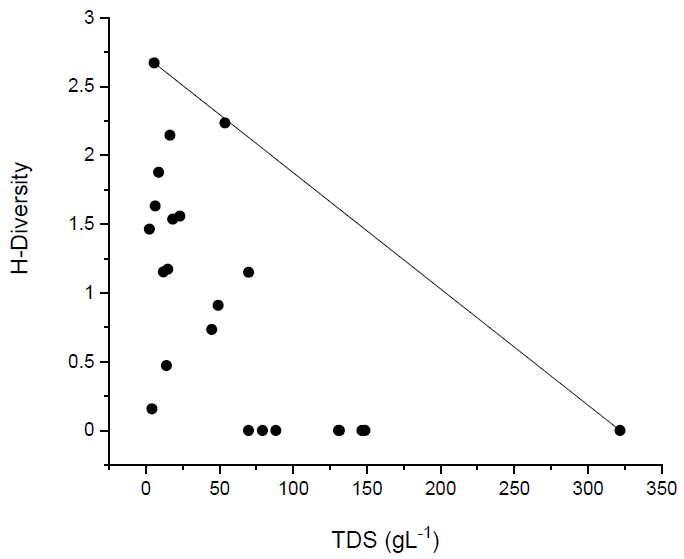
3.2. The Influence of Salinity on Zooplankton Abundance and Structure
3.3. Salt Tolerance of Registered Zooplankton Species
3.3.1. Rotifera Assemblage
3.3.2. Cladocera Assemblage
3.3.3. Copepoda Assemblage
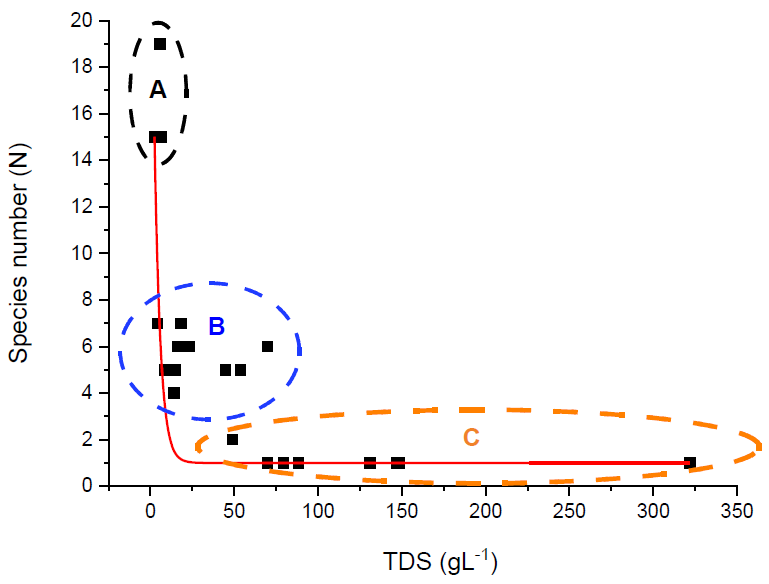
3.3.4. Grouping of Waters according to Zooplankton Species Composition
- In the first group, one can find the low-salt sub-and hyposaline waters. The zooplankton community consists of euryhaline copepod and rotifer species with a wide ecological valence.
- -
- There are: Teniz, Sukyrkol, Shoshkakol, Little Aqsuat, Zharkol.
- In the second group, there are those meso- and hyposaline waters, in which microcrustaceans are the main constituents, sometimes with large numbers of individuals. Brachionus asplanchnoides, Brachionus dimidiatus, Brachionus plicatilis, Hexarthra fennica, Moina salina, Arctodiaptomus salinus, and Cletocamptus rectirostris are present as saltwater indicator species.
- -
- There are: No.8. Unnamed, No.10. Unnanmed, No.11. Unnamed, No.14. Unnamed, Zharman Koli, Saryqopa Koli, Balyksor, Big Saryoba, Karasor.
- In the third group, there are the shallow, hypersaline water bodies, with few species, the characteristic organism is Artemia, which is present as a constituent of a dominant or monodominant association.
- -
- There are: Kaiyndsor, Abubastysor, Ukrash, Zharsor, No.7. Unnamed, No.9. Unnamed, Little Tengiz, Kalmakty, Boshchesor.
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williams, W.D. Environmental threats to salt lakes and the likely status of inland saline ecosystems in 2025. Environ. Conserv. 2002, 29, 154–167. [Google Scholar] [CrossRef]
- Ginatullina, E.; Atwell, L.; Saito, L. Resilience and resistance of zooplankton communities to drought-induced salinity in freshwater and saline lakes of Central Asia. J. Arid Environ. 2017. [Google Scholar] [CrossRef]
- Shadrin, N.V.; Anufriieva, E.V. Structure and Trophic Relations in Hypersaline Environments. Biol. Bull. Rev. 2020, 10, 48–56. [Google Scholar] [CrossRef]
- Aladin, N.V.; Plotnikov, I.S. Large saline lakes of former USSR: A summary review. Hydrobiologia 1993, 267, 1–12. [Google Scholar] [CrossRef]
- Saduakosova, R.E. Zooplankton of Lake Balkhash. Fish Resour. Kazakhstan Water Bodies 1972, 7, 97–100. (In Russian) [Google Scholar]
- Loginovskikh, E.V.; Dyusengaliev, T. Quantitative characteristics of the zooplankton of the Alakol Lake. Fish Resour. Kazakhstan Water Bodies 1972, 7, 89–94. (In Russian) [Google Scholar]
- Burlibaeva, M.Z.; Kurocskinoi, L.J.; Kaseeva, B.A.; Erokova, S.; Ivasenko, N. Globally significant wetlands of Kazakhstan. Tengiz-Korgalzhyn Lake System. Astana. Kazakhstan. 2007, 2, 286. (In Russian) [Google Scholar]
- Yermolaeva, N.I. Some results of Studying Zooplankton in Lakes of Northern Kazahkstan. Arid Ecosyst. 2013, 3, 263–275. [Google Scholar] [CrossRef]
- Aubakirova, G.A.; Syzdykov, K.N.; Kurzhykayev, Z.; Uskenov, R.B.; Narbayev, S.; Begenova, A.B.; Zhumakayeva, A.N.; Sabdinova, D.K.; Akhmedinov, A.N. Quantitative Development and Distribution of Zooplankton in Medium Lakes of the Kostanay Region (North Kazakhstan Region). Int. J. Environ. Sci. Educ. 2016, 11, 8193–8210. [Google Scholar]
- Boros, E.; Vörös, L.; Burlibajeva, D.; Aldiyarova, A.; Narbayeva, K. Salinity and trophic status of the shallow standing water bodies in the Central Asian steppe (North Kazakhstan). In Water Supply and Water Management in Irrigated Agriculture and Irrigation of Pastures. 2015. Conference Book; Kaz NAU: Almaty, Kazakhstan, 2015; pp. 164–172. [Google Scholar]
- Boros, E.; Jurecska, L.; Tatár, E.; Vörös, L.; Kolpakova, M. Chemical composition and trophic state of shallow saline steppe lakes in central Asia (North Kazakhstan). Environ. Monit. Assess. 2017, 189, 546. [Google Scholar] [CrossRef]
- Issanova, G.T.; Abuduwailii, J.; Mamutov, Z.U.A.; Kaldybaev, A.; Saparov, G.A.; Bazarbaeva, T.A. Saline Soils and Identification of Salt Accumulation Provinces in Kazakhstan. Arid Ecosyst. 2017, 7, 243–250. [Google Scholar] [CrossRef]
- Available online: https://central-asia.guide/kazakhstan/destinations-kz/naurzum-nature-reserve/20210410 (accessed on 15 April 2021).
- Van Stappen, G. Introduction, biology and ecology of Artemia. In Manual on the Production and Use of Live Food for Aquaculture; Lavens, P., Sorgeles, P., Eds.; FAO: Roma, Italy, 1996. [Google Scholar]
- Hammer, U.T. Saline Lake Ecosystems of the World. Monographiae Biologica; Dumont, H.J., Dr, W., Eds.; Springer: Berlin, Germany, 1986; Volume 59. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University Illinois Press: Urbana, IL, USA, 1964. [Google Scholar]
- Available online: http://www.originlab.com (accessed on 15 April 2021).
- Anufriieva, E. Copepods in Hypersaline Waters Worldwide: Diversity, Environmental, Social, and Economic Roles. Acta Geol. Sin. 2014, 88 (Suppl. 1), 43–45. [Google Scholar] [CrossRef]
- Derry, A.M.; Prepas, E.E.; Herbert, P.N.D. A comparison of zooplankton communities in saline lakewater with variable anion composition. Hydrobiologia 2003, 505, 199–215. [Google Scholar] [CrossRef]
- Belmonte, G.; Moscatello, S.; Batogova, E.A.; Pavlovskaya, T.; Shadrin, N.V.; Litvinchuk, L.F. Fauna of hypersaline lakes of the Crimea (Ukraine). Thalass. Salentina 2012, 34, 11–24. [Google Scholar]
- Wang, H.; Yuan, X.; Zheng, M. Ecological Distribution and Significance of Metadiaptomus asiaticus Uljanirs, a Copepod Recently Found on the Qinghai-Tibetan Platau. Acta Geol. Sin. 2014, 88.1, 108–110. [Google Scholar]
- Shadrin, N.V.; Anufriieva, E.V. Dependence of Arctodiaptomus salinus (Calanoida, Copepoda) halotolerance on exoosmolytes: New data and a hypothesis. J. Mediterr. Ecol. 2013, 12, 21–26. [Google Scholar]
- Afonina, E.Y.; Tashlykova, N.A. Plankton of Saline Lakes: Transformation and Environmental Factors. Contemp. Probl. Ecol. 2019, 12, 155–170. [Google Scholar] [CrossRef]
- Fontaneto, D.; De Smet, W.H.; Ricci, C. Review Rotifers in Saltwater Environments, Re-evolution of an Inconspicuous Taxon. J. Mar. Assoc. UK 2006, 86, 623–656. [Google Scholar] [CrossRef]
- Tóth, A.; Horváth, Z.; Vad, C.F.; Zsuga, K.; Nagy, S.A.; Boros, E. Zooplankton of the European soda ponds: Fauna and conservation of a unique habitat type. Int. Rev. Hydrobiol. 2014, 99, 255–276. [Google Scholar] [CrossRef]
- Anufriieva, E.V. Do copepods inhabit hypersaline waters worldwide? A short review discussion. Chin. J. Oceanol. Limnol. 2015, 33, 1354–1361. [Google Scholar] [CrossRef]
- Anufriieva, E.V.; Shadrin, N. Diversity of Fauna in Crimean Hypersaline Water Bodies. J. Sib. Fed. Univ. Biol. 2018, 11, 294–305. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Name of Water Body | WGS (X) | WGS (Y) | Area (km2) | Depth (cm) | TOC (mgL−1) | CHL (µgL−1) | TDS (gL−1) | ZOO SUM (ind.100 L−1) | TAX NUM | TD |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Teniz | 64.588 | 51.682 | 6.4 | 70 | 56 | 378 | 6 | 5845 | 15 | 1.63 |
| 2 | Sukyrkol | 64.446 | 51.589 | 1.4 | 60 | 51 | 169 | 4 | 83,983 | 7 | 0.16 |
| 3 | Kaiyndsor | 64.515 | 51.524 | 5.4 | 20 | 48 | 23 | 131 | 799 | 1 | 0 |
| 4 | Asubastysor | 64.460 | 51.520 | 1–1.5 | 20 | 66 | 41 | 88 | 1865 | 1 | 0 |
| 5 | Ukrash | 64.494 | 51.505 | 11.5 | 10 | 48 | 6 | 322 | 17 | 1 | 0 |
| 6 | Zharsor | 64.502 | 51.461 | 6.0 | 10 | 25 | 9 | 70 | 383 | 1 | 0 |
| 7 | Unnamed | 64.311 | 51.357 | 1.5–2 | 20 | 125 | 10 | 79 | 3623 | 1 | 0 |
| 8 | Unnamed | 63.693 | 51.548 | 1–1.5 | 70 | 43 | 8 | 14 | 5428 | 4 | 0.47 |
| 9 | Unnamed | 63.691 | 51.534 | 3–4 | 10 | 74 | 10 | 149 | 200 | 1 | 0 |
| 10 | Unnamed | 63.683 | 51.535 | <1 | 10 | 79 | 47 | 23 | 11,523 | 6 | 1.56 |
| 11 | Unnamed | 63.023 | 51.411 | <1 | 20 | 63 | 57 | 16 | 1818 | 6 | 2.15 |
| 12 | Shoshkakol | 63.030 | 51.370 | 5.1 | 25 | 38 | 34 | 6 | 3144 | 19 | 2.67 |
| 13 | Little Aqsuat | 62.807 | 51.368 | 1.3 | 5 | 25 | 26 | 9 | 108 | 5 | 1.88 |
| 14 | Unnamed | 62.673 | 51.329 | 0.5–1 | 20 | 48 | 6 | 49 | 40 | 2 | 0.91 |
| 15 | Zharman Koli | 62.526 | 51.210 | 56.9 | 5 | 32 | 11 | 54 | 14 | 5 | 2.24 |
| 16 | Zharkol | 62.393 | 51.342 | 3–6 | 100 | 25 | 10 | 2 | 7740 | 15 | 1.46 |
| 17 | Saryqopa Koli | 62.357 | 51.322 | 336 | 50 | 61 | 10 | 45 | 6361 | 5 | 0.73 |
| 18 | Little Tengiz | 64.093 | 50.160 | 454 | 5 | 109 | 5 | 131 | 118 | 1 | 0 |
| 19 | Kalmakty | 69.504 | 50.640 | 7–8 | 10 | 51 | 5 | 147 | 310 | 1 | 0 |
| 20 | Balyksor | 69.744 | 50.643 | 6–7 | 10 | 24 | 27 | 15 | 4400 | 5 | 1.17 |
| 21 | Boshchesor | 70.058 | 50.526 | 5.5–6 | 10 | 52 | 8 | 70 | 96 | 6 | 1.98 |
| 22 | Big Saryoba | 70.134 | 50.313 | 12–13 | 50 | 43 | 9 | 18 | 21,132 | 7 | 1.54 |
| 23 | Karasor | 72.084 | 51.171 | 55–60 | 30 | 12 | 13 | 12 | 1002 | 5 | 1.15 |
| Salinity Category | Name of Water Body |
|---|---|
| Subsaline 0.5–3 gL−1 | Zharkol |
| Hyposaline ˃3–20 gL−1 | Teniz, Sukyrkol, No.8. Unnamed, No.11. Unnamed, Shoshkakol, Little Aqsuat, Balyksor, Big Saryoba, Karasor |
| Mesosaline ˃20–50 gL−1 | No.10. Unnamed, No.14. Unnamed, Saryqopa Koli |
| Hypersaline ˃50 gL−1 | Kaindysor, Asubastysor, Ukrash, Zharsor, No.7. Unnamed, No.9. Unnamed, Zharman Koli, Little Tengiz, Kalmaty, Boshchesor |
| Rotifera | Cladocera | Copepoda | Artemia | ZOO SUM | Tax Num | |
|---|---|---|---|---|---|---|
| Z | 0.30152 | 0.43239 | 0.45722 | −0.24425 | 0.49704 | 0.32947 |
| pH | 0.30115 | 0.05342 | 0.32491 | 0.04822 | 0.36876 | 0.30047 |
| TDS | −0.83494 | −0.69204 | −0.6944 | 0.37294 | −0.58103 | −0.82013 |
| Na | −0.47061 | −0.63707 | −0.41634 | 0.36991 | −0.24111 | −0.57075 |
| K | 0.30152 | 0.43239 | 0.45722 | −0.24425 | 0.49704 | 0.32947 |
| Ca | 0.5931 | 0.56692 | 0.55663 | −0.50667 | 0.21146 | 0.61941 |
| Mg | 0.30833 | 0.45542 | 0.27251 | −0.12011 | 0.25395 | 0.46532 |
| Cl | −0.45595 | −0.34759 | −0.52686 | 0.32298 | −0.24308 | −0.4567 |
| SO4 | 0.3523 | 0.29629 | 0.43703 | −0.30178 | 0.15316 | 0.35836 |
| HCO3 + CO3 | 0.78783 | 0.52871 | 0.62325 | −0.08327 | 0.82016 | 0.65894 |
| TOC | 0.5931 | 0.56692 | 0.55663 | −0.50667 | 0.21146 | 0.61941 |
| TN | 0.30833 | 0.45542 | 0.27251 | −0.12011 | 0.25395 | 0.46532 |
| TP | −0.45595 | −0.34759 | −0.52686 | 0.32298 | −0.24308 | −0.4567 |
| CHL | 0.3523 | 0.29629 | 0.43703 | −0.30178 | 0.15316 | 0.35836 |
| Rotifera | 1 | 0.55685 | 0.67478 | −0.13929 | 0.58001 | 0.86545 |
| Cladocera | 0.55685 | 1 | 0.73413 | −0.5256 | 0.55855 | 0.79349 |
| Copepoda | 0.67478 | 0.73413 | 1 | −0.55309 | 0.76001 | 0.77169 |
| Artemia | −0.13929 | −0.5256 | −0.55309 | 1 | −0.07873 | −0.35025 |
| ZOO SUM | 0.58001 | 0.55855 | 0.76001 | −0.07873 | 1 | 0.54844 |
| Tax Num | 0.86545 | 0.79349 | 0.77169 | −0.35025 | 0.54844 | 1 |
| Name of Inland Water | Zharkol | Sukyrkol | Shoshkakol | Teniz | Little Aqsuat | Karasor | No.8. Unnamed | No.11. Unnamed | Big Saryoba | No.10. Unnamed | Boshchesor |
|---|---|---|---|---|---|---|---|---|---|---|---|
| salinity TDS gL−1 | 2 | 4 | 6 | 6 | 9 | 12 | 14 | 16 | 18 | 23 | 70 |
| Bdelloid unidentified | 267 | 12 | 67 | ||||||||
| Brachionus asplanchnoides Charin, 1947 | 266 | 200 | 133 | ||||||||
| Brachionus dimidiatus Bryce, 1931 | 17 | 513 | |||||||||
| Brachionus plicatilis Müller, 1786 | 67 | 93 | |||||||||
| Brachionus quadridentatus Hermann, 1783 | 40 | 180 | 17 | ||||||||
| Brachionus urceolaris Müller, 1773 | 36 | ||||||||||
| Cephalodella catellina Müller, 1786 | 12 | 100 | |||||||||
| Colurella adriatica Ehrenberg, 1831 | 40 | ||||||||||
| Colurella colurus Ehrenberg, 1830 | 12 | 120 | |||||||||
| Eosphora ehrenbergii Weber and Montet, 1918 | 120 | ||||||||||
| Hexarthra fennica Levander, 1892 | 84 | ||||||||||
| Keratella quadrata Müller, 1786 | 360 | ||||||||||
| Keratella sp. | 2 | ||||||||||
| Lecane luna Müller, 1776 | 40 | ||||||||||
| Lophocharis oxysternon Gosse, 1851 | 36 | 36 | |||||||||
| Lophocharis salpina Ehrenberg, 1834 | 12 | ||||||||||
| Mytilina ventralis Ehrenberg, 1830 | 40 | ||||||||||
| Notholca acuminata Ehrenberg, 1832 | 160 | 266 | 912 | 33 | 18 | ||||||
| Paradicranophorus hudsoni Glascott, 1893 | 33 | 233 | |||||||||
| Paradicranophorus sordidus Donner, 1968 | 200 | 400 | |||||||||
| Synchaeta oblonga Ehrenberg, 1832 | 932 | ||||||||||
| SUM ROTIFERA ind 100 L−1 | 800 | 569 | 1200 | 1399 | 18 | 96 | 333 | 839 | 320 | 533 | 2 |
| Name of Inland Water | Zharkol | Sukyrkol | Shoshkakol | Teniz | Little Aqsuat | Karasor | Balyksor | Big Saryoba | No.10. Unnamed | Saryqopa Koli | Boshchesor |
|---|---|---|---|---|---|---|---|---|---|---|---|
| salinity TDS gL−1 | 2 | 4 | 6 | 6 | 9 | 12 | 15 | 18 | 23 | 45 | 70 |
| Alona affinis Leydig, 1860 | 67 | ||||||||||
| Alona rectangular Sars, 1861 | 12 | 24 | |||||||||
| Ceriodaphnia reticulate Jurine, 1820 | 84 | ||||||||||
| Chydorus latus Sars, 1862 | 24 | ||||||||||
| Chydorus sphaericus Müller, 1776 | 40 | ||||||||||
| Daphnia curvirostris Eylmann and Johnson | 160 | 36 | |||||||||
| Daphnia longispina O.F.Müller, 1776 | 40 | 24 | |||||||||
| Daphnia magna Straus, 1820 | 80 | 33 | 100 | 54 | |||||||
| Moina brachiata Jurine, 1820 | 12 | ||||||||||
| Moina salina Daday, 1888 | 12 | 48 | 2584 | 5280 | 2 | ||||||
| Moina sp. | 32 | ||||||||||
| Scapholeberis ramneri Dumont and Pensaert, | 12 | ||||||||||
| Simocephalus exspinosus De Geer, 1776 | 48 | ||||||||||
| SUM CLADOCERA ind 100 L−1 | 320 | 33 | 252 | 100 | 78 | 12 | 80 | 2584 | 67 | 5280 | 2 |
| Name of Inland Water | Zharkol | Sukyrkol | Shoshkakol | Teniz | Little Aqsuat | Karasor | No.8. Unnamed | Balyksor | No.11. Unnamed | Big Saryoba | No.10. Unnamed | Saryqopa Koli | No.14. Unnamed | Zharman Koli | Boshchesor |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| salinity TDS gL−1 | 2 | 4 | 6 | 6 | 9 | 12 | 14 | 15 | 16 | 18 | 23 | 45 | 49 | 54 | 70 |
| COPEPODA-Calanoida | |||||||||||||||
| Arctodiaptomus bacillifer Koelbel, 1885 | 360 | 132 | |||||||||||||
| Arctodiaptomus salinus Daday, 1885 | 756 | 7600 | 996 | 4 | |||||||||||
| Hemidiaptomus amblyodon Marenzeller, 1873 | 20 | 360 | 746 | ||||||||||||
| Metadiptomus asiaticus Uljanine,1875 | 2432 | 7060 | |||||||||||||
| Cyclopoida | |||||||||||||||
| Cyclops furcifer Claus, 1857 | 266 | ||||||||||||||
| Cyclops strenuus Fischer, 1851 | 33 | 33 | |||||||||||||
| Cyclops scutifer G.O.Sars, 1863 | 17 | ||||||||||||||
| Cyclops vicinus Ulianine, 1875 | 33 | ||||||||||||||
| Diacyclops bisetosus Rehberg, 1880 | 67 | ||||||||||||||
| Megacyclops viridis Jurine, 1820 | 200 | 133 | 20 | ||||||||||||
| Harpacticoida | |||||||||||||||
| Cletocamptus retrogressus Schmankevitch, 1875 | 6 | 48 | 72 | 6 | |||||||||||
| copepodites | 534 | 48 | 34 | 6 | 32 | 4 | |||||||||
| nauplii | 6040 | 82,548 | 1152 | 4096 | 138 | 5028 | 1856 | 93 | 10,560 | 2531 | 12 | 13 | 6 | 62 | |
| SUM COPEPODA ind 100 L−1 | 6620 | 83,381 | 1692 | 4346 | 12 | 894 | 5095 | 4320 | 839 | 18,228 | 9591 | 1080 | 13 | 10 | 72 |
| Name of Inland Water | No.11. Unnamed | No.10. Unnamed | Saryqopa Koli | No.14. Unnamed | Zharman Koli | Zharsor | Boshchesor | No.7. Unnamed | Abubastysor | Kalyndsor | Little Tengiz | Kalmakty | No.9 Unnamed | Ukrash |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| salinity TDS gL−1 | 16 | 23 | 45 | 49 | 54 | 70 | 70 | 79 | 88 | 131 | 131 | 147 | 149 | 322 |
| SUM ARTEMIA ind 100 L−1 | 140 | 1332 | 1 | 27 | 2 | 383 | 20 | 3623 | 1865 | 799 | 118 | 310 | 200 | 17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zsuga, K.; Inelova, Z.; Boros, E. Zooplankton Community Structure in Shallow Saline Steppe Inland Waters. Water 2021, 13, 1164. https://doi.org/10.3390/w13091164
Zsuga K, Inelova Z, Boros E. Zooplankton Community Structure in Shallow Saline Steppe Inland Waters. Water. 2021; 13(9):1164. https://doi.org/10.3390/w13091164
Chicago/Turabian StyleZsuga, Katalin, Zarina Inelova, and Emil Boros. 2021. "Zooplankton Community Structure in Shallow Saline Steppe Inland Waters" Water 13, no. 9: 1164. https://doi.org/10.3390/w13091164
APA StyleZsuga, K., Inelova, Z., & Boros, E. (2021). Zooplankton Community Structure in Shallow Saline Steppe Inland Waters. Water, 13(9), 1164. https://doi.org/10.3390/w13091164