
Fresh- and Brackish-Water Cold-Tolerant Species of Southern Europe: Migrants from the Paratethys That Colonized the Arctic

 ,
,  and
and 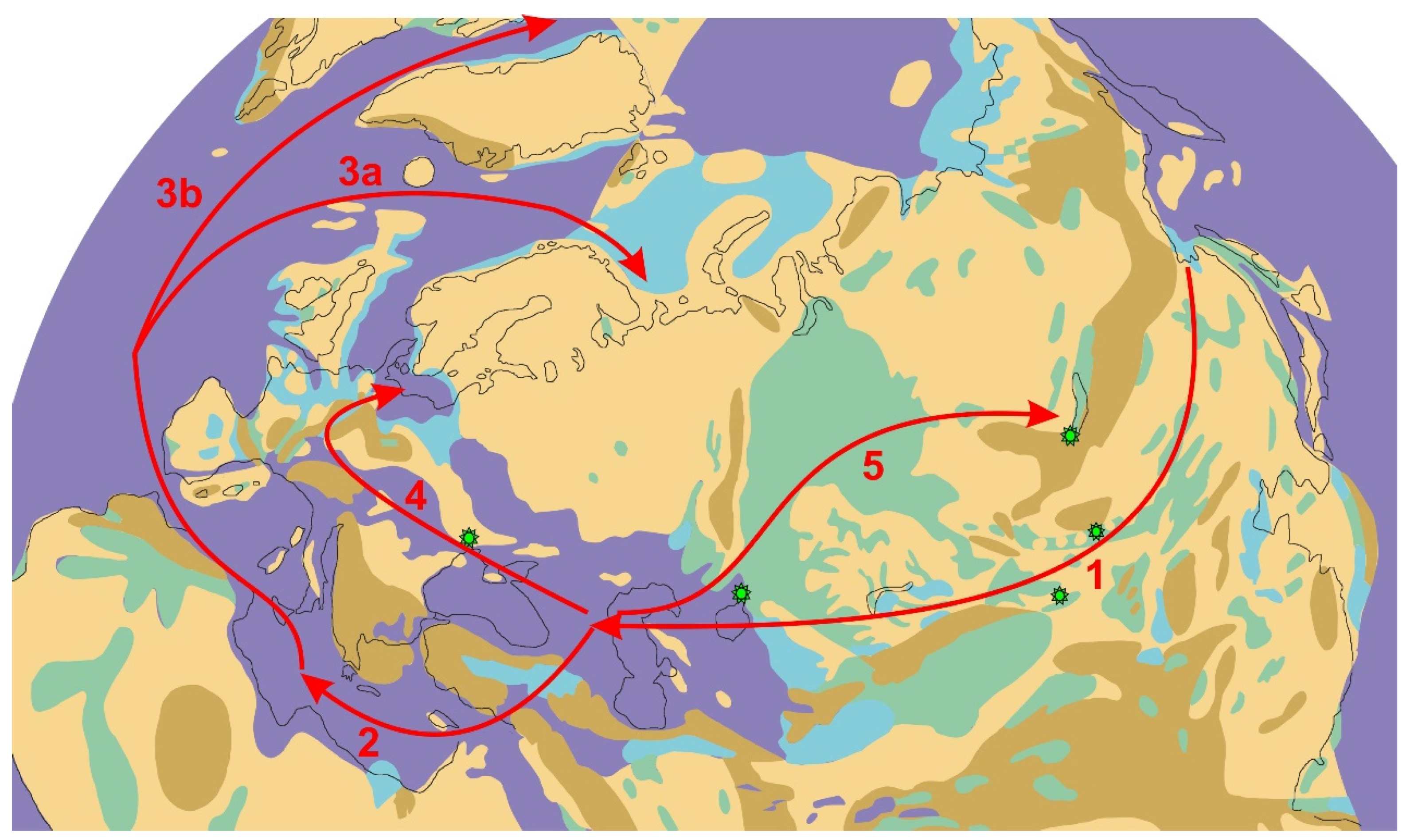{kind=link}
Abstract
1. Introduction
2. Material and Methods
3. Factual Data
3.1. Schizothoracine Fishes (Subfamily Schizothoracinae)
3.2. Nemacheiline Loaches (Subfamily Nemacheilinae)
3.3. Sticklebacks (Family Gasterosteidae)
3.4. Salmonids
3.5. Freshwater Pearl Mussels of the Genus Margaritifera
3.6. Unionid Mussels (Family Unionidae)
3.7. Snails of the Genus Ecrobia Stimpson, 1865 (Family Hydrobiidae)
3.8. Planarians (Order Tricladida)
3.9. Seals (Family Phocidae)
3.10. Cold-Water Hydrobionts of Southern Europe Whose Ways of Dispersal Remain Insufficiently Studied
4. Discussion
4.1. The Routes of Invasion of Cold-Tolerant Hydrobionts from the Pacific Ocean Basin and Central Asia to the Paratethys Basin
4.2. The Routes of the Spread of Cold-Water Species from the Paratethys to the Mediterranean Region
4.3. Dispersal of Cold-Water Species from the Ponto-Caspian and Mediterranean to Northern Europe and North America
4.4. Evolutionary and Ecological Characteristics of Fresh- and Brackish-Water Cold-Tolerant Species of Southern Europe
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simpson, G.G. Splendid Isolation; Yale University Press: New Haven, CT, USA; London, UK, 1980; xi+266p. [Google Scholar]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967; 203p. [Google Scholar]
- Comte, L.; Buisson, T.; Daufresne, M.; Grenouillet, G. Climate-induced changes in the distribution of freshwater fish: Observed and predicted trends. Freshw. Biol. 2013, 58, 625–639. [Google Scholar] [CrossRef]
- Santiago, J.M.; García de Jalón, D.; Alonso, C.; Solana, J.; Ribalaygua, J.; Pórtoles, J.; Monjo, R. Brown trout thermal niche and climate change: Expected changes in the distribution of cold-water fish in central Spain. Ecohydrology 2016, 9, 514–528. [Google Scholar] [CrossRef]
- Abeli, T.; Vamosi, J.C.; Orsenigo, S. The importance of marginal population hotspots of cold-adapted species for research on climate change and conservation. J. Biogeogr. 2018, 45, 977–985. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Makhrov, A.A.; Gofarov, M.Y.; Aksenova, O.V.; Aspholm, P.E.; Bespalaya, Y.V.; Kabakov, M.B.; Kolosova, Y.S.; Kondakov, A.V.; Ofenböck, T.; et al. Climate warming as a possible trigger of keystone mussel population decline in oligotrophic rivers at the continental scale. Sci. Rep. UK 2018, 8, 35. [Google Scholar] [CrossRef]
- Morato, T.; González-Irusta, J.-M.; Dominguez-Carrió, C.; Wei, C.-L.; Davies, A.; Sweetman, A.K.; Taranto, G.H.; Beazley, L.; Garca-Alegre, A.; Grehan, A.; et al. Climate-induced changes in the suitable habitat of cold-water corals and commercially important deep-sea fishes in the North Atlantic. Glob. Chang. Biol. 2020, 26, 2181–2202. [Google Scholar] [CrossRef] [PubMed]
- Regan, C.T. The geographical distribution of salmon and trout. Salmon Trout Mag. 1920, 22, 25–35. [Google Scholar]
- Deng, T.; Wu, F.; Zhou, Z.; Su, T. Tibetan Plateau: An evolutionary junction for the history of modern biodiversity. Sci. China Earth Sci. 2020, 63, 172–187. [Google Scholar] [CrossRef]
- Makhrov, A.A.; Bolotov, I.N.; Artamonova, V.S.; Borovikova, E.A. Mountain regions are the putative place of origin of many Arctic animal and plant forms. Zool. Zh. 2019, 98, 1291–1303. [Google Scholar] [CrossRef]
- Ekman, S. Zoogeography of the Sea; Sidgwick and Jackson Limited: London, UK, 1953; 417p. [Google Scholar]
- Por, F.D. The Legacy of Tethys: An Aquatic Biogeography of the Levant; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1989; 214p. [Google Scholar]
- Hou, Z.; Li, S. Tethyan changes shaped aquatic diversification. Biol. Rev. 2018, 93, 874–896. [Google Scholar] [CrossRef] [PubMed]
- Berg, L.S. Über die amphiboreale (discontinuierliche) Verbreitung der Meeresfauna in der nördlichen Hemisphäre. Zoogeographica 1934, 2, 393–409. [Google Scholar]
- Kosswig, C. Tethys and its relation to the peri-Mediterranean faunas of freshwater fishes. In Aspects of Tethyan Biogeography: A Symposium; Adams, C.G., Ager, D.V., Eds.; The Systematics Association: London, UK, 1967; pp. 313–324. [Google Scholar]
- Kazmin, V.G.; Natapov, L.M. (Eds.) The Paleogeographic Atlas of Northern Eurasia; Institute of Tectonics of Lithospheric Plates: Moscow, Russia, 1998. [Google Scholar]
- Popov, S.V.; Rögl, F.; Rozanov, A.Y.; Steininger, F.F.; Shcherba, I.G.; Kovac, M. (Eds.) Lithological-paleogeographic maps of Paratethys: 10 maps Late Eocene to Pliocene. Cour. Forsch. Senck. 2004, 250, 1–46. [Google Scholar]
- Bianco, P.G. Potential role of the paleohistory of the Mediterranean and Paratethys basin on the early dispersal of Euro-Mediterranean freshwater fishes. Ichthyol. Explor. Freshw. 1990, 1, 167–184. [Google Scholar]
- Stanković, S. The Balkan Lake Ohrid and Its Living World; Junk, W.: Den Haag, The Netherlands, 1960; 357p. [Google Scholar]
- Sychevskaya, E.K. The History of the Northern Eurasia Freshwater Ichthyofauna. Ph.D. Thesis, Paleontological Institute of the Soviet Academy of Sciences, Moscow, Russia, 1991. (In Russian). [Google Scholar]
- Chapsky, K.K. The concept of the Arctic origin of the pinnipeds and alternate way to resolve this problem. In The Arctic Ocean and Its Coastal Zone in Cenozoic; Tolmachev, A.I., Ed.; Gidrometeoizdat: Leningrad, Russia, 1970; pp. 166–173. (In Russian) [Google Scholar]
- Dorofeeva, E.A. Systematics and distribution history of European salmonid fishes of the genus Salmo. J. Ichthyol. 1998, 38, 419–429. [Google Scholar]
- Alekseev, V.R.; Sukhikh, N.M. On time and place of origin of continental calanoids families: A hypothesis. Crustaceana 2020, 93, 245–260. [Google Scholar] [CrossRef]
- Makhrov, A.A.; Vinarski, M.V.; Gofarov, M.Y.; Dvoryankin, G.A.; Novoselov, A.P.; Bolotov, I.N. Faunal Exchanges between the Basins of the Arctic Ocean and the Caspian Sea: Their History and Current Processes. Biol. Bulletin. 2021, 48. in press. [Google Scholar]
- Calatayud, J.; Rodríguez, M.A.; Molina-Venegas, R.; Leo, M.; Horreo, J.L.; Hortal, J. Pleistocene climate change and the formation of regional species pools. Proc. R. Soc. B 2019, 286, 20190291. [Google Scholar] [CrossRef] [PubMed]
- Sworobowicz, L.; Mamos, T.; Grabowski, M.; Wysocka, A. Lasting through the ice age: The role of the proglacial refugia in the maintenance of genetic diversity, population growth, and high dispersal rate in a widespread freshwater crustacean. Freshwater Biol. 2020, 65, 1028–1046. [Google Scholar] [CrossRef]
- Schmitt, T.; Muster, C.; Schönswetter, P. Are disjunct Alpine and Arctic-Alpine animal and plant species in the Western Palearctic really “Relics of a cold past”? In Relict Species. Phylogeography and Conservation Biology; Habel, J.C., Assmann, T., Eds.; Springer: Berlin/Heidelberg, Germany; Dordrecht, The Netherlands; London, UK; New York, NY, USA, 2010; pp. 239–252. [Google Scholar]
- Csapó, H.; Krzywoźniak, P.; Grabowski, M.; Wattier, R.; Bącela-Spychalska, K.; Mamos, T.; Jelić, M.; Rewicz, T. Successful post-glacial colonization of Europe by single lineage of freshwater amphipod from its Pannonian Plio-Pleistocene diversification hotspot. Sci. Rep. UK 2020, 10, 18695. [Google Scholar] [CrossRef] [PubMed]
- Von Richthofen, F.F. China. In Ergebnisse Eigener Reisen und Darauf Gegründeter Studien. Band 1; Verlag von Dietrich Reimer: Berlin, Germany, 1877; p. 758S. [Google Scholar]
- Cowan, P.J. Geographic usage of the terms Middle Asia and Central Asia. J. Arid. Environ. 2007, 69, 359–363. [Google Scholar] [CrossRef]
- Vasnetsov, V.V. Phylogeny of the cyprinid Schizothoracinae of Mountain Asia. Proceedings of A.N. Sev. Inst. Evol. Morphol. Anim. 1950, 2, 3–84. (In Russian) [Google Scholar]
- Timirkhanov, S.R. The Schizothoracine Cyprinids (Schizothoracinae, Cypriniformes, Osteichthyes) of Waterbodies of Middle Asia and Kazakhstan: Morphology, Biology, and Systenatics. Ph.D. Thesis, Institute of Zoology of the Ministry of Education and Science of Kazakhstan, Almaty, Kazakhstan, 2001. (In Russian). [Google Scholar]
- He, D.; Chen, Y.; Chen, Y.; Chen, Z. Molecular phylogeny of the specialized schizothoracine fishes (Teleostei: Cyprinidae), with their implications for the uplift of the Qinghai-Tibetan Plateau. Chin. Sci. Bull. 2004, 49, 39–48. [Google Scholar] [CrossRef]
- Li, Y.; Ren, Z.; Shedlock, A.M.; Wu, J.; Sang, L.; Tersing, T.; Hasegawa, M.; Yonezawa, T.; Zhong, Y. High altitude adaptation of the schizothoracine fishes (Cyprinidae) revealed by the mitochondrial genome analyses. Gene 2013, 517, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, Z.; Zhou, C.; Kong, X. Molecular phylogeny of the subfamily Schizothoracinae (Teleostei: Cypriniformes: Cyprinidae) inferred from complete mitochondrial genomes. Biochem. Syst. Ecol. 2016, 64, 6–13. [Google Scholar] [CrossRef]
- Sychevskaya, E.K. The Freshwater Paleogene Ichthyofauna of the USSR and Mongolia; Nauka: Moscow, Russia, 1986; 157p. (In Russian) [Google Scholar]
- Yang, T.; Zhang, L.; Li, W.; Jia, J.; Han, L.; Zhang, Y.; Chen, Y.; Yan, D. New schizothoracine from Oligocene of Qaidam Basin, northern Tibetan Plateau, China, and its significance. J. Vertebr. Paleontol. 2018, 38, e1442840. [Google Scholar] [CrossRef]
- Berg, L.S. Zoogeography of the freshwater fishes of Western Asia. Uchenyye Zapiski LGU 1940, 56, 3–31. (In Russian) [Google Scholar]
- Esmaeili, H.R.; Mehraban, H.; Abbasi, K.; Keivany, Y.; Coad, B.W. Review and updated checklist of freshwater fishes of Iran: Taxonomy, distribution and conservation status. Iran. J. Ichthyol. 2017, 4 (Suppl. S1), 1–114. [Google Scholar]
- Carrapa, B.; DeCelles, P.G.; Wang, X.; Clementz, M.T.; Mancin, N.; Stoica, M.; Kraatz, B.; Meng, J.; Abdulov, S.; Chen, F. Tectono-climatic implications of Eocene Paratethys regression in the Tajik basin of central Asia. Earth Planet. Sci. Lett. 2015, 424, 168–178. [Google Scholar] [CrossRef]
- Li, Q.; Li, L.; Zhang, Y.; Guo, Z. Oligocene incursion of the Paratethys seawater to the Junggar Basin, NW China: Insight from multiple isotopic analysis of carbonate. Sci. Rep. UK 2020, 10, 6601. [Google Scholar] [CrossRef]
- Prejevalsky, N. Mongolia, the Tangut Country, and the Solitudes of Northern Tibet, Being a Narrative of Three Years’ Travel in Eastern High Asia; Sampson Low, Marston, Searle, & Rivington: London, UK, 1876; Volume 1, 287p. [Google Scholar]
- Tong, C.; Li, M. Genomic signature of accelerated evolution in a saline-alkaline lake-dwelling Schizothoracine fish. Int. J. Biol. Macromol. 2020, 149, 341–347. [Google Scholar] [CrossRef]
- Prokofiev, A.M. Loaches of the Subfamily Nemacheilinae of the World Fauna; Filigran: Yaroslavl, Russia, 2017; 315p. (In Russian) [Google Scholar]
- Freyhof, J.; Erk’akan, F.; Özeren, C.; Perdices, A. An overview of the western Palaearctic loach genus Oxynoemacheilus (Teleostei: Nemacheilidae). Ichthyol. Explor. Freshw. 2011, 22, 301–312. [Google Scholar]
- Šedivá, A.; Apostolou, A.; Kohout, J.; Bohlen, J. Molecular phylogeographic analyses of the loach Oxynoemacheilus bureschi reveal post-glacial range extensions across the Balkans. J. Fish Biol. 2010, 76, 357–368. [Google Scholar] [CrossRef]
- Conway, K.W.; Edds, D.R.; Shrestha, J.; Mayden, R.L. A new species of gravel-dwelling loach (Ostariophysi: Nemacheilidae) from the Nepalese Himalayan foothills. J. Fish Biol. 2011, 79, 1746–1759. [Google Scholar] [CrossRef]
- Sayyadzaden, G.; Eagderi, S.; Esmaeili, H.R. A new loach of the genus Oxynoemacheilus from the Tigris River drainage and its phylogenetic relationships among the nemacheilid fishes (Teleostei: Nemacheilidae) in the Middle East based on mtDNA COI sequences. Iran. J. Ichthyol. 2016, 3, 236–250. [Google Scholar]
- Keivany, Y.; Nelson, J.S. Phylogenetic relationships of sticklebacks (Gasterosteidae), with emphasis on ninespine sticklebacks (Pungitius spp.). Behaviour 2004, 141, 1485–1497. [Google Scholar] [CrossRef]
- Münzing, J. Variabilität, Verbreitung und Systematik der Arten und Unterarten in der Gattung Pungitius Coste, 1848 (Pisces, Gasterosteidae). Z. Zool. Syst. Evolut. Forsch. 1969, 7, 208–233. [Google Scholar] [CrossRef]
- Guo, B.; Fang, B.; Shikano, T.; Momigliano, P.; Wang, C.; Kravchenko, A.; Merilä, J. A phylogenomic perspective on diversity, hybridization and evolutionary affinities in the stickleback genus Pungitius. Molec. Ecol. 2019, 28, 4046–4064. [Google Scholar] [CrossRef] [PubMed]
- Shtyl’ko, B.A. The Neogene freshwater ichthyofauna of Western Siberia. Tr. Vsesoyuznogo Geol. Razvedochnogo ob’edinenia NKTP SSSR 1934, 359, 1–96. (In Russian) [Google Scholar]
- Lebedev, V.D. The Neogene fauna of freshwater fishes of the Zaisan Depression and the Western Siberian Lowland. Voprosy Ikhtiologii. 1959, 12, 28–69. (In Russian) [Google Scholar]
- Wang, C.; Shikano, T.; Persat, H.; Merilä, J. Mitochondrial phylogeography and cryptic divergence in the stickleback genus Pungitius. J. Biogeogr. 2015, 42, 2334–2348. [Google Scholar] [CrossRef]
- Ziuganov, V.V. The Family Gasterosteidae of World Fish Fauna; Nauka: Leningrad, Russia, 1991; 261p. (In Russian) [Google Scholar]
- Bell, M.A.; Foster, S.A. (Eds.) The Evolutionary Biology of the Threespine Stickleback; Oxford University Press: New York, NY, USA; Tokyo, Japan, 1994; 571p. [Google Scholar]
- Bennike, O. Quaternary vertebrates from Greenland: A review. Quat. Sci. Rev. 1997, 16, 899–909. [Google Scholar] [CrossRef]
- Nazarkin, M.V.; Yabumoto, Y.; Urabe, A. A new Miocene three-spined stickleback (Pisces: Gasterosteidae) from Central Japan. Paleontol. Res. 2013, 16, 318–328. [Google Scholar] [CrossRef]
- DeFaveri, J.; Zanella, L.N.; Zanella, D.; Mrakovčić, M.; Merilä, J. Phylogeography of isolated freshwater three-spined stickleback Gasterosteus aculeatus populations in the Adriatic Sea basin. J. Fish Biol. 2012, 80, 61–85. [Google Scholar] [CrossRef]
- Lucek, K.; Seehausen, O. Distinctive insular forms of threespine stickleback (Gasterosteus aculeatus) from western Mediterranean islands. Conserv. Genet. 2015, 16, 1319–1333. [Google Scholar] [CrossRef]
- Sanz, N.; Araguas, R.M.; Vidal, O.; Viñas, J. Glacial refuges for three-spined stickleback in the Iberian Peninsula: Mitochondrial DNA phylogeography. Freshw. Biol. 2015, 60, 1794–1809. [Google Scholar] [CrossRef]
- Vila, M.; Hermida, M.; Fernândez, C.; Perea, S.; Doadrio, I.; Amaro, R.; San Miguel, E. Phylogeography and conservation genetics of the Ibero-Balearic three-Spined stickleback (Gasterosteus aculeatus). PLoS ONE 2017, 12, e0170685. [Google Scholar] [CrossRef] [PubMed]
- Fang, B.; Merilä, J.; Ribeiro, F.; Alexandre, C.M.; Momigliano, P. Worldwide phylogeny of three-spined sticklebacks. Mol. Phylogenet. Evol. 2018, 127, 613–625. [Google Scholar] [CrossRef]
- Artamonova, V.S.; Bardukov, N.V.; Kulish, A.V.; Makhrov, A.A.; Laius, D.L. The origin and dispersal ways of the three-spined stickelback Gasterosteus aculeatus) of Europe. In Biological Diversity: Its Study, Conservation, Restoration, and Rational Use; Arial: Simpheropol, Russia, 2020; pp. 253–256. (In Russian) [Google Scholar]
- Gibson, D.I.; Køie, M. Magnibursatus caudofilamentosa (Reimer, 1971) n. comb. (Digenea: Derogenidae) from the stickleback Gasterosteus aculeatus L. in Danish waters: A zoogeographical anomaly? Syst. Parasitol. 1991, 20, 221–228. [Google Scholar] [CrossRef]
- Kostadinova, A.; Gibson, D.I. New records of rare derogenids (Digenea: Hemiuroidea) from Mediterranean sparids, including the description of a new species of Magnibursatus Naidenova, 1969 and redescription of Derogenes adriaticus Nikolaeva, 1966. Syst. Parasitol. 2009, 74, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Kostadinova, A.; Power, A.M.; Fernández, M.; Balbuena, J.A.; Raga, J.A.; Gibson, D.I. Three species of Magnibursatus Naidenova, 1969 (Digenea: Derogenidae) from Atlantic and Black Sea marine teleosts. Folia Parasitol. 2003, 50, 202–210. [Google Scholar] [CrossRef]
- Gorjanović-Kramberger, D. Palaeoichthyoložki prilozi (Collectae palaeoichthyologicae). Rad Jugoslavenske Akademije Znanosti i Umjetnosti. Knjiga CVI. Razred Mat. Prirodosl. 1891, 12, 59–129. [Google Scholar]
- Tortonese, E. The trouts of Asiatic Turkey. Publ. Hydrobiol. Res. Institure Univ. Istanb. 1955, 2, 1–26. [Google Scholar]
- Balon, E.K. Notes to the origin and evolution of trouts and salmons with special reference to the Danubian trouts. Acta Soc. Zool. Bohemoslov. 1968, 32, 1–21. [Google Scholar]
- Kydersky, L.A. On the origin of salmon and trout (Salmo trutta L.) in the basins of the Aral, Caspian, and Black seas. Izv. GosNIORKH 1974, 97, 187–216. (In Russian) [Google Scholar]
- Rukhkian, R.G. Karyology and Origin of Trouts of Transcaucasia; The Armenian SSR Academy of Sciences Press: Yerevan, Armenia; 166p. (In Russian)
- Simonovic, P.; Maric, S.; Nikolic, V. Trout Salmo spp. complex in Serbia and adjacent regions of the western Balkans: Reconstruction of evolutionary history from external morphology. J. Fish Biol. 2007, 70 (Suppl. SC), 359–380. [Google Scholar] [CrossRef]
- Makhrov, A.A.; Bolotov, I.N. Ecological causes of high morphological plasticity of members of a taxon inhabiting the center of its origin (Exemplified by the Noble Salmons, genus Salmo). Biol. Bulletin. 2019, 46, 38–46. [Google Scholar] [CrossRef]
- Sanz, N. Phylogeographic history of brown trout: A review. In Brown Trout: Biology, Ecology and Management; Lobón-Cerviá, J., Sanz, N., Eds.; Wiley: Hoboken, NJ, USA, 2018; pp. 17–63. [Google Scholar]
- Artamonova, V.S.; Afanasyev, S.A.; Bardukov, N.V.; Golod, V.M.; Kokodiy, S.V.; Koulish, A.V.; Pashkov, A.N.; Pipoyan, S.K.; Reshetnikov, S.I.; Makhrov, A.A. The center of origin and colonization routes of noble salmons of the genus Salmo (Salmonidae, Actinopterigii). Dokl. Biochem. Biophys. 2020, 493, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhang, S.-H.; Wang, D.-Q.; Wu, J.-M.; Wang, C.-Y.; Wei, Q.-W. Conservation genetics assessment and phylogenetic relationships of critically endangered Hucho bleekeri in China. J. Appl. Ichthyol. 2016, 32, 343–349. [Google Scholar] [CrossRef]
- Sychevskaya, E.K. The Freshwater Ichthyofauna of the Neogene of Mongolia; Nauka: Moscow, Russia, 1989; 141p. (In Russian) [Google Scholar]
- Holčík, J.; Hensel, K.; Nieslanik, J.; Skácel, L. The Eurasian Huchen, Hucho Hucho. Largest Salmon in the World; Junk, W., Ed.; Publishers and VEDA: Dordrecht, The Netherlands, 1988; 239p. [Google Scholar]
- Kovalchuk, O.M. The first appearance of Hucho (Salmonidae) in the fossil record of Eastern Europe. Vestn. Zool. 2015, 49, 413–420. [Google Scholar] [CrossRef][Green Version]
- Wang, Y.; Guo, R.; Li, H.; Zhang, X.; Du, J.; Song, Z. The complete mitochondrial genome of the Sichuan taimen (Hucho bleekeri): Repetitive sequences in the control region and phylogenetic implications for Salmonidae. Mar. Genom. 2011, 4, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Artamonova, V.S.; Kolmakova, O.V.; Kirillova, E.A.; Makhrov, A.A. Phylogeny of salmonoid fishes (Salmonoidei) based on mtDNA COI gene sequences (barcoding). Contemp. Probl. Ecol. 2018, 11, 271–285. [Google Scholar] [CrossRef]
- Taeubert, J.-E.; Geist, J. The relationship between the freshwater pearl mussel (Margaritifera margaritifera) and its hosts. Biol. Bull. 2017, 44, 67–73. [Google Scholar] [CrossRef]
- Chepalyga, A.L. Fossil Margaritifera of Moldavia and southwestern Ukraine. Int. Geol. Rev. 1965, 7, 1847–1857. [Google Scholar] [CrossRef]
- Shchelinsky, V.E.; Gurova, M.; Tesakov, A.S.; Titov, V.V.; Frolov, P.D.; Simakova, A.N. The Early Pleistocene site of Kermek in western Ciscaucasia (southern Russia): Stratigraphy, biotic record and lithic industry (preliminary results). Quatern. Int. 2016, 393, 51–69. [Google Scholar] [CrossRef]
- Lyubas, A.A.; Obada, T.F.; Gofarov, M.Y.; Kriauciunas, V.V.; Vikhrev, I.V.; Nicoara, I.N.; Bolotov, I.N. A taxonomic revision of fossil freshwater pearl mussels (Bivalvia: Unionoida: Margaritiferidae) from Pliocene and Pleistocene deposits of Southeastern Europe. Ecol. Montenegrina 2019, 21, 1–16. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Vikhrev, I.V.; Bespalaya, Y.V.; Gofarov, M.Y.; Kondakov, A.V.; Konopleva, E.S.; Bolotov, N.N.; Lyubas, A.A. Multi-locus fossil-calibrated phylogeny, biogeography and a subgeneric revision of the Margaritiferidae (Mollusca: Bivalvia: Unionoida). Mol. Phylogenet. Evol. 2016, 103, 104–121. [Google Scholar] [CrossRef] [PubMed]
- Vikhrev, I.V.; Makhrov, A.A.; Artamonova, V.S.; Ermolenko, A.V.; Gofarov, M.Y.; Kabakov, M.B.; Kondakov, A.V.; Chukhchin, D.G.; Lyubas, A.A.; Bolotov, I.N. Fish hosts, glochidia features and life cycle of the endemic freshwater pearl mussel Margaritifera dahurica from the Amur Basin. Sci. Rep. UK 2019, 9, 8300. [Google Scholar] [CrossRef]
- Maderni, U.N. Mollusks of the Continental Cenozoic of the Turgai Depression and Adjacent Regions; Nedra: Leningrad, Russia, 1990; 192p. (In Russian) [Google Scholar]
- Bogachev, V.V. Materials to the History of the Freshwater Fauna of Eurasia; The Ukrainian Academy of Sciences Press: Kiev, Russia, 1961; 403p. (In Russian) [Google Scholar]
- Deviatkin, E.V.; Liskun, I.G.; Chepalyga, A.L. The freshwater malacofauna of the Pliocene of western Mongolia. In Mesozoic and Cenozoic Fauna of Western Mongolia; Trofimov, B.A., Ed.; Nauka: Moscow, Russia, 1971; pp. 33–47. (In Russian) [Google Scholar]
- Treasurer, J.W.; Turnbull, T. The pathology and seawater performance of farmed Atlantic salmon infected with glochidia of Margaritifera margaritifera. J. Fish Biol. 2000, 57, 858–866. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Kondakov, A.V.; Vikhrev, I.V.; Aksenova, O.V.; Bespalaya, Y.V.; Gofarov, M.Y.; Kolosova, Y.S.; Konopleva, E.S.; Spitsyn, V.M.; Tanmuangpak, K.; et al. Ancient River Inference Explains Exceptional Oriental Freshwater Mussel Radiations. Sci. Rep. UK 2017, 7, 2135. [Google Scholar] [CrossRef]
- Froufe, E.; Bolotov, I.; Aldridge, D.C.; Bogan, A.E.; Breton, S.; Gan, H.M.; Kovivadhi, U.; Riccardi, N.; Secci-Petretto, G.; Sousa, R.; et al. Mesozoic mitogenome rearrangements and freshwater mussel (Bivalvia: Unionoidea) macroevolution. Heredity 2020, 124, 182–196. [Google Scholar] [CrossRef]
- Lopes-Lima, M.; Bolotov, I.N.; Aldridge, D.C.; Fonseca, M.M.; Gan, H.M.; Gofarov, M.Y.; Kondakov, A.V.; Prié, V.; Sousa, R.; Varandas, S.; et al. Expansion and systematics redefinition of the most threatened freshwater mussel family, the Margaritiferidae. Mol. Phylogenet. Evol. 2018, 127, 98–118. [Google Scholar] [CrossRef]
- Froufe, E.; Prié, V.; Faria, J.; Ghamizi, M.; Gonçalves, D.V.; Gürlek, M.E.; Karaouzas, I.; Kebapçi, Ü.; Şereflişan, H.; Sobral, C.; et al. Phylogeny, phylogeography, and evolution in the Mediterranean region: News from a freshwater mussel (Potomida, Unionida). Molec. Phylogenet. Evol. 2016, 100, 322–332. [Google Scholar] [CrossRef]
- Lopes-Lima, M.; Hattori, A.; Kondo, T.; Hee Lee, J.; Ki Kim, S.; Shirai, A.; Hayashi, H.; Usui, T.; Sakuma, K.; Toriya, T.; et al. Freshwater mussels (Bivalvia: Unionidae) from the rising sun (Far East Asia): Phylogeny, systematics, and distribution. Mol. Phylogenet. Evol. 2020, 146, 106755. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.J.; Chen, J.H.; Chen, C.Z.; Sha, J.G.; Lan, X.; Wen, S.X. Supraspecific taxa of the Bivalvia first named, described, and pulished in China (1927–2007). Univ. Kans. Paleontol. Contrib. 2009, 17, 1–157. [Google Scholar]
- Lopes-Lima, M.; Sousa, R.; Geist, J.; Aldridge, D.C.; Araujo, R.; Bergengren, J.; Bespalaya, Y.; Bódis, E.; Burlakova, L.; Van Damme, D.; et al. Conservation status of freshwater mussels in Europe: State of the art and future challenges. Biol. Rev. 2016, 92, 572–607. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Kondakov, A.V.; Konopleva, E.S.; Vikhrev, I.V.; Aksenova, O.V.; Aksenov, A.S.; Bespalaya, Y.V.; Borovskoy, A.V.; Danilov, P.P.; Dvoriankin, G.A.; et al. Integrative taxonomy, biogeography and conservation of freshwater mussels (Unionidae) in Russia. Sci. Rep. UK 2020, 10, 3072. [Google Scholar] [CrossRef] [PubMed]
- Tomilova, A.A.; Lyubas, A.A.; Kondakov, A.V.; Konopleva, E.S.; Vikhrev, I.V.; Gofarov, M.Y.; Ozcan, T.; Altun, A.; Ozcan, G.; Gürlek, M.E.; et al. An endemic freshwater mussel species from the Orontes River basin in Turkey and Syria represents duck mussel’s intraspecific lineage: Implications for conservation. Limnologica 2020, 84, 125811. [Google Scholar] [CrossRef]
- Sha, J.; Vajda, V.; Pan, Y.; Larsson, L.; Yao, X.; Zhang, X.; Wang, Y.; Cheng, X.; Jiang, B.; Deng, S.; et al. Stratigraphy of the Triassic-Jurassic boundary successions of the southern margin of the Junggar basin, Northwestern China. Acta Geol. Sin. 2011, 85, 421–436. [Google Scholar] [CrossRef]
- Mrkvicka, A.C. Populations of Microcondylaea bonellii (Férussac 1827), Unionidae—An european freshwater mussel at rapid decline—and Unio mancus in Istria, Croatia. Ecol. Montenegrina 2018, 17, 53–59. [Google Scholar] [CrossRef]
- Govedič, M.; Govedič, T.B. The distribution of Microcondylaea bonellii (Bivalvia: Unionidae) in Slovenia. Natura Slov. 2019, 21, 5–20. [Google Scholar]
- Schütt, H. Die Molluskenfauna der Süßwässer im Einzugsgebiet des Orontes unter Berücksichtigung benachbarter Flußsysteme. Arch. Molluskenk. 1983, 113, 17–91. [Google Scholar]
- Zieritz, A.; Bogan, A.E.; Froufe, E.; Klishko, O.; Kondo, T.; Kovitvadhi, U.; Kovitvadhi, S.; Lee, J.H.; Lopes-Lima, M.; Pfeiffer, J.M.; et al. Diversity, biogeography and conservation of freshwater mussels (Bivalvia: Unionida) in East and Southeast Asia. Hydrobiologia 2017, 810, 29–44. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Kondakov, A.V.; Konopleva, E.S.; Vikhrev, I.V. A new genus of ultra-elongate freshwater mussels from Vietnam and eastern China (Bivalvia: Unionidae). Ecol. Montenegrina 2021, 39, 1–6. [Google Scholar] [CrossRef]
- Kaya, M.Y.; Dupont-Nivet, G.; Proust, J.N.; Roperch, P.; Bougeois, L.; Meijer, N.; Frieling, J.; Fioroni, C.; Alttiner, S.Ö.; Vardar, E.; et al. Paleogene evolution and demise of the proto-Paratethys Sea in Central Asia (Tarim and Tajik basins): Role of intensified tectonic activity at ca. 41 Ma. Basin Res. 2019, 31, 461–486. [Google Scholar] [CrossRef]
- Kaya, M.Y.; Dupont-Nivet, G.; Proust, J.N.; Roperch, P.; Meijer, N.; Frieling, J.; Fioroni, C.; Alttiner, S.Ö.; Stoica, M.; Aminov, J.; et al. Cretaceous evolution of the Central Asian proto-Paratethys Sea: Tectonic, eustatic, and climatic controls. Tectonics 2020, 39, e2019TC005983. [Google Scholar] [CrossRef]
- Vinarski, M.V.; Bolotov, I.N.; Aksenova, O.V.; Babushkin, E.S.; Bespalaya, Y.V.; Makhrov, A.A.; Nekhaev, I.O.; Vikhrev, I.V. Freshwater Mollusca of the Circumpolar Arctic: A review on their taxonomy, diversity and biogeography. Hydrobiologia 2020, in press. [Google Scholar] [CrossRef]
- Tomilova, A.A.; Lyubas, A.A.; Kondakov, A.V.; Vikhrev, I.V.; Gofarov, M.Y.; Kolosova, Y.S.; Vinarski, M.V.; Palatov, D.M.; Bolotov, I.N. Evidence for Plio-Pleistocene duck mussel refugia in the Azov Sea river basins. Diversity 2020, 12, 118. [Google Scholar] [CrossRef]
- Vandendorpe, J.; van Baak, C.G.C.; Stelbrink, B.; Delicado, D.; Albrecht, C.; Wilke, T. Historical faunal exchange between the Pontocaspian Basin and North America. Ecol. Evol. 2019, 9, 10816–10827. [Google Scholar] [CrossRef]
- Wesselingh, F.P.; Neubauer, T.A.; Anistratenko, V.V.; Vinarski, M.V.; Yanina, T.; ter Poorten, J.J.; Kijashko, P.; Albrecht, C.; Anistratenko, O.Y.; D’ Hont, A.; et al. Mollusc species from the Pontocaspian region—An expert opinion list. ZooKeys 2019, 827, 31–124. [Google Scholar] [CrossRef] [PubMed]
- Haase, M.; Naser, M.D.; Wilke, T. Ecrobia grimmi in brackish Lake Sawa, Iraq: Indirect evidence for long-distance dispersal of hydrobiid gastropods (Caenogastropoda: Rissooidea) by birds. J. Moll. Stud. 2010, 76, 101–105. [Google Scholar] [CrossRef]
- Glöer, P.; Pešić, V. The freshwater snails (Gastropoda) of Iran, with descriptions of two new genera and eight new species. ZooKeys 2012, 219, 11–61. [Google Scholar] [CrossRef] [PubMed]
- Asnafi, M.R. Biostratigraphy of Miocene sediments of southern Caspian sea. In Proceedings of the The 1st International Applied Geological Congress, Mashhad, Iran, 26–28 April 2010; Department of Geology, Islamic Azad University: Mashhad, Iran, 2010; p. 1025. [Google Scholar]
- Büyükmeriç, Y.; Tekįn, E.; Herece, E.; Sözerį, K.; Akça, N.; Varol, B. Early Pliocene molluscs from the easternmost Mediterranean region (SE Turkey): Biostratigraphic, ecostratigraphic, and palaeobiogeographic implications. Turk. J. Earth Sci. 2018, 27, 127–151. [Google Scholar] [CrossRef]
- Sluys, R.; Ribas, M.; Baguñà, J. Taxonomy, ecology, and karyology of a new species of Phagocata from Spain, with a discussion on the phylogenetic systematics of the genus Phagocata s.1. (Platyhelminthes, Tricladida, Paludicola). Can. J. Zool. 1995, 73, 557–568. [Google Scholar] [CrossRef]
- Porfirieva, N.A. Planarians of the Baikal Lake; Nauka: Novosibirsk, Russia, 1977; 207p. (In Russian) [Google Scholar]
- Koretsky, I.A.; Rahmat, S.J. First records of fossil Cystophorinae (Carnivora, Phocidae): Middle miocene seals from the Northern Paratethys. Riv. Ital. Paleontol. S. 2013, 119, 325–350. [Google Scholar]
- Koretsky, I.A.; Barnes, L.S. Pinniped evolutionary history and paleobiogeography. In Mesozoic and Cenozoic Vertebrates and Paleoenvironments. Tributes to the Career of Prof. Dan Grigorescu; Csiki, Z., Ed.; Ars Docendi: Bucharest, Romania, 2006; pp. 143–153. [Google Scholar]
- Arnason, U.; Gullberg, A.; Janke, A.; Kullberg, M.; Lehman, N.; Petrov, E.A.; Väinölä, R. Pinniped phylogeny and a new hypothesis for their origin and dispersal. Mol. Phylogenet. Evol. 2006, 41, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Palo, J.U.; Väinölä, R. The enigma of the landlocked Baikal and Caspian seals addressed through phylogeny of phocine mitochondrial sequences. Biol. J. Linn. Soc. 2006, 88, 61–72. [Google Scholar] [CrossRef]
- Fulton, T.L.; Strobeck, C. Multiple fossil calibrations, nuclear loci and mitochondrial genomes provide new insight into biogeography and divergence timing for true seals (Phocidae, Pinnipedia). J. Biogeogr. 2010, 37, 814–829. [Google Scholar] [CrossRef]
- Gadzhiev, D.V. An outline of the formation of the vertabrate fauna of Azerbaijan. In Animal World of Azerbaijan; Gadzhiev, D.V., Rakhmatulina, I.K., Eds.; Elm: Baku, Azerbaijan, 2000; Volume 3, pp. 5–42. (In Russian) [Google Scholar]
- Kaygorodova, I.A.; Sherbakov, D.Y. Molecular Phylogenetic Study of the Systematic Position of Baikalian Oligochaetes in Clitellata. Russ. J. Genet. 2006, 42, 1390–1397. [Google Scholar] [CrossRef]
- Rodriguez, P.; Giani, N. A preliminary review of the taxonomic characters used for the systematics of the genus Trichodrilus Claparède (Oligochaeta, Lumbriculidae). Hydrobiologia 1994, 278, 35–51. [Google Scholar] [CrossRef]
- Potapova, M.G. Occurrence of two Krsticiella species in Beringia suggests this genus is a Cenozoic relict. Diatom Res. 2019, 34, 109–114. [Google Scholar] [CrossRef]
- Artamonova, V.S.; Kucheryavyy, A.V.; Makhrov, A.A. Nucleotide sequence diversity of the mitochondrial cytochrome oxidase subunit I (COI) gene of the Arctic lamprey (Lethenteron camtschaticum) in the Eurasian part of the range. Hydrobiologia 2015, 757, 197–208. [Google Scholar] [CrossRef]
- Makhrov, A.A.; Popov, I.Y. Life forms of lampreys (Petromyzontidae) as a manifestation of intraspecific diversity of ontogenesis. Russ. J. Dev. Biol. 2015, 46, 196–207. [Google Scholar] [CrossRef]
- Levin, B.; Ermakov, A.; Ermakov, O.; Levina, M.; Sarycheva, O.; Sarychev, V. Ukranian brook lamprey Eudontomyzon mariae (Berg): Phylogenetic position, genetic diversity, distribution, and some data on biology. In Jawless Fishes of the World; Orlov, A., Beamish, R., Eds.; Cambridge Scholars Publishing: Newcastle upon Tyne, UK, 2016; pp. 58–82. [Google Scholar]
- Tutman, P.; Freyhof, J.; Dulčič, J.; Glamuzina, B.; Geiger, M. Lampetra soljani, a new brook lamprey from the southern Adriatic Sea basin (Petromyzontiformes: Petromyzontidae). Zootaxa 2017, 4273, 531–548. [Google Scholar] [CrossRef]
- Tuniyev, S.; Naseka, A.; Renaud, C. Review of Western Transcaucasian brook lamprey, Lethenteron ninae Naseka, Tunyev & Renaud, 2009 (Petromyzontidae). In Jawless Fishes of the World; Orlov, A., Beamish, R., Eds.; Cambridge Scholars Publishing: Newcastle upon Tyne, UK, 2016; Volume 2, pp. 154–190. [Google Scholar]
- Berg, L.S. A review of the lampreys of the northern hemisphere. Annu. Musée Zool. l’Académie Sci. l’URSS 1931, 32, 87–116. [Google Scholar]
- Lang, N.J.; Roe, K.J.; Renaud, C.B.; Gill, H.S.; Potter, I.C.; Freyhof, J.; Naseka, A.M.; Cochran, P.; Perez, H.E.; Habit, E.M.; et al. Novel relationships among lampreys (Petromyzontiformes) revealed by a taxonomically comprehensive molecular data set. Am. Fish. Soc. Symp. 2009, 72, 41–55. [Google Scholar]
- Gvozdev, E.V.; Mitrofanov, V.P. (Eds.) Fishes of Kazakhstan, in Five Volumes; Nauka: Alma-Ata, Kazakhstan, 1986; Volume 1, 272p. (In Russian) [Google Scholar]
- Pietschmann, V. Lota hulai, eine neue Fischart aus dem Wiener Becken. Palaeontol. Z. 1934, 16, 48–52. [Google Scholar] [CrossRef]
- Van Houdt, J.K.; Hellemans, B.; Volckaert, F.A.M. Phylogenetic relationships among Palearctic and Nearctic burbot (Lota lota): Pleistocene extinctions and recolonization. Mol. Phylogenet. Evol. 2003, 29, 599–612. [Google Scholar] [CrossRef]
- Weiss, S.J.; Gonçalves, D.V.; Secci-Petretto, G.; Englmaier, G.K.; Gomes-Dos-Santos, A.; Denys, G.P.J.; Persat, H.; Antonov, A.; Hahn, C.; Taylor, E.B.; et al. Global systematic diversity, range distributions, conservation and taxonomic assessments of graylings (Teleostei: Salmonidae; Thymallus spp.). Org. Divers. Evol. 2020, in press. [Google Scholar] [CrossRef]
- Koskinen, M.T.; Ranta, E.; Piironen, J.; Veselov, A.; Titov, S.; Haugen, T.O.; Nilsson, J.; Carlstein, M.; Primmer, C.R. Genetic lineages and postglacial colonization of grayling (Thymallus thymallus, Salmonidae) in Europe, as revealed by mitochondrial DNA analyses. Mol. Ecol. 2000, 9, 1609–1624. [Google Scholar] [CrossRef] [PubMed]
- Sušnik, S.; Snoj, A.; Dovč, P. Evolutionary distinctness of grayling (Thymallus thymallus) inhabiting the Adriatic river system, as based on mtDNA variation. Biol. J. Linn. Soc. 2001, 74, 375–385. [Google Scholar] [CrossRef][Green Version]
- Weiss, S.; Persat, H.; Eppe, R.; Schlötterer, C.; Uiblein, F. Complex patterns of colonization and refugia revealed for European grayling Thymallus thymallus, based on complete sequencing of the mitochondrial DNA control region. Molec. Ecol. 2002, 11, 1393–1407. [Google Scholar] [CrossRef]
- Marić, S.; Kalamujić, B.; Snoj, A.; Razpet, A.; Lukić-Bilela, L.; Pojskić, N.; Bajec, S.S. Genetic variation of European grayling (Thymallus thymallus) populations in the Western Balkans. Hydrobiologia 2012, 691, 225–237. [Google Scholar]
- Marić, S.; Askeyev, I.V.; Askeyev, O.V.; Monakhov, S.P.; Bravničar, J.; Snoj, A. Phylogenetic and population genetic analysis of Thymallus thymallus (Actinopterygii, Salmonidae) from the middle Volga and upper Ural drainage. Hydrobiologia 2014, 740, 167–176. [Google Scholar] [CrossRef]
- Meraner, A.; Gandolfi, A. Phylogeography of European grayling, Thymallus thymallus (Actinopterygii, Salmonidae), within the Northern Adriatic basin: Evidence for native and exotic mitochondrial DNA lineages. Hydrobiologia 2012, 693, 205–221. [Google Scholar] [CrossRef]
- Rückert-Ülkümen, N. Otolithen aus dem Mio-Pliozän von Yalova bei Istanbul, Türkei. N. Jb. Geol. Paläont. 2006, 10, 577–594. [Google Scholar] [CrossRef] [PubMed]
- Palandačić, A.; Naseka, A.; Ramler, D.; Ahnelt, H. Contrasting morphology with molecular data: An approach to revision of species complexes based on the example of European Phoxinus (Cyprinidae). BMC Evol. Biol. 2017, 17, 184. [Google Scholar] [CrossRef]
- Mieszkowska, A.; Górniak, M.; Jurczak-Kurek, A.; Ziętara, M.S. Revision of Gyrodactylus salaris phylogeny inspired by new evidence for Eemian crossing between lineages living on grayling in Baltic and White sea basins. PeerJ 2018, 6, e5167. [Google Scholar] [CrossRef]
- Vinarski, M.V.; Palatov, D.M.; Glöer, P. Revision of ‘Horatia’ snails (Mollusca, Gastropoda, Hydrobiidae sensu lato) from South Caucasus with description of two new genera. J. Nat. Hist. 2014, 48, 2237–2253. [Google Scholar] [CrossRef]
- Vinarski, M.V.; Palatov, D.M. A survey of the Belgrandiella-like gastropods of the Northern Black Sea region (Mollusca, Gastropoda, Hydrobiidae s.l.): Morphological variability and morphospecies. Zool. Zh. 2019, 98, 988–1002. [Google Scholar]
- Grego, J.; Mumladze, L.; Falniowski, A.; Osikowski, A.; Rysiewska, A.; Palatov, D.; Hofman, S. Revealing the stygobiont and crenobiont Mollusca biodiversity hotspot in Caucasus: Part I. The phylogeny of stygobiont Sadlerianinae Szarowska, 2006 (Mollusca; Gastropoda; Hydrobiidae) from Georgia with description of five new genera and twenty-one new species. ZooKeys 2020, 955, 1–77. [Google Scholar]
- Radoman, P. Hydrobioidea—A superfamily of Prosobranchia (Gastropoda). I. Systematics. Monogr. Serb. Acad. Sci. Arts. 1983, 547, 1–256. [Google Scholar]
- Szarowska, M. Molecular phylogeny, systematics and morphological character evolution in the Balkan Rissooidea (Caenogastropoda). Folia Malacol. 2006, 14, 99–168. [Google Scholar] [CrossRef]
- Aksenova, O.V.; Bolotov, I.N.; Gofarov, M.Y.; Kondakov, A.V.; Vinarski, M.V.; Bespalaya, Y.V.; Kolosova, Y.S.; Palatov, D.M.; Sokolova, S.E.; Spitsyn, V.M.; et al. Species richness, molecular taxonomy and biogeography of the radicine pond snails (Gastropoda: Lymnaeidae) in the Old World. Sci. Rep. UK 2018, 8, 11199. [Google Scholar] [CrossRef] [PubMed]
- Gray, J. Evolution of the freshwater ecosystem: The fossil record. Palaeogeog. Palaeocl. 1988, 62, 1–214. [Google Scholar]
- Vinarski, M.V.; Aksenova, O.V.; Bolotov, I.N. Taxonomic assessment of genetically-delineated species of radicine snails (Mollusca, Gastropoda, Lymnaeidae). Zoosyst. Evol. 2020, 96, 577–608. [Google Scholar] [CrossRef]
- Clewing, C.; von Oheimb, P.V.; Vinarski, M.; Wilke, T.; Albrecht, C. Freshwater mollusc diversity at the roof of the world: Phylogenetic and biogeographical affinities of Tibetan Plateau. Valvata J. Molluscan Stud. 2014, 80, 452–455. [Google Scholar] [CrossRef]
- Bǎnǎrescu, P. Zoogeography of Fresh Waters. Vol. 2. Distribution and Dispersal of Freshwater Animals in North. America and Eurasia; AULA-Verlag: Wiesbaden, Germany, 1991; 1091p. [Google Scholar]
- Martinson, G.G. The problem of the origin of Lake Baikal fauna. Zool. Zh. 1967, 46, 1594–1598. (In Russian) [Google Scholar]
- Bǎnǎrescu, P.M. On the affinities and derivation of the aquatic fauna of High Asia. Rev. Roum. Biol. Ser. Biol. Anim. 1983, 28, 97–101. [Google Scholar]
- Kovalchuk, O.; Ferraris, C.J. Late Cenozoic catfishes of Southeastern Europe with inference to their taxonomy and palaeogeography. Palaeontol. Electron. 2016, 19, 1–17. [Google Scholar] [CrossRef]
- Naseka, A.M. Zoogeographical freshwater divisions of the Caucasus as a part of the west Asian transitional region. Proc. Zool. Inst. RAS 2010, 314, 469–492. [Google Scholar]
- Wu, Z.; Barosh, P.J.; Wu, Z.; Hu, D.; Zhao, X.; Ye, P. Vast early Miocene lakes of the central Tibetan Plateau. Geol. Soc. Am. Bull. 2008, 120, 1326–1337. [Google Scholar]
- DeCelles, P.G.; Castañeda, I.S.; Carrapa, B.; Liu, J.; Quade, J.; Leary, R.; Zhang, L. Oligocene-Miocene Great Lakes in the India-Asia Collision Zone. Basin Res. 2018, 30, 228–247. [Google Scholar] [CrossRef]
- Dgebuadze, Y.Y. (Ed.) Limnology and Paleolimnology of Mongolia, 2nd ed.; The Joint Russian-Mongolian Complex Biological Expedition RAS and MAS: Moscow, Russia, 2014; 412p. (In Russian) [Google Scholar]
- Liu, W.; Liu, Z.; An, Z.; Sun, J.; Chang, H.; Wang, N.; Dong, J.; Wang, H. Late Miocene episodic lakes in the arid Tarim Basin, western China. Proc. Natl. Acad. Sci. USA 2014, 111, 16292–16296. [Google Scholar] [CrossRef] [PubMed]
- Nevesskaya, L.A. (Ed.) Paleogeography and Biogeography of Paratethys Basins. Part. 1. Late Eocene—Early Miocene; Scientific World: Moscow, Russia, 2009; 200p. (In Russian) [Google Scholar]
- Krijgsman, W.; Palcu, D.V.; Andreetto, F.; Stoica, M.; Mandic, O. Changing seas in the late Miocene Northern Aegean: A Paratethyan approach to Mediterranean basin evolution. Earth Sci. Rev. 2020, 210, 103386. [Google Scholar] [CrossRef]
- Amin, O.M.; Heckmann, R.A.; Evans, R.P.; Tepe, Y. A description of Echinorhynchus baeri Kostylew, 1928 (Acanthocephala: Echinorhynchidae) from Salmo trutta in Turkey, with notes on synonymy, geographical origins, geological history, molecular profile, and X-ray microanalysis. Parasite 2016, 23, 56. [Google Scholar] [CrossRef][Green Version]
- Mordukhay-Boltovskoy, P.D. Caspian fauna in fresh water outside the Ponto-Caspian Basin. Hydrobiologia 1964, 23, 159–164. [Google Scholar] [CrossRef]
- Albrecht, C.; Wilke, T. Ancient Lake Ohrid: Biodiversity and evolution. Hydrobiologia 2008, 615, 103–140. [Google Scholar] [CrossRef]
- Wysocka, A.; Grabowski, M.; Sworobowicz, L.; Mamos, T.; Burzynski, A.; Sell, J. Origin of the Lake Ohrid gammarid species flock: Ancient local phylogenetic lineage diversification. J. Biogeogr. 2014, 41, 1758–1768. [Google Scholar] [CrossRef]
- Panagiotopoulos, K.; Holtvoeth, J.; Kouli, K.; Marinova, E.; Francke, A.; Cvetkovska, A.; Jovanovska, E.; Lacey, J.H.; Lyons, E.T.; Buckel, C.; et al. Insights into the evolution of the young Lake Ohrid ecosystem and vegetation succession from a southern European refugium during the Early Pleistocene. Quaternary Sci. Rev. 2020, 227, e106044. [Google Scholar] [CrossRef]
- Klishko, O.K.; Kovychev, E.V.; Vinarski, M.V.; Bogan, A.E.; Yurgenson, G.A. The Pleistocene-Holocene aquatic molluscs as indicators of the past ecosystem changes in Transbaikalia (Eastern Siberia, Russia). PLoS ONE 2020, 15, e0235588. [Google Scholar] [CrossRef] [PubMed]
- Wesselingh, F.P.; Alçiçek, H. A new cardiid bivalve from the pliocene Baklan basin (Turkey) and the origin of modern Ponto-Caspian taxa. Palaeontology 2010, 53, 711–719. [Google Scholar] [CrossRef]
- Zander, C.D. Parasite diversity of sticklebacks from the Baltic Sea. Parasitol. Res. 2007, 100, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Veuille, M. L’évolution du genre Jaera Leach (Isopodes; Asellotes) et ses rapports avec l’histoire de la Méditerranée. Bijdr. Dievkunde 1979, 49, 195–217. [Google Scholar] [CrossRef]
- Riedl, R. Neue nordatlantische Formen von adriatishen Schlammböden. Zool. Anz. 1960, 165, 297–311. [Google Scholar]
- Nevesskaya, L.A.; Goncharova, I.A.; Ilyina, L.B.; Popov, S.V. The evolutionary transformations of malacofauna in the Neogene basins of Paratethys as an example of development of island-like ecosystems. Zh. Obshch. Biol. 2009, 70, 396–414. (In Russian) [Google Scholar]
- Taktakishvili, I.G. The Historical Development of the Family Valenciennidae; Metsniereba: Tbilisi, Georgia, 1967; 194p. (In Russian) [Google Scholar]
- Bouchet, P.; Rocroi, J.P.; Hausdorf, B.; Kaim, A.; Kano, Y.; Nützel, A.; Parkhaev, P.; Schrödl, M.; Strong, E.E. Revised classification, nomenclator and typification of gastropod and monoplacophoran families. Malacologia 2017, 61, 1–526. [Google Scholar] [CrossRef]
- Bolotov, I.; Bespalaya, Y.; Aksenova, O.; Aksenov, A.; Bolotov, N.; Gofarov, M.; Kondakov, A.; Paltser, I.; Vikhrev, I. A taxonomic revision of two local endemic Radix spp. (Gastropoda: Lymnaeidae) from Khodutka geothermal area, Kamchatka, Russian Far East. Zootaxa 2014, 3869, 585–593. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Aksenova, O.V.; Bespalaya, Y.V.; Gofarov, M.Y.; Kondakov, A.V.; Paltser, I.S.; Stefansson, A.; Travina, O.V.; Vinarski, M.V. Origin of a divergent mtDNA lineage of a freshwater snail species, Radix balthica, in Iceland: Cryptic glacial refugia or a postglacial founder event? Hydrobiologia 2017, 787, 73–98. [Google Scholar] [CrossRef]
- Vinarski, M.V.; Aksenova, O.V.; Bespalaya, Y.V.; Kondakov, A.V.; Tomilova, A.A.; Khrebtova, I.S.; Gofarov, M.Y.; Bolotov, I.N. One Beringian genus less: A re-assesment of Pacifimyxas Kruglov & Starobogatov, 1985 (Mollusca: Gastropoda: Lymnaeidae) questions the current estimates of Beringian biodiversity. J. Zool. Syst. Evol. Res. 2021, 59, 44–59. [Google Scholar]
- Makhrov, A.A. Decreased evolutionary plasticity as a result of phylogenetic immobilization and its ecological significance. Contemp. Probl. Ecol. 2019, 12, 405–417. [Google Scholar] [CrossRef]
- Makhrov, A.A.; Artamonova, V.S. Instability stabilized: Mechanisms of evolutionary stasis and genetic diversity accumulation in fishes and lampreys from environments with unstable abiotic factors. Contemp. Probl. Ecol. 2020, 13, 370–381. [Google Scholar] [CrossRef]
- Ketelaars, H.A.M. Range extensions of Ponto-Caspian aquatic invertebrates in continental Europe. In Aquatic Invasions in the Black, Caspian, and Mediterranean Seas; Dumont, H., Shiganova, T.A., Niermann, U., Eds.; Kluwer Academic Publishers: New York, NY, USA, 2004; pp. 209–236. [Google Scholar]
- MacIsaac, H.J.; Grigorovich, I. Ponto-Caspian invaders in the Great Lakes: Editorial. J. Great Lakes Res. 1999, 25, 1–2. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Artamonova, V.S.; Bolotov, I.N.; Vinarski, M.V.; Makhrov, A.A. Fresh- and Brackish-Water Cold-Tolerant Species of Southern Europe: Migrants from the Paratethys That Colonized the Arctic. Water 2021, 13, 1161. https://doi.org/10.3390/w13091161
Artamonova VS, Bolotov IN, Vinarski MV, Makhrov AA. Fresh- and Brackish-Water Cold-Tolerant Species of Southern Europe: Migrants from the Paratethys That Colonized the Arctic. Water. 2021; 13(9):1161. https://doi.org/10.3390/w13091161
Chicago/Turabian StyleArtamonova, Valentina S., Ivan N. Bolotov, Maxim V. Vinarski, and Alexander A. Makhrov. 2021. "Fresh- and Brackish-Water Cold-Tolerant Species of Southern Europe: Migrants from the Paratethys That Colonized the Arctic" Water 13, no. 9: 1161. https://doi.org/10.3390/w13091161
APA StyleArtamonova, V. S., Bolotov, I. N., Vinarski, M. V., & Makhrov, A. A. (2021). Fresh- and Brackish-Water Cold-Tolerant Species of Southern Europe: Migrants from the Paratethys That Colonized the Arctic. Water, 13(9), 1161. https://doi.org/10.3390/w13091161