
Assessment of Water Quality Based on Trophic Status and Nutrients-Chlorophyll Empirical Models of Different Elevation Reservoirs




Abstract
1. Introduction
2. Materials and Methods
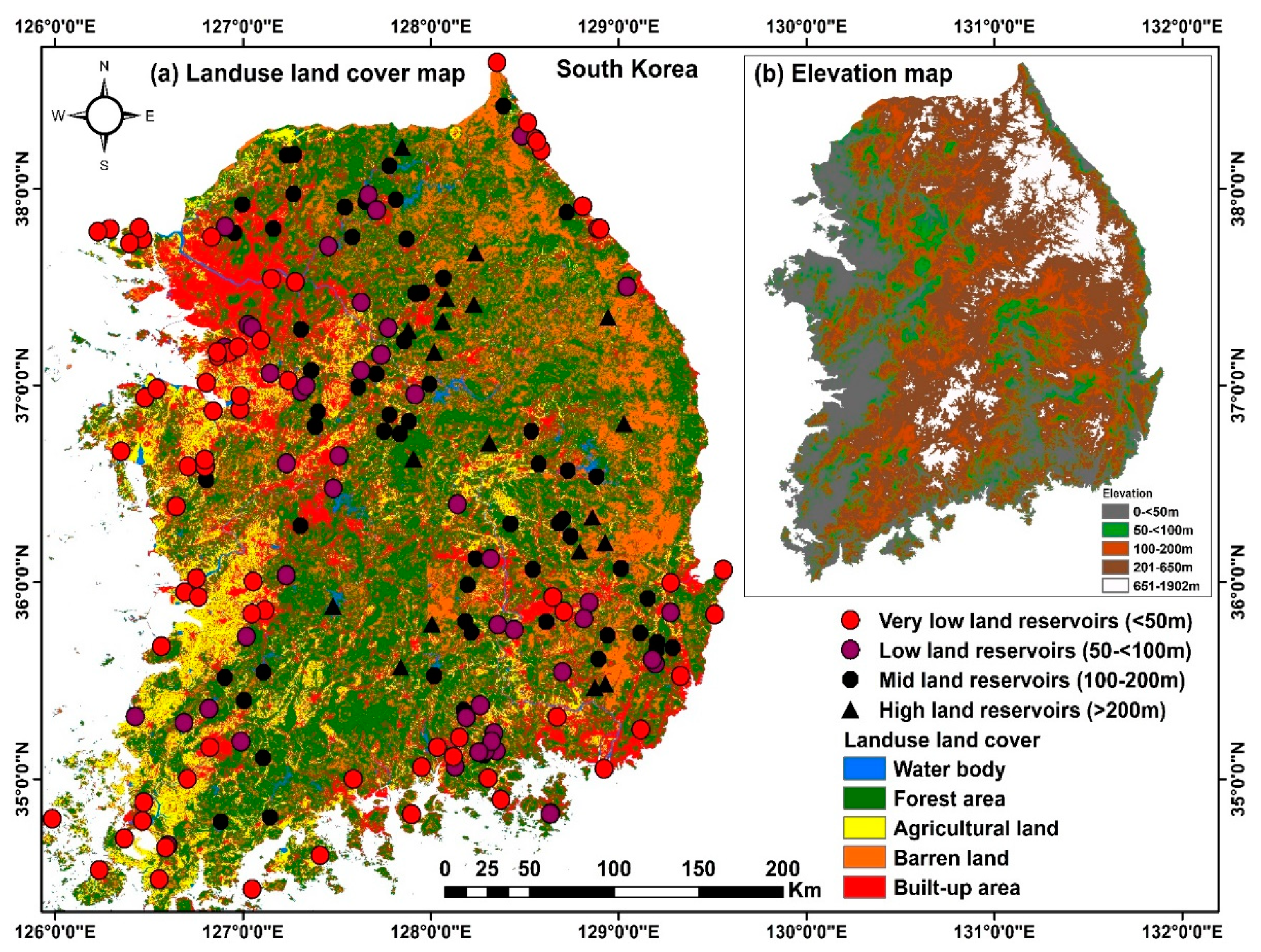
2.1. The Study Region
2.2. Data Source and Analysis of Water Quality
2.3. Trophic Status Index Deviation
2.4. Non-Algal Turbidity
2.5. Statistical Analysis
3. Results
3.1. Physicochemical Characteristics of Korean Reservoirs
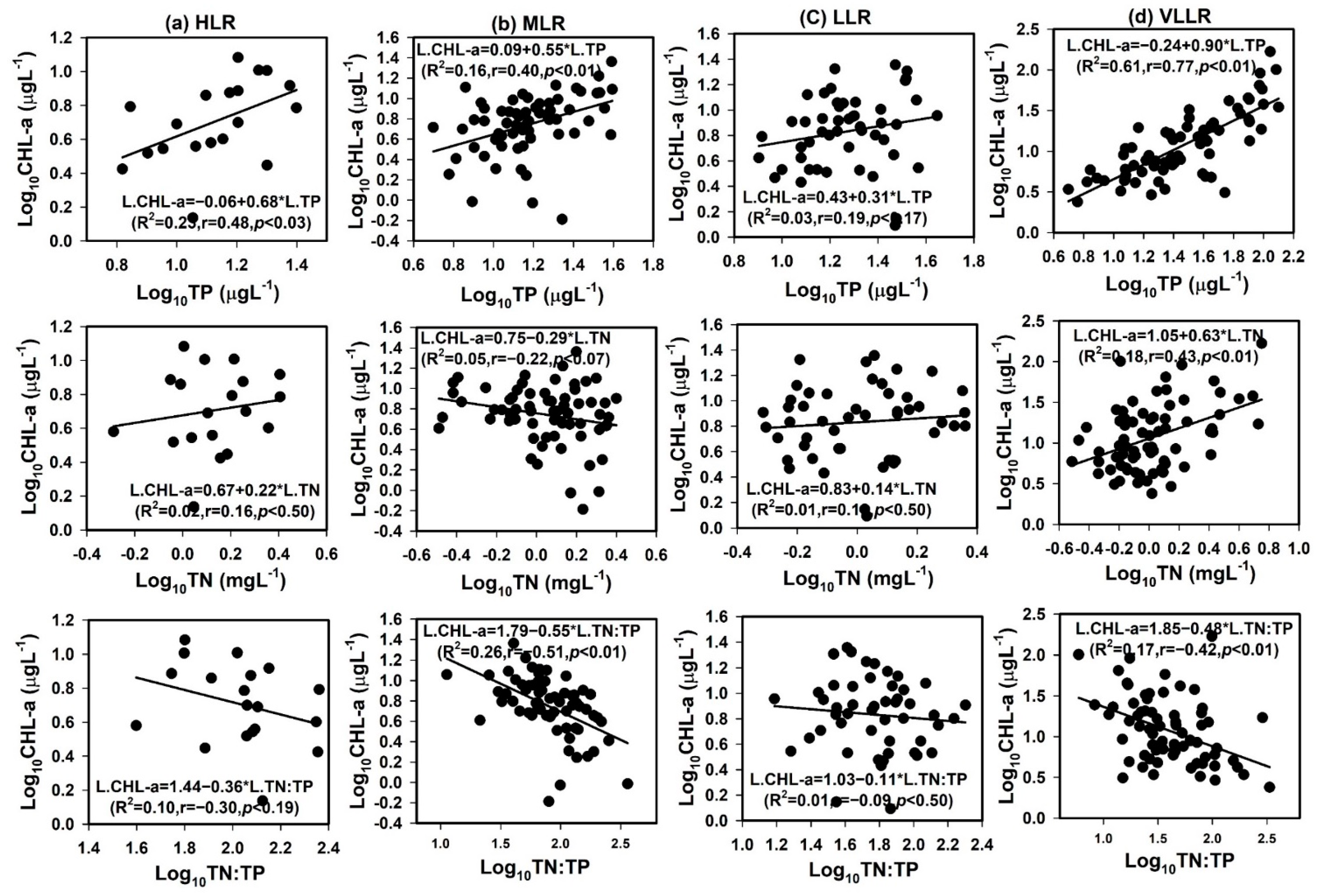
3.2. Assessment of Nutrients, NAT and Algal CHL-a
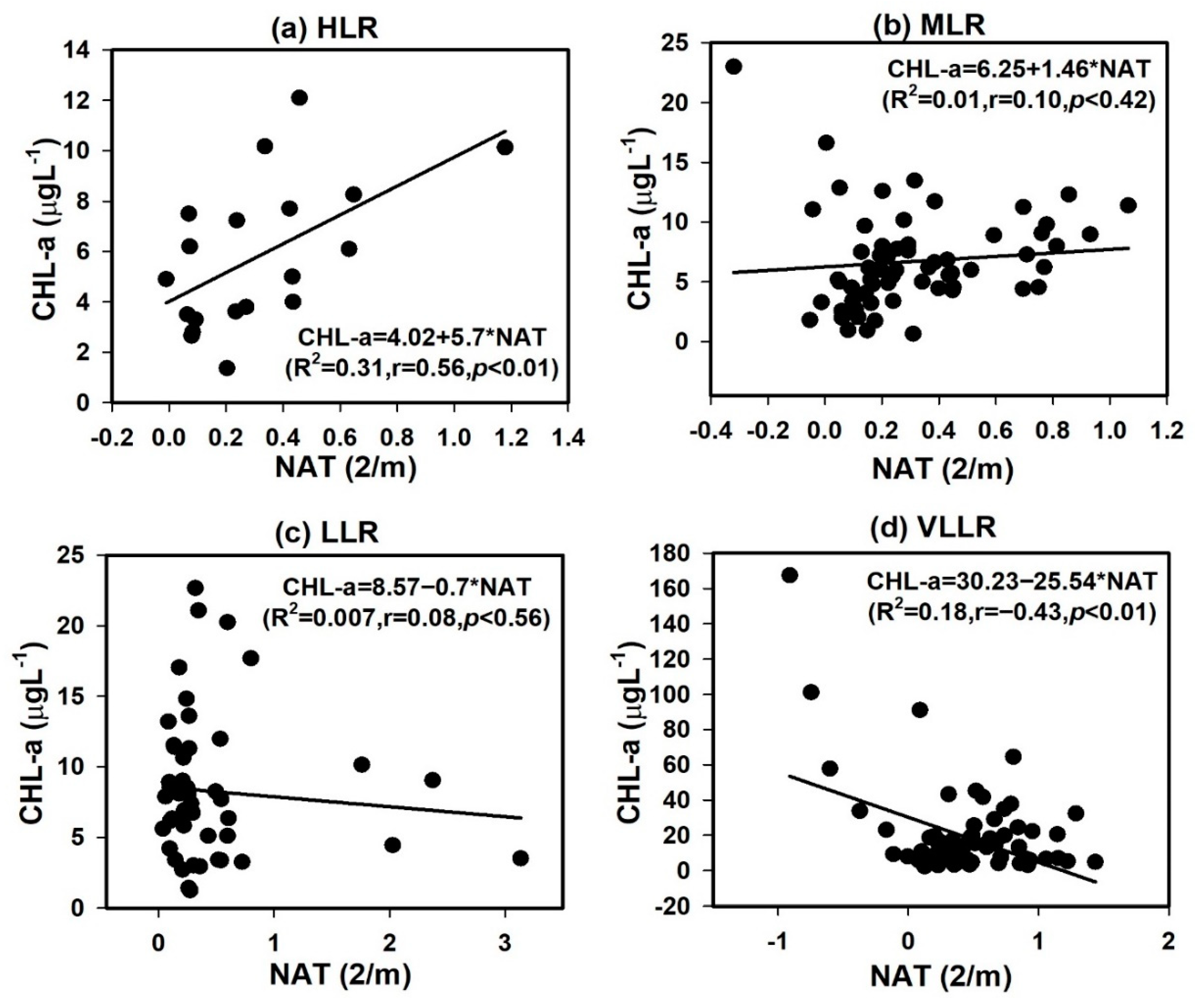
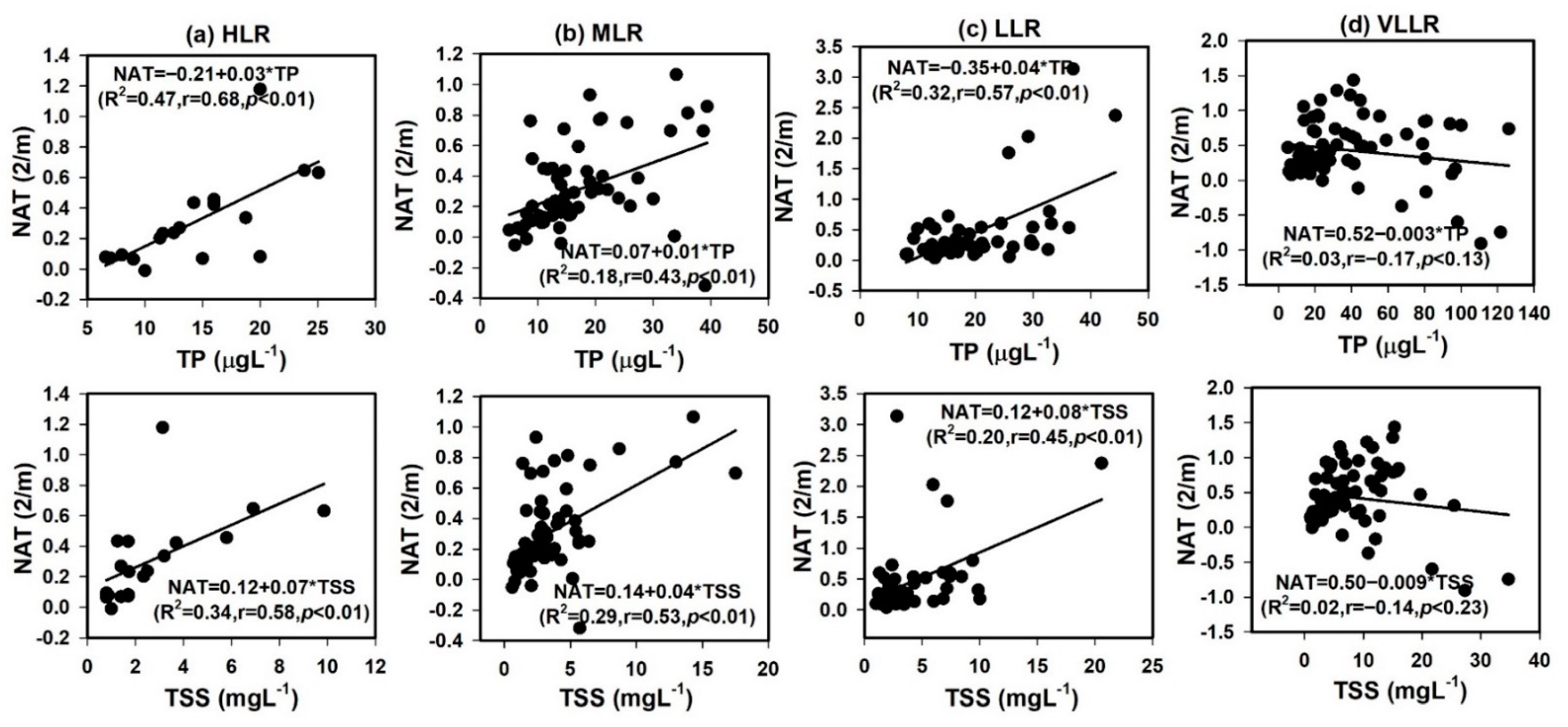
3.3. Water Clarity, NAT, and Other Variables
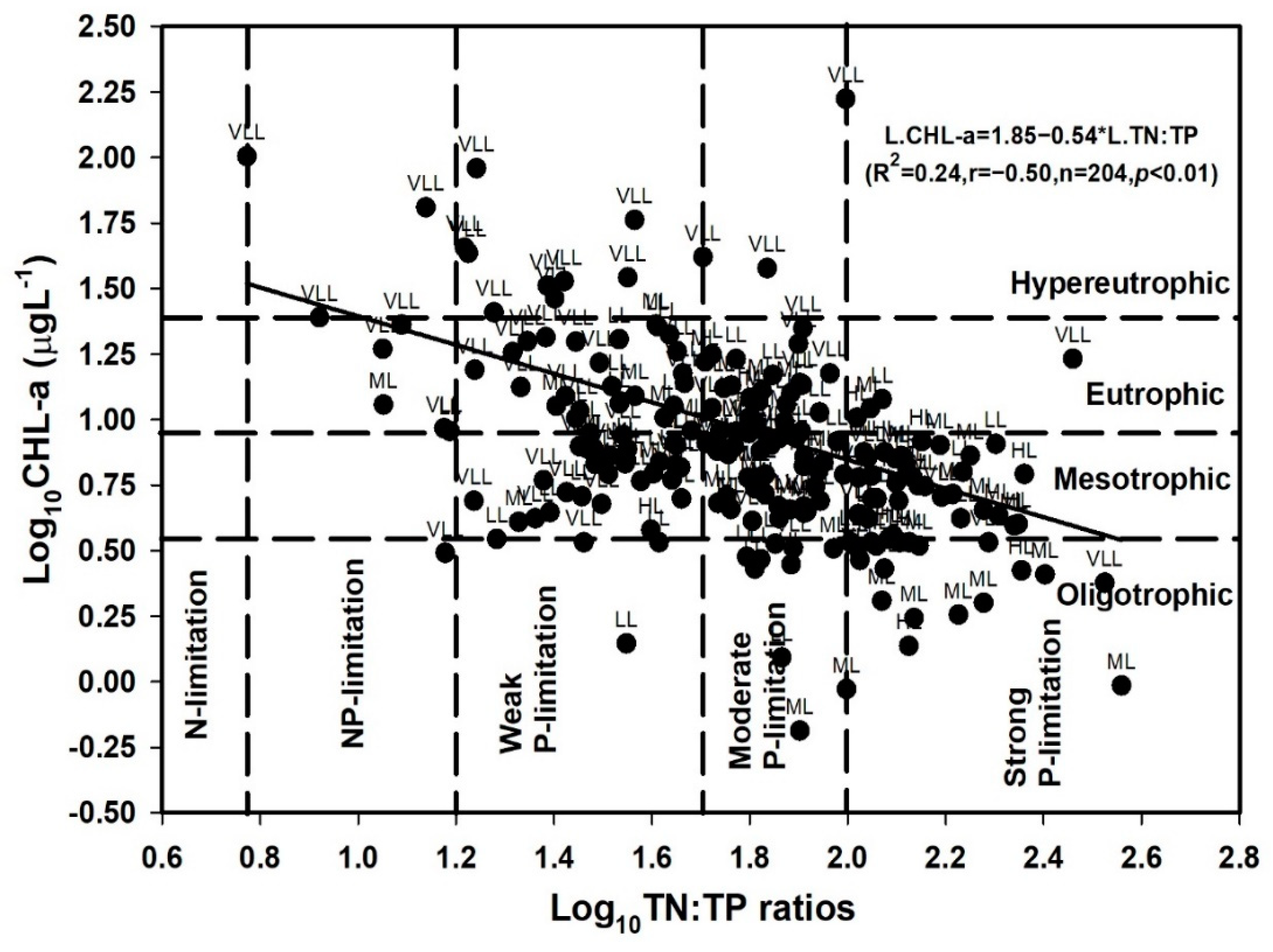
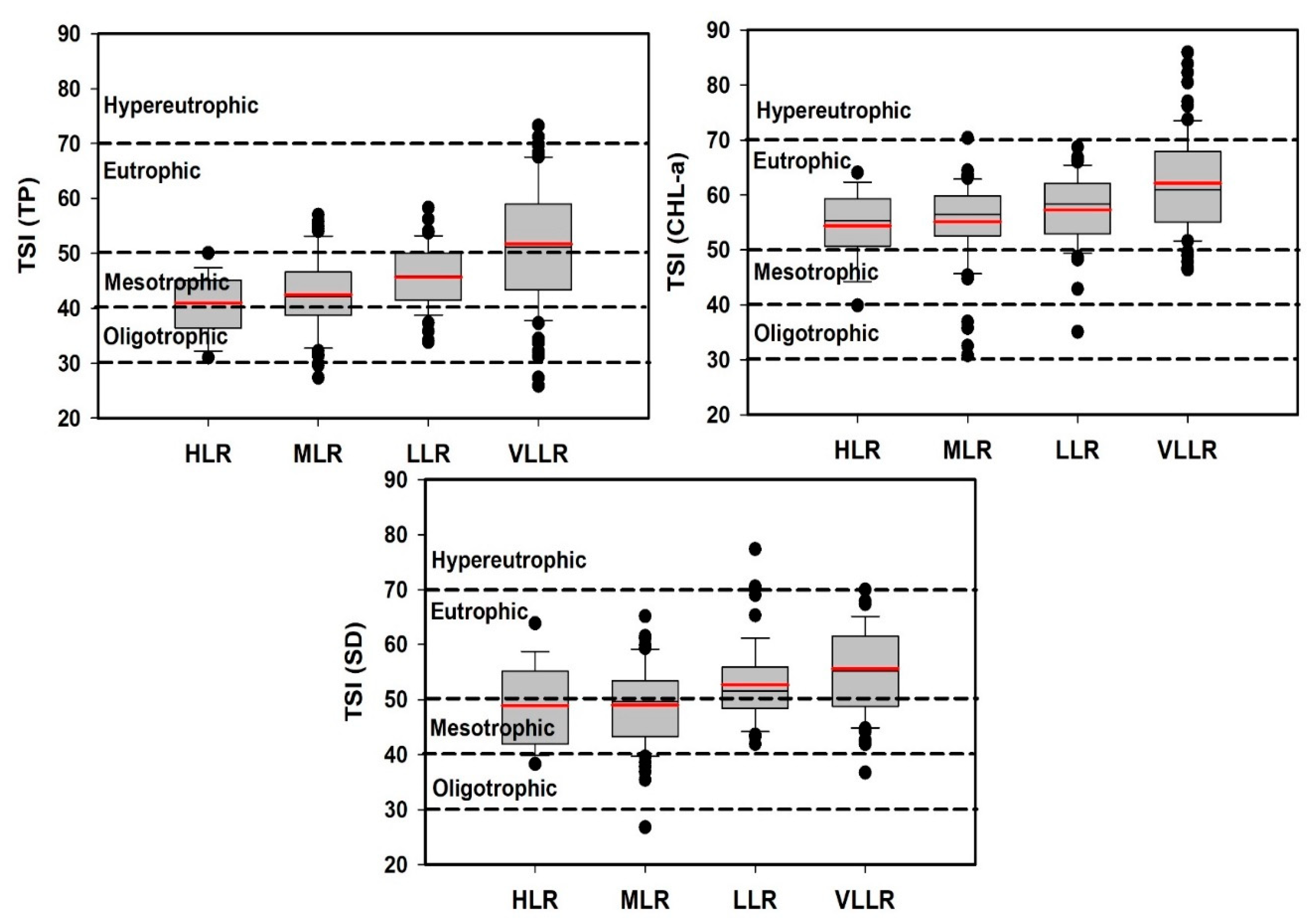
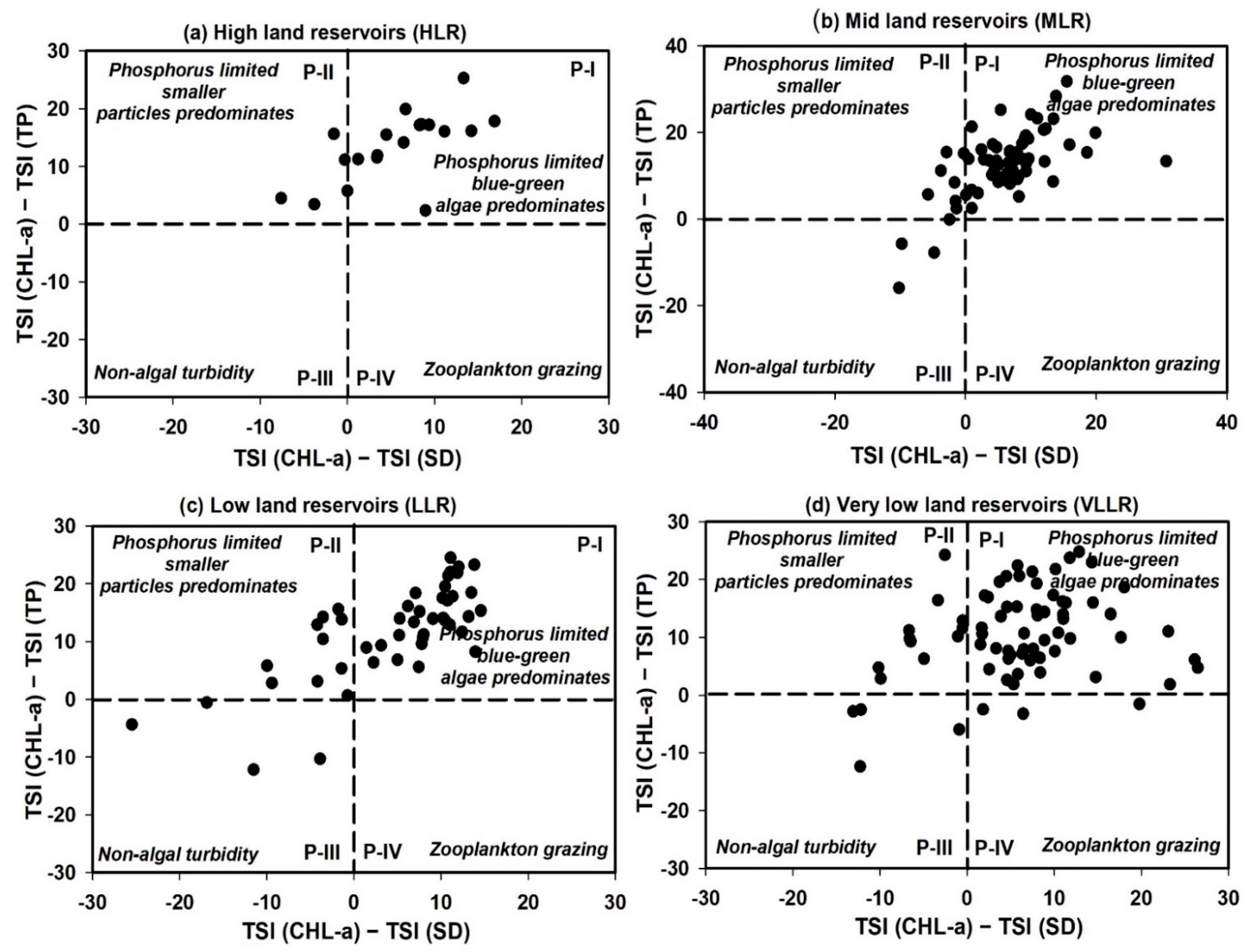
3.4. Trophic Status Index and Its Deviation
4. Discussion
4.1. Reservoirs Physicochemical Water Quality Attributes
4.2. Limiting Factors for Algal Growth
4.3. Light Availability with Nutrients, Solids, and Algal Chlorophyll
4.4. Trophic Status Evaluation of Reservoirs with Its Deviation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cunha, D.G.F.; do Carmo Calijuri, M.; Lamparelli, M.C. A trophic state index for tropical/subtropical reservoirs (TSItsr). Ecol. Eng. 2013, 60, 126–134. [Google Scholar] [CrossRef]
- Cunha, D.G.F.; Finkler, N.R.; Lamparelli, M.C.; do Carmo Calijuri, M.; Dodds, W.K.; Carlson, R.E. Characterizing Trophic State in Tropical/Subtropical Reservoirs: Deviations among Indexes in the Lower Latitudes. Environ. Manag. 2021, 68, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Atique, U.; Mamun, M.; An, K.G. Long-term interannual and seasonal links between the nutrient regime, sestonic chlorophyll and dominant bluegreen algae under the varying intensity of monsoon precipitation in a drinking water reservoir. Int. J. Environ. Res. Public Health 2021, 18, 2871. [Google Scholar] [CrossRef]
- Mamun, M.; Kwon, S.; Kim, J.E.; An, K.G. Evaluation of algal chlorophyll and nutrient relations and the N:P ratios along with trophic status and light regime in 60 Korea reservoirs. Sci. Total Environ. 2020, 741, 140451. [Google Scholar] [CrossRef]
- Mamun, M.; An, K.G. Major nutrients and chlorophyll dynamics in Korean agricultural reservoirs along with an analysis of trophic state index deviation. J. Asia-Pacific Biodivers. 2017, 10, 183–191. [Google Scholar] [CrossRef]
- Lacoul, P.; Freedman, B. Physical and chemical limnology of 34 lentic waterbodies along a tropical-to-alpine altitudinal gradient in Nepal. Int. Rev. Hydrobiol. 2005, 90, 254–276. [Google Scholar] [CrossRef]
- Abell, J.M.; Özkundakci, D.; Hamilton, D.P.; Jones, J.R. Latitudinal variation in nutrient stoichiometry and chlorophyll-nutrient relationships in lakes: A global study. Fundam. Appl. Limnol. 2012, 181, 1–14. [Google Scholar] [CrossRef]
- Jones, J.R.; Knowlton, M.F.; Obrecht, D.V. Role of land cover and hydrology in determining nutrients in mid-continent reservoirs: Implications for nutrient criteria and management. Lake Reserv. Manag. 2008, 24, 1–9. [Google Scholar] [CrossRef]
- Jones, J.R.; Obrecht, D.V.; Perkins, B.D.; Knowlton, M.F.; Thorpe, A.P.; Watanabe, S.; Bacon, R.R. Nutrients, seston, and transparency of missouri reservoirs and oxbow lakes: An analysis of regional limnology. Lake Reserv. Manag. 2008, 24, 155–180. [Google Scholar] [CrossRef]
- Jones, J.R.; Knowlton, M.F.; An, K.G. Trophic state, seasonal patterns and empirical models in South Korean Reservoirs. Lake Reserv. Manag. 2003, 19, 64–78. [Google Scholar] [CrossRef]
- Laperriere, J.D.; Simpson, T.D.; Jones, J.R. Comparative Limnology of Some Lakes in Interior Alaska. Lake Reserv. Manag. 2003, 19, 122–132. [Google Scholar] [CrossRef][Green Version]
- Hwang, S.-J.; Kwun, S.-K.; Yoon, C.-G. Water quality and limnology of Korean reservoirs. Paddy Water Environ. 2003, 1, 43–52. [Google Scholar] [CrossRef]
- Sarin, M.M.; Krishnaswami, S. Major ion chemistry of the Ganga-Brahmaputra river systems, India. Nature 1984, 312, 538–541. [Google Scholar] [CrossRef]
- Sarin, M.M.; Krishnaswami, S.; Dilli, K.; Somayajulu, B.L.K.; Moore, W.S. Major ion chemistry of the Ganga-Brahmaputra river system: Weathering processes and fluxes to the Bay of Bengal. Geochim. Cosmochim. Acta 1989, 53, 997–1009. [Google Scholar] [CrossRef]
- Ahmad, T.; Khanna, P.P.; Chakrapani, G.J.; Balakrishnan, S. Geochemical characteristics of water and sediment of the Indus River, Trans-Himalaya, India: Constraints on weathering and erosion. J. Asian Earth Sci. 1998, 16, 333–346. [Google Scholar] [CrossRef]
- Atique, U.; An, K.G. Landscape heterogeneity impacts water chemistry, nutrient regime, organic matter and chlorophyll dynamics in agricultural reservoirs. Ecol. Indic. 2020, 110, 105813. [Google Scholar] [CrossRef]
- An, K.G.; Jones, J.R. Factors regulating bluegreen dominance in a reservoir directly influenced by the asian monsoon. Hydrobiologia 2000, 432, 37–48. [Google Scholar] [CrossRef]
- Sakamoto, M. Primary production by phytoplankton community in some Japanese lakes and its dependence on lake depth. Arch. Hydrobiol. 1966, 62, 1–28. [Google Scholar]
- Jargal, N.; Lee, H.S.; An, K.G. Long-term water quality patterns in an estuarine reservoir and the functional changes in relations of trophic state variables depending on the construction of serial weirs in upstream reaches. Int. J. Environ. Res. Public Health 2021, 18, 2568. [Google Scholar] [CrossRef] [PubMed]
- Guildford, S.J.; Hecky, R.E. Total nitrogen, total phosphorus, and nutrient limitation in lakes and oceans: Is there a common relationship? Limnol. Oceanogr. 2000, 45, 1213–1223. [Google Scholar] [CrossRef]
- Smith, V.H.; Smith, L.H. Predictive models for the biomass of blue-green algae in lakes. Water Resour. Bull. 1985, 21, 433–439. [Google Scholar] [CrossRef]
- Redfield, A.C. The biological control of chemical factors in the environment. Am. Sci. 1958, 46, 205–212. [Google Scholar]
- Carlson, R.E. A trophic state index for lakes. Limnol. Oceanogr. 1977, 22, 361–369. [Google Scholar] [CrossRef]
- Cunha, D.G.F.; Benassi, S.F.; de Falco, P.B.; do Carmo Calijuri, M. Trophic State Evolution and Nutrient Trapping Capacity in a Transboundary Subtropical Reservoir: A 25-Year Study. Environ. Manag. 2016, 57, 649–659. [Google Scholar] [CrossRef] [PubMed]
- An, K.G.; Park, S.S. Influence of seasonal monsoon on the trophic state deviation in an Asian reservoir. Water Air Soil Pollut. 2003, 145, 267–287. [Google Scholar] [CrossRef]
- Carlson, R.E.; Simpson, J. A Coordinator’s Guide to Volunteer Lake Monitoring Methods. N. Am. Lake Manag. Soc. 1996, 96, 1–92. [Google Scholar]
- Smith, V.H.; Schindler, D.W. Eutrophication science: Where do we go from here? Trends Ecol. Evol. 2009, 24, 201–207. [Google Scholar] [CrossRef]
- Wang, L.; Liu, L.; Zheng, B. Eutrophication development and its key regulating factors in a water-supply reservoir in North China. J. Environ. Sci. 2013, 25, 962–970. [Google Scholar] [CrossRef]
- Atique, U.; An, K.G. Reservoir water quality assessment based on chemical parameters and the chlorophyll dynamics in relation to nutrient regime. Polish J. Environ. Stud. 2019, 28, 1043–1061. [Google Scholar] [CrossRef]
- Mamun, M.; An, K.G. Stream health assessment using chemical and biological multi-metric models and their relationships with fish trophic and tolerance indicators. Ecol. Indic. 2020, 111, 106055. [Google Scholar] [CrossRef]
- MOE. Standard Methods for the Examination of Water Quality Contamination, 7th ed.; Ministry of Environemnt (MOE): Gwacheon, Korea, 2000; p. 435. (In Korean)
- Hara, J.; Atique, U.; An, K.G. Multiyear links between water chemistry, algal chlorophyll, drought-flood regime, and nutrient enrichment in a morphologically complex reservoir. Int. J. Environ. Res. Public Health 2020, 17, 3139. [Google Scholar] [CrossRef]
- Carlson, R.E.; Havens, K.E. Simple graphical methods for the interpretation of relationships between trophic state variables. Lake Reserv. Manag. 2005, 21, 107–118. [Google Scholar] [CrossRef]
- Jones, J.R.; Hubbart, J.A. Empirical estimation of non-chlorophyll light attenuation in Missouri reservoirs using deviation from the maximum observed value in the Secchi-Chlorophyll relationship. Lake Reserv. Manag. 2011, 27, 1–5. [Google Scholar] [CrossRef]
- Walker, W.W., Jr. Probablistic Methods in Lake Management. Models and Software for Reservoir Eutrophication Assessment. Lake Reserv. Manag. 1986, 2, 143–148. [Google Scholar] [CrossRef]
- Nadarajah, S.; Wijenayake, W.M.H.K.; Amarasinghe, U.S. Influence of hydrology on water quality and trophic state of irrigation reservoirs in Sri Lanka. Lakes Reserv. Res. Manag. 2019, 24, 287–298. [Google Scholar] [CrossRef]
- Skjelkvile, L.B.; Wright, R.F. Mountain Lakes; Sensitivity to Acid Deposition and Global Climate Change. Ambio 1998, 27, 280–286. [Google Scholar]
- Drever, J.I.; Zobrist, J. Chemical weathering of silicate rocks as a function of elevation in the southern Swiss Alps. Geochim. Cosmochim. Acta 1992, 56, 3209–3216. [Google Scholar] [CrossRef]
- Tiberti, R.; Tartari, G.A.; Marchetto, A. Geomorphology and hydrochemistry of 12 Alpine lakes in the Gran Paradiso National Park, Italy. J. Limnol. 2010, 69, 242–256. [Google Scholar] [CrossRef]
- Laperriere, J.D.; Jones, J.R.; Swanson, D.K. Limnology of Lakes in Gates of the Arctic national Park and Preserve, Alaska. Lake Reserv. Manag. 2003, 19, 108–121. [Google Scholar] [CrossRef]
- Paerl, H.W.; Gardner, W.S.; Havens, K.E.; Joyner, A.R.; McCarthy, M.J.; Newell, S.E.; Qin, B.; Scott, J.T. Mitigating cyanobacterial harmful algal blooms in aquatic ecosystems impacted by climate change and anthropogenic nutrients. Harmful Algae 2016, 54, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Rigosi, A.; Carey, C.C.; Ibelings, B.W.; Brookes, J.D. The interaction between climate warming and eutrophication to promote cyanobacteria is dependent on trophic state and varies among taxa. Limnol. Ocean 2014, 59, 99–144. [Google Scholar] [CrossRef]
- Scavia, D.; DePinto, J.V.; Bertani, I. A multi-model approach to evaluating target phosphorus loads for Lake Erie. J. Great Lakes Res. 2016, 42, 1139–1150. [Google Scholar] [CrossRef]
- Filstrup, C.T.; Downing, J.A. Relationship of chlorophyll to phosphorus and nitrogen in nutrient-rich lakes. Inl. Waters 2017, 7, 385–400. [Google Scholar] [CrossRef]
- Markad, A.T.; Landge, A.T.; Nayak, B.B.; Inamdar, A.B.; Mishra, A.K. Trophic state modeling for shallow freshwater reservoir: A new approach. Environ. Monit. Assess. 2019, 191, 586. [Google Scholar] [CrossRef]
- Dzialowski, A.R.; Smith, V.H.; Wang, S.H.; Martin, M.C.; DeNoyelles, F. Effects of non-algal turbidity on cyanobacterial biomass in seven turbid Kansas reservoirs. Lake Reserv. Manag. 2011, 27, 6–14. [Google Scholar] [CrossRef]
- Knowlton, M.F.; Jones, J.R. Non-algal seston, light, nutrients and chlorophyll in missouri reservoirs. Lake Reserv. Manag. 2000, 16, 322–332. [Google Scholar] [CrossRef]
- Huo, S.; Ma, C.; Xi, B.; Gao, R.; Deng, X.; Jiang, T.; He, Z.; Su, J.; Wu, F.; Liu, H. Lake ecoregions and nutrient criteria development in China. Ecol. Indic. 2014, 46, 1–10. [Google Scholar] [CrossRef]
- Yan, Z.; Han, W.; Peñuelas, J.; Sardans, J.; Elser, J.J.; Du, E.; Reich, P.B.; Fang, J. Phosphorus accumulates faster than nitrogen globally in freshwater ecosystems under anthropogenic impacts. Ecol. Lett. 2016, 19, 1237–1246. [Google Scholar] [CrossRef] [PubMed]
- Mamun, M.; Kim, J.Y.; An, K.G. Trophic responses of the Asian reservoir to long-term seasonal and interannual dynamic monsoon. Water 2020, 12, 2066. [Google Scholar] [CrossRef]
- Nürnberg, G.K. Trophic state of clear and colored, soft- and hardwater lakes with special consideration of nutrients, anoxia, phytoplankton and fish. Lake Reserv. Manag. 1996, 12, 432–447. [Google Scholar] [CrossRef]
- Zou, W.; Zhu, G.; Cai, Y.; Vilmi, A.; Xu, H.; Zhu, M.; Gong, Z.; Zhang, Y.; Qin, B. Relationships between nutrient, chlorophyll a and Secchi depth in lakes of the Chinese Eastern Plains ecoregion: Implications for eutrophication management. J. Environ. Manag. 2020, 260, 109923. [Google Scholar] [CrossRef]
- Duan, W.; He, B.; Chen, Y.; Zou, S.; Wang, Y.; Nover, D.; Chen, W.; Yang, G. Identification of long-term trends and seasonality in high-frequency water quality data from the Yangtze River basin, China. PLoS ONE 2018, 13, e0188889. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Elhadj, A.; Xu, H.; Xu, X.; Qiao, Z. A study on the relationship between land use change and water quality of the Mitidja watershed in Algeria based on GIS and RS. Sustainability 2020, 12, 3510. [Google Scholar] [CrossRef]
- WHO. Guidelines for Drinking Water Quality: Management of Cyanobacteria in Drinking Water Suppliers Information for Regulators and Water Suppliers, 4th ed.; WHO: Geneva, Switzerland, 2015. [Google Scholar]
- Miller, A.W. Trophic state evaluation for selected lakes in Yellowstone National Park, USA. WIT Trans. Ecol. Environ. 2010, 135, 143–155. [Google Scholar] [CrossRef]
- Carlson, R.E. Expanding the Trophic State Concept to Identify Non-Nutrient Limited Lakes and Reservoirs. In Enhancing the States’s Lake Management Programs; North American Lake Management Society: Chicago, IL, USA, 1991; pp. 59–71. [Google Scholar]
- Carlson, R. Estimating Trophic State. Lakeline 2007, 27, 25–28. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reservoir Types | Summary Attributes | WT | EC | DO | TSS | COD | TOC | TN | TP | TN:TP | CHL-a | CHL-a:TP | SD | NAT |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HLR (n = 19) | Min | 4.70 | 40.00 | 6.70 | 0.80 | 1.80 | 1.20 | 0.52 | 6.58 | 39.62 | 1.37 | 0.14 | 0.90 | −0.01 |
| Max | 17.50 | 312.88 | 14.30 | 9.86 | 8.20 | 5.90 | 2.55 | 25.06 | 229.43 | 12.10 | 0.89 | 4.50 | 1.18 | |
| Mean | 9.06 | 103.85 | 10.02 | 2.73 | 3.86 | 2.40 | 1.45 | 14.41 | 119.44 | 5.81 | 0.44 | 2.48 | 0.31 | |
| SE | 0.82 | 15.25 | 0.50 | 0.55 | 0.39 | 0.29 | 0.13 | 1.23 | 12.73 | 0.68 | 0.04 | 0.25 | 0.07 | |
| MLR (n = 64) | Min | 5.00 | 26.50 | 6.15 | 0.60 | 1.98 | 1.30 | 0.33 | 5.00 | 11.26 | 0.65 | 0.03 | 0.70 | −0.32 |
| Max | 18.80 | 333.00 | 13.10 | 17.50 | 12.53 | 9.23 | 2.51 | 39.33 | 362.40 | 23.00 | 1.89 | 10.00 | 1.06 | |
| Mean | 10.43 | 112.84 | 9.54 | 3.49 | 4.57 | 2.89 | 1.25 | 16.84 | 97.51 | 6.71 | 0.47 | 2.55 | 0.31 | |
| SE | 0.41 | 7.76 | 0.21 | 0.39 | 0.26 | 0.18 | 0.07 | 1.09 | 7.75 | 0.50 | 0.04 | 0.18 | 0.03 | |
| LLR (n = 49) | Min | 2.50 | 27.75 | 4.63 | 1.00 | 1.94 | 1.18 | 0.49 | 8.00 | 15.40 | 1.24 | 0.05 | 0.30 | 0.04 |
| Max | 18.63 | 414.20 | 13.45 | 20.57 | 9.13 | 6.37 | 2.29 | 44.33 | 200.75 | 22.67 | 1.27 | 3.50 | 3.13 | |
| Mean | 11.22 | 128.65 | 9.70 | 4.26 | 4.90 | 2.96 | 1.12 | 20.32 | 68.55 | 8.24 | 0.47 | 1.91 | 0.47 | |
| SE | 0.56 | 12.68 | 0.25 | 0.49 | 0.23 | 0.16 | 0.07 | 1.21 | 5.56 | 0.73 | 0.04 | 0.11 | 0.09 | |
| VLLR (n = 72) | Min | 5.70 | 36.00 | 4.83 | 1.03 | 1.60 | 1.10 | 0.31 | 5.00 | 5.93 | 2.38 | 0.06 | 0.50 | −0.91 |
| Max | 24.30 | 54,320.08 | 15.02 | 34.68 | 20.00 | 9.61 | 5.64 | 126.20 | 334.96 | 167.48 | 1.88 | 5.11 | 1.43 | |
| Mean | 13.44 | 2913.95 | 10.14 | 7.98 | 7.05 | 4.12 | 1.33 | 39.10 | 57.21 | 19.26 | 0.51 | 1.61 | 0.43 | |
| SE | 0.51 | 1126.26 | 0.23 | 0.78 | 0.44 | 0.23 | 0.13 | 3.65 | 6.81 | 3.04 | 0.04 | 0.11 | 0.05 |
| Reservoir Types | Models | R-Value | R2 | p-Value |
|---|---|---|---|---|
| HLR | Log10 (SD) = 1.35 − 0.89 × Log10 (TP) | −0.72 | 0.53 | <0.01 |
| Log10 (SD) = 0.37 − 0.20 × Log10 (TN) | −0.17 | 0.02 | <0.48 | |
| Log10 (SD) = 0.51 − 0.51 × Log10 (TSS) | −0.77 | 0.60 | <0.01 | |
| Log10 (SD) = 0.78 − 0.6 × Log10 (CHL-a) | −0.70 | 0.48 | <0.01 | |
| MLR | Log10 (SD) = 1.02 − 0.57 × Log10 (TP) | −0.56 | 0.31 | <0.01 |
| Log10 (SD) = 0.56 − 0.49 × Log10 (TSS) | −0.72 | 0.52 | <0.01 | |
| Log10 (SD) = 0.67 − 0.43 × Log10 (CHL-a) | −0.60 | 0.36 | <0.01 | |
| LLR | Log10 (SD) = 0.94 − 0.54 × Log10 (TP) | −0.49 | 0.24 | <0.01 |
| Log10 (SD) = 0.44 − 0.37 × Log10 (TSS) | −0.52 | 0.28 | <0.01 | |
| Log10 (SD) = 0.34 − 0.12 × Log10 (CHL-a) | −0.17 | 0.03 | <0.22 | |
| VLLR | Log10 (SD) = 0.62 − 0.33 × Log10 (TP) | −0.48 | 0.23 | <0.01 |
| Log10 (SD) = 0.14 − 0.12 × Log10 (TN) | −0.14 | 0.02 | <0.23 | |
| Log10 (SD) = 0.45 − 0.41 × Log10 (TSS) | −0.62 | 0.38 | <0.01 | |
| Log10 (SD) = 0.45 − 0.29 × Log10 (CHL-a) | −0.48 | 0.24 | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mamun, M.; Atique, U.; An, K.-G. Assessment of Water Quality Based on Trophic Status and Nutrients-Chlorophyll Empirical Models of Different Elevation Reservoirs. Water 2021, 13, 3640. https://doi.org/10.3390/w13243640
Mamun M, Atique U, An K-G. Assessment of Water Quality Based on Trophic Status and Nutrients-Chlorophyll Empirical Models of Different Elevation Reservoirs. Water. 2021; 13(24):3640. https://doi.org/10.3390/w13243640
Chicago/Turabian StyleMamun, Md, Usman Atique, and Kwang-Guk An. 2021. "Assessment of Water Quality Based on Trophic Status and Nutrients-Chlorophyll Empirical Models of Different Elevation Reservoirs" Water 13, no. 24: 3640. https://doi.org/10.3390/w13243640
APA StyleMamun, M., Atique, U., & An, K.-G. (2021). Assessment of Water Quality Based on Trophic Status and Nutrients-Chlorophyll Empirical Models of Different Elevation Reservoirs. Water, 13(24), 3640. https://doi.org/10.3390/w13243640