Trophic Magnification of Legacy (PCB, DDT and Hg) and Emerging Pollutants (PFAS) in the Fish Community of a Small Protected Southern Alpine Lake (Lake Mergozzo, Northern Italy)


 ,
,  ,
,  , and
, and 
Abstract
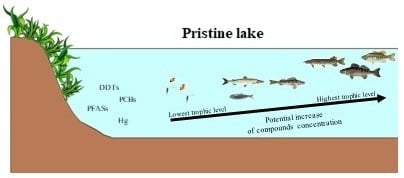
1. Introduction
2. Materials and Methods
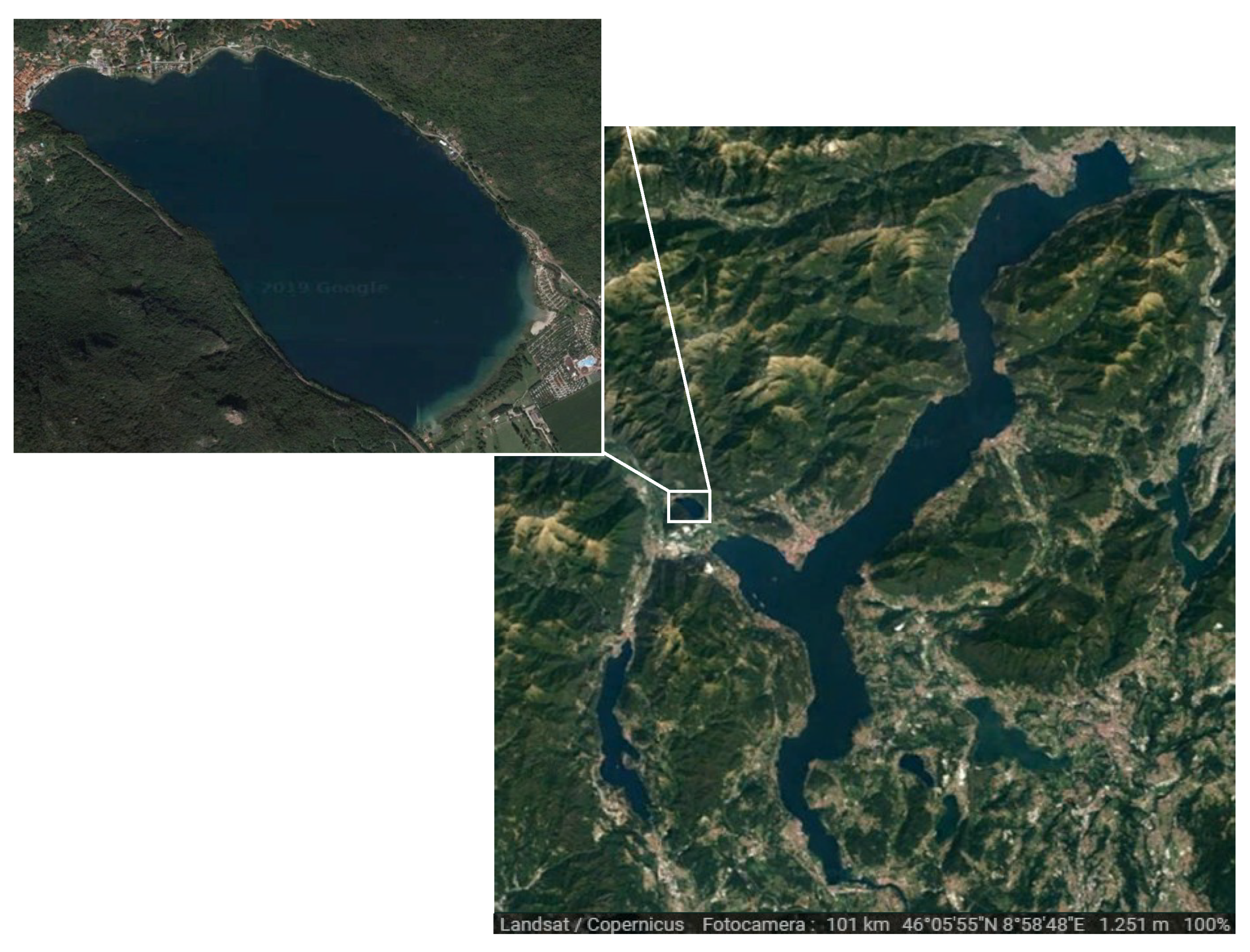
2.1. Study Area
2.2. Zooplankton, Benthos and Fish Sampling
2.3. Stable Isotopes Analysis (SIA)
2.4. Analysis of Contaminants
2.5. Determination of Trophic Levels and Trophic Magnification Factors
3. Results and Discussion
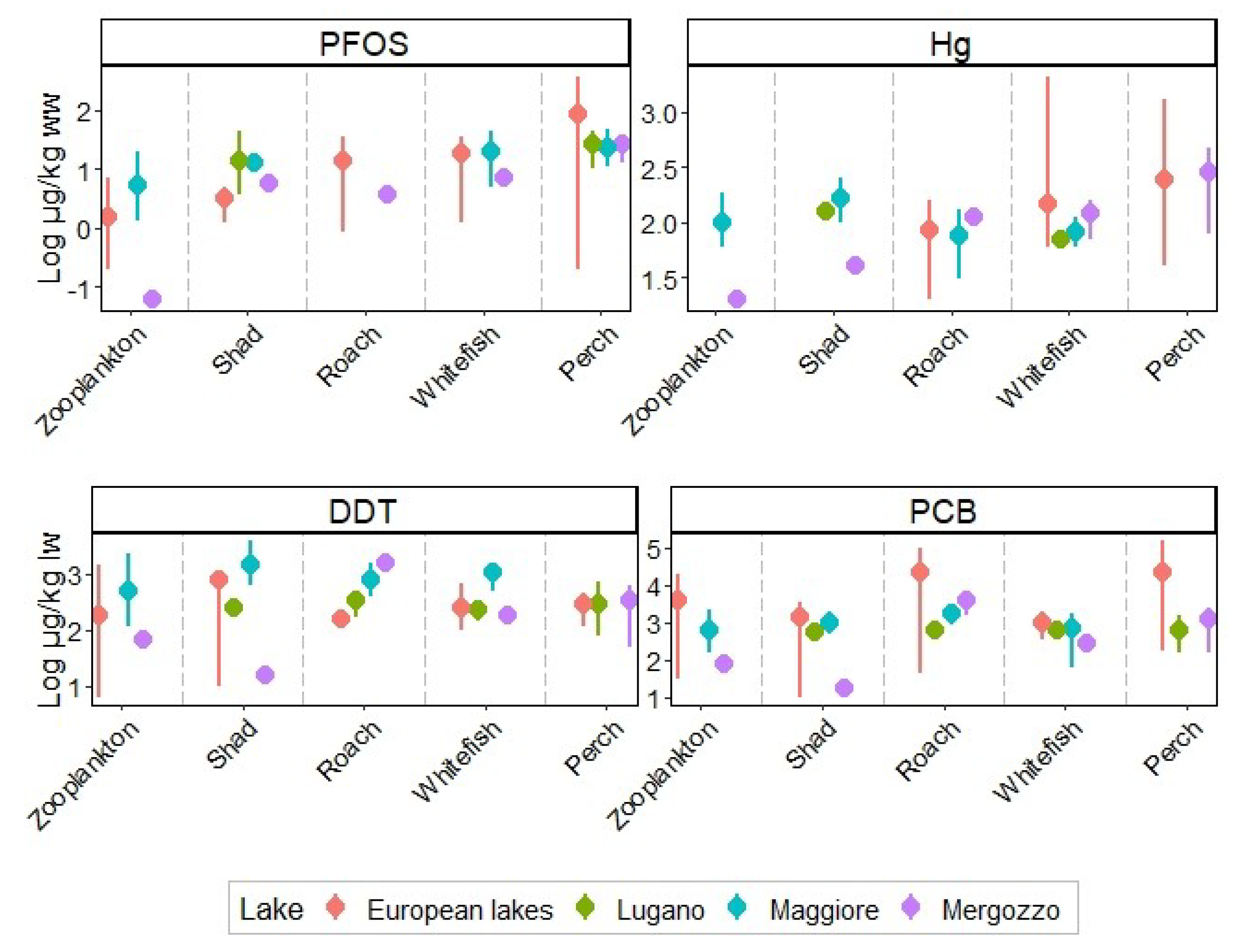
3.1. Chemicals Concentrations in Biota of Lake Mergozzo
3.2. Trophic Levels
3.3. Trophic Magnification Factors
3.4. Adjusting Monitoring Data for the Trophic Level for Comparison with the Established EQSbiota
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mackay, D.; Fraser, A. Bioaccumulation of persistent organic chemicals: Mechanisms and models. Environ. Pollut. 2000, 110, 375–391. [Google Scholar] [CrossRef]
- Conder, J.M.; Gobas, F.A.P.C.; Borgå, K.; Muir, D.C.G.; Powell, D.E. Use of trophic magnification factors and related measures to characterize bioaccumulation potential of chemicals. Integr. Environ. Assess. Manag. 2012, 8, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Borgå, K.; Kidd, K.A.; Muir, D.C.G.; Berglund, O.; Conder, J.M.; Gobas, F.A.P.C.; Kucklick, J.; Malm, O.; Powell, D.E. Trophic magnification factors: Considerations of ecology, ecosystems, and study design. Integr. Environ. Assess. Manag. 2012, 8, 64–84. [Google Scholar] [CrossRef] [PubMed]
- Gobas, F.A.P.C.; de Wolf, W.; Burkhard, L.P.; Verbruggen, E.; Plotzke, K. Revisiting bioaccumulation criteria for POPs and PBT assessments. Integr. Environ. Assess. Manag. 2009, 5, 624–637. [Google Scholar] [CrossRef] [PubMed]
- Franklin, J. How reliable are field-derived biomagnification factors and trophic magnification factors as indicators of bioaccumulation potential? Conclusions from a case study on per- and polyfluoroalkyl substances. Integr. Environ. Assess. Manag. 2016, 12, 6–20. [Google Scholar] [CrossRef] [PubMed]
- Won, E.J.; Choi, B.; Hong, S.; Khim, J.S.; Shin, K.H. Importance of accurate trophic level determination by nitrogen isotope of amino acids for trophic magnification studies: A review. Environ. Pollut. 2018, 238, 677–690. [Google Scholar] [CrossRef]
- Jardine, T.D.; Kidd, K.A.; Fisk, A.T. Applications, considerations, and sources of uncertainty when using stable isotope analysis in ecotoxicology. Environ. Sci. Technol. 2006, 40, 7501–7511. [Google Scholar] [CrossRef]
- Post, D.M. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar]
- Gioia, R.; Akindele, A.J.; Adebusoye, S.A.; Asante, K.A.; Tanabe, S.; Buekens, A.; Sasco, A.J. Polychlorinated biphenyls (PCBs) in Africa: A review of environmental levels. Environ. Sci. Pollut. Res. 2014, 21, 6278–6289. [Google Scholar] [CrossRef]
- Jones, K.C.; de Voogt, P. Persistent organic pollutants (POPs): State of the science. Environ. Pollut. 1999, 100, 209–221. [Google Scholar] [CrossRef]
- Ribeiro, E.; Ladeira, C.; Viegas, S. EDCs mixtures: A stealthy hazard for human health? Toxics 2017, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Ruzzin, J. Public health concern behind the exposure to persistent organic pollutants and the risk of metabolic diseases. BMC Public Health 2012, 12, 1. [Google Scholar] [CrossRef] [PubMed]
- Sijm, D.T.H.M.; Rikken, M.G.J.; Rorije, E.; Traas, T.P.; McLachan, M.S.; Peijnenburg, W.J.G.M. Transport, accumulation and transformation processes. In Risk Assessment of Chemicals: An Introduction; Leeuwen, C.J., van Vermeire, T.G., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 73–158. ISBN 978-1-4020-6102-8. [Google Scholar]
- Mitra, S.; Corsolini, S.; Pozo, K.; Audy, O.; Sarkar, S.K.; Biswas, J.K. Characterization, source identification and risk associated with polyaromatic and chlorinated organic contaminants (PAHs, PCBs, PCBs and OCPs) in the surface sediments of Hooghly estuary, India. Chemosphere 2019, 221, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Olatunji, O.S. Evaluation of selected polychlorinated biphenyls (PCBs) congeners and dichlorodiphenyltrichloroethane (DDT) in fresh root and leafy vegetables using GC-MS. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Yang, J.; Qadeer, A.; Liu, M.; Zhu, J.M.; Huang, Y.P.; Du, W.N.; Wei, X.Y. Occurrence, source, and partition of PAHs, PCBs, and OCPs in the multiphase system of an urban lake, Shanghai. Appl. Geochem. 2019, 106, 17–25. [Google Scholar] [CrossRef]
- Atmaca, E.; Das, Y.K.; Yavuz, O.; Aksoy, A. An evaluation of the levels of organochlorine compounds (OCPs and PCBs) in cultured freshwater and wild sea fish eggs as an exposure biomarker for environmental contamination. Environ. Sci. Pollut. Res. 2019, 26, 7005–7012. [Google Scholar] [CrossRef]
- Rigét, F.; Bignert, A.; Braune, B.; Dam, M.; Dietz, R.; Evans, M.; Green, N.; Gunnlaugsdóttir, H.; Hoydal, K.S.; Kucklick, J.; et al. Temporal trends of persistent organic pollutants in Arctic marine and freshwater biota. Sci. Total Environ. 2019, 649, 99–110. [Google Scholar] [CrossRef]
- Robinson, T.; Ali, U.; Mahmood, A.; Chaudhry, M.J.I.; Li, J.; Zhang, G.; Jones, K.C.; Malik, R.N. Concentrations and patterns of organochlorines (OCs) in various fish species from the Indus River, Pakistan: A human health risk assessment. Sci. Total Environ. 2016, 541, 1232–1242. [Google Scholar] [CrossRef]
- Corsolini, S.; Sarà, G. The trophic transfer of persistent pollutants (HCB, DDTs, PCBs) within polar marine food webs. Chemosphere 2019, 177, 189–199. [Google Scholar] [CrossRef]
- Zhang, H.; Teng, Y.; Doan, T.T.T.; Yat, Y.W.; Chan, S.H.; Kelly, B.C. Stable nitrogen and carbon isotopes in sediments and biota from three tropical marine food webs: Application to chemical bioaccumulation assessment. Environ. Toxicol. Chem. 2017, 36, 2521–2532. [Google Scholar] [CrossRef]
- Helou, K.; Harmouche-karaki, M.; Karake, S.; Narbonne, J. A review of organochlorine pesticides and polychlorinated biphenyls in Lebanon: Environmental and human contaminants. Chemosphere 2019, 231, 357–368. [Google Scholar] [CrossRef] [PubMed]
- UNEP Stockholm convention on persistent organic pollutants (POPs). Geneva, 2001. Available online: http://chm.pops.int/Portals/0/Repository/convention_text/UNEP-POPS-COP-CONVTEXT-FULL.English.PDF (accessed on 3 June 2020).
- Driscoll, C.T.; Mason, R.P.; Chan, H.M.; Jacob, D.J.; Pirrone, N. Mercury as a global pollutant: Sources, pathways, and effects. Environ. Sci. Technol. 2013, 47, 4967–4983. [Google Scholar] [CrossRef]
- Streets, D.G.; Horowitz, H.M.; Lu, Z.; Levin, L.; Thackray, C.P.; Sunderland, E.M. Five hundred years of anthropogenic mercury: Spatial and temporal release profiles. Environ. Res. Lett. 2019, 14, 084004. [Google Scholar] [CrossRef]
- Azad, A.M.; Frantzen, S.; Bank, M.S.; Johnsen, I.A.; Tessier, E.; Amouroux, D.; Madsen, L.; Maage, A. Spatial distribution of mercury in seawater, sediment, and seafood from the Hardangerfjord ecosystem, Norway. Sci. Total Environ. 2019, 667, 622–637. [Google Scholar] [CrossRef] [PubMed]
- Vardè, M.; Servidio, A.; Vespasiano, G.; Pasti, L.; Cavazzini, A.; Di Traglia, M.; Rosselli, A.; Cofone, F.; Apollaro, C.; Cairns, W.R.L.; et al. Ultra-trace determination of total mercury in Italian bottled waters. Chemosphere 2019, 219, 896–913. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.L.; Der Fang, M.; Chien, L.C.; Lin, C.C.; Hsi, H.C. Distribution of mercury and methylmercury in surface water and surface sediment of river, irrigation canal, reservoir, and wetland in Taiwan. Environ. Sci. Pollut. Res. 2019, 26, 17762–17773. [Google Scholar] [CrossRef] [PubMed]
- Bargagli, R. Moss and lichen biomonitoring of atmospheric mercury: A review. Sci. Total Environ. 2016, 572, 216–231. [Google Scholar] [CrossRef]
- Lin, Y.; Vogt, R.; Larssen, T. Environmental mercury in China: A review. Environ. Toxicol. Chem. 2012, 31, 431–2444. [Google Scholar] [CrossRef]
- Rigét, F.; Braune, B.; Bignert, A.; Wilson, S.; Aars, J.; Born, E.; Dam, M.; Dietz, R.; Evans, M.; Evans, T.; et al. Temporal trends of Hg in Arctic biota, an update. Sci. Total Environ. 2011, 409, 3520–3526. [Google Scholar] [CrossRef]
- Lavoie, R.A.; Jardine, T.D.; Chumchal, M.M.; Kidd, K.A.; Campbell, L.M. Biomagnification of mercury in aquatic food webs: A worldwide meta-analysis. Environ. Sci. Technol. 2013, 47, 13385–13394. [Google Scholar] [CrossRef]
- Rice, K.M.; Walker, E.M.; Wu, M.; Gillette, C.; Blough, E.R. Environmental mercury and its toxic effects. J. Prev. Med. Public Health 2014, 47, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Gantner, N.; Power, M.; Iqaluk, D.; Meili, M.; Borg, H.; Sundbom, M.; Solomon, K.R.; Lawson, G.; Muir, D.C.G. Mercury concentrations in landlocked arctic char (Salvelinus alpinus) from the Canadian arctic. Part I: Insights from trophic relationships in 18 lakes. Environ. Toxicol. Chem. 2010, 29, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Kidd, K.A.; Bootsma, H.A.; Hesslein, R.H.; Muir, D.C.G.; Hecky, R.E. Biomagnification of DDT through the benthic and pelagic food webs of Lake Malawi, East Africa: Importance of trophic level and carbon source. Environ. Sci. Technol. 2001, 35, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Houde, M.; Muir, D.C.G.; Kidd, K.A.; Guildford, S.; Drouillard, K.; Evans, M.S.; Wang, X.; Whittle, D.M.; Haffner, D.; Kling, H. Influence of lake characteristics on the biomagnification of persistent organic pollutants in lake trout food webs. Environ. Toxicol. Chem. 2008, 27, 2169–2178. [Google Scholar] [CrossRef]
- Kelly, B.C.; Ikonomou, M.G.; Blair, J.D.; Surridge, B.; Hoover, D.; Grace, R.; Gobas, F.A.P.C. Perfluoroalkyl contaminants in an arctic marine food web: Trophic magnification and wildlife exposure. Environ. Sci. Technol. 2009, 43, 4037–4043. [Google Scholar] [CrossRef]
- Ahrens, L.; Bundschuh, M. Fate and effects of poly- and perfluoroalkyl substances in the aquatic environment: A review. Environ. Toxicol. Chem. 2014, 33, 1921–1929. [Google Scholar] [CrossRef]
- Lau, C.; Anitole, K.; Hodes, C.; Lai, D.; Pfahles-Hutchens, A.; Seed, J. Perfluoroalkyl acids: A review of monitoring and toxicological findings. Toxicol. Sci. 2007, 99, 366–394. [Google Scholar] [CrossRef]
- Houde, M.; De Silva, A.O.; Muir, D.C.G.; Letcher, R.J. Monitoring of Perfluorinated Compounds in Aquatic Biota: An Updated Review. Environ. Sci. Technol. 2011, 45, 7962–7973. [Google Scholar] [CrossRef]
- Kidd, K.A.; Burkhard, L.P.; Babut, M.; Borgå, K.; Muir, D.C.G.; Perceval, O.; Ruedel, H.; Woodburn, K.; Embry, M.R. Practical advice for selecting or determining trophic magnification factors for application under the European Union Water Framework Directive. Integr. Environ. Assess. Manag. 2019, 15, 266–277. [Google Scholar] [CrossRef]
- Volta, P.; Jeppesen, E.; Campi, B.; Sala, P.; Emmrich, M.; Winfield, I.J. The population biology and life history traits of Eurasian ruffe [Gymnocephalus cernuus (L.), Pisces: Percidae] introduced into eutrophic and oligotrophic lakes in Northern Italy. J. Limnol. 2013, 72, 280–290. [Google Scholar] [CrossRef]
- Volta, P.; Yan, N.D.; Gunn, J.M. Past, present and future of the fish community of Lake Orta (Italy), one of the world’s largest acidified lakes. J. Limnol. 2016, 75, 131–141. [Google Scholar] [CrossRef]
- ARPA Piemonte Attività Arpa nella gestione delle reti di monitoraggio regionale delle acque superficiali—Corsi d’ acqua e laghi. Relazione monitoraggio anno 2016. 2017. Available online: http://www.arpa.piemonte.it/approfondimenti/temi-ambientali/acqua/acque-superficiali-corsi-dacqua/documentazione-e-dati/relazioni-tecniche-fiumi-e-laghi-2016 (accessed on 3 June 2020).
- Volta, P.; Jeppesen, E.; Sala, P.; Galafassi, S.; Foglini, C.; Puzzi, C.; Winfield, I.J. Fish assemblages in deep Italian subalpine lakes: History and present status with an emphasis on non-native species. Hydrobiologia 2018, 824, 255–270. [Google Scholar] [CrossRef]
- Visconti, A.; Caroni, R.; Rawcliffe, R.; Fadda, A.; Piscia, R.; Manca, M. Defining seasonal functional traits of a freshwater zooplankton community using δ13C and δ15N stable isotope analysis. Water (Switzerland) 2018, 10, 108. [Google Scholar] [CrossRef]
- Visconti, A.; Manca, M. Seasonal changes in the δ13C and δ15N signatures of the Lago Maggiore pelagic food web. J. Limnol. 2011, 70, 263–271. [Google Scholar] [CrossRef]
- ISPRA. Linea guida per il monitoraggio delle sostanze prioritarie (secondo D.Lgs. 172/2015); ISPRA Manuali e linee guida 143/2016; ISPRA: Roma, Italy, 2016; ISBN 978-88-448-0795-5.
- Hitchcock, D.J.; Andersen, T.; Varpe, Ø.; Loonen, M.J.J.E.; Warner, N.A.; Herzke, D.; Tombre, I.M.; Griffin, L.R.; Shimmings, P.; Borgå, K. Potential Effect of Migration Strategy on Pollutant Occurrence in Eggs of Arctic Breeding Barnacle Geese (Branta leucopsis). Environ. Sci. Technol. 2019, 53, 5427–5435. [Google Scholar] [CrossRef] [PubMed]
- Bettinetti, R.; Quadroni, S.; Manca, M.; Piscia, R.; Volta, P.; Guzzella, L.; Roscioli, C.; Galassi, S. Seasonal fluctuations of DDTs and PCBs in zooplankton and fish of Lake Maggiore (Northern Italy). Chemosphere 2012, 88, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Mazzoni, M.; Polesello, S.; Rusconi, M.; Valsecchi, S. Liquid chromatography mass spectrometry determination of perfluoroalkyl acids in environmental solid extracts after phospholipid removal and on-line turbulent flow chromatography purification. J. Chromatogr. A 2016, 1453, 62–70. [Google Scholar] [CrossRef]
- McConnaughey, T.; McRoy, C.P. Food-Web structure and the fractionation of Carbon isotopes in the Bering sea. Mar. Biol. 1979, 53, 257–262. [Google Scholar] [CrossRef]
- Post, D.M.; Layman, C.A.; Arrington, D.A.; Takimoto, G.; Quattrochi, J.; Montaña, C.G. Getting to the fat of the matter: Models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia 2007, 152, 179–189. [Google Scholar] [CrossRef]
- Fliedner, A.; Rüdel, H.; Lohmann, N.; Buchmeier, G.; Koschorreck, J. Biota monitoring under the Water Framework Directive: On tissue choice and fish species selection. Environ. Pollut. 2018, 235, 129–140. [Google Scholar] [CrossRef]
- European Union. CIS-WFD Guidance Document No. 32 On biota monitoring (the implementation of EQSbiota) under the Water Framework Directive Technical Report 2014-083; Office for Official Publications of the European Communities: Luxembourg, 2014; ISBN 9789279446344. [Google Scholar]
- Di Cesare, A.; Eckert, E.M.; Corno, G. Co-selection of antibiotic and heavy metal resistance in freshwater bacteria. J. Limnol. 2016, 75, 59–66. [Google Scholar] [CrossRef]
- Pisanello, F.; Marziali, L.; Rosignoli, F.; Poma, G.; Roscioli, C.; Pozzoni, F.; Guzzella, L. In situ bioavailability of DDT and Hg in sediments of the Toce River (Lake Maggiore basin, Northern Italy): Accumulation in benthic invertebrates and passive samplers. Environ. Sci. Pollut. Res. 2016, 23, 10542–10555. [Google Scholar] [CrossRef] [PubMed]
- Post, D.M.; Pace, M.L.; Halrston, N.G. Ecosystem size determines food-chain length in lakes. Nature 2000, 405, 1047–1049. [Google Scholar] [CrossRef] [PubMed]
- Vander Zanden, M.J.; Rasmussen, J.B. Variation in δ15N and δ13C trophic fractionation: Implications for aquatic food web studies. Limnol. Oceanogr. 2001, 46, 2061–2066. [Google Scholar] [CrossRef]
- Leoni, B. Zooplankton predators and preys: Body size and stable isotope to investigate the pelagic food web in a deep lake (Lake Iseo, Northern Italy). J. Limnol. 2017, 76, 85–93. [Google Scholar] [CrossRef]
- Perga, M.-E.; Gerdeaux, D. Seasonal variability in the δ13C and δ15N values of the zooplankton taxa in two alpine lakes. Acta Oecologica 2006, 30, 69–77. [Google Scholar] [CrossRef]
- Wyn, B.; Kidd, K.A.; Burgess, N.M.; Curry, R.A. Mercury biomagnification in the food webs of acidic lakes in Kejimkujik National Park and National Historic Site, Nova Scotia. Can. J. Fish. Aquat. Sci. 2009, 66, 1532–1545. [Google Scholar] [CrossRef]
- Walters, D.M.; Jardine, T.D.; Cade, B.S.; Kidd, K.A.; Muir, D.C.G.; Leipzig-Scott, P. Trophic Magnification of Organic Chemicals: A Global Synthesis. Environ. Sci. Technol. 2016, 50, 4650–4658. [Google Scholar] [CrossRef]
- Åkerblom, S.; Negm, N.; Wu, P.; Bishop, K.; Ahrens, L. Variation and accumulation patterns of poly- and perfluoroalkyl substances (PFAS) in European perch (Perca fluviatilis) across a gradient of pristine Swedish lakes. Sci. Total Environ. 2017, 599-600, 1685–1692. [Google Scholar] [CrossRef]
- Lescord, G.L.; Kidd, K.A.; De Silva, A.O.; Williamson, M.; Spencer, C.; Wang, X.; Muir, D.C.G. Perfluorinated and polyfluorinated compounds in lake food webs from the Canadian High Arctic. Environ. Sci. Technol. 2015, 49, 2694–2702. [Google Scholar] [CrossRef]
- Zhou, Z.; Shi, Y.; Li, W.; Xu, L.; Cai, Y. Perfluorinated compounds in surface water and organisms from Baiyangdian Lake in North China: Source profiles, bioaccumulation and potential risk. Bull. Environ. Contam. Toxicol. 2012, 89, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Chen, X.; Zhao, S.; Zhang, Y.; Jiang, W.; Yang, L.; Zhu, L. Trophic magnification and isomer fractionation of perfluoroalkyl substances in the food web of Taihu Lake, China. Environ. Sci. Technol. 2014, 48, 2173–2182. [Google Scholar] [CrossRef] [PubMed]
- Munoz, G.; Budzinski, H.; Babut, M.; Drouineau, H.; Lauzent, M.; Le Menach, K.; Lobry, J.; Selleslagh, J.; Simonnet-Laprade, C.; Labadie, P. Evidence for the Trophic Transfer of Perfluoroalkylated Substances in a Temperate Macrotidal Estuary. Environ. Sci. Technol. 2017, 51, 8450–8459. [Google Scholar] [CrossRef] [PubMed]
- Ballutaud, M.; Drouineau, H.; Carassou, L.; Munoz, G.; Chevillot, X.; Labadie, P.; Budzinski, H.; Lobry, J. EStimating Contaminants tRansfers Over Complex food webs (ESCROC): An innovative Bayesian method for estimating POP’s biomagnification in aquatic food webs. Sci. Total Environ. 2019, 658, 638–649. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.A.; Hungerbühler, K. Bioconcentration of perfluorinated alkyl acids: How important is specific binding? Environ. Sci. Technol. 2013, 47, 7214–7223. [Google Scholar] [CrossRef] [PubMed]
- Armitage, J.M.; Arnot, J.A.; Wania, F.; Mackay, D. Development and evaluation of a mechanistic bioconcentration model for ionogenic organic chemicals in fish. Environ. Toxicol. Chem. 2013, 32, 115–128. [Google Scholar] [CrossRef]
- Simmonet-Laprade, C.; Budzinski, H.; Babut, M.; Le Menach, K.; Munoz, G.; Lauzent, M.; Ferrari, B.J.D.; Labadie, P. Investigation of the spatial variability of poly- and perfluoroalkyl substance trophic magnification in selected riverine ecosystems. Sci. Total Environ. 2019, 686, 393–401. [Google Scholar] [CrossRef]
- Gebbink, W.A.; Bignert, A.; Berger, U. Perfluoroalkyl Acids (PFAAs) and Selected Precursors in the Baltic Sea Environment: Do Precursors Play a Role in Food Web Accumulation of PFAAs? Environ. Sci. Technol. 2016, 50, 6354–6362. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Species Common Name | Species Scientific Name | Size | α | TL (SIA) | TL (Fishbase) |
|---|---|---|---|---|---|
| Zooplankton | >450 µm | 0.0 | 2.5 | ||
| Shad | Alosa agone | 0.3 | 4.1 | 3.0 | |
| Roach1 | Rutilus rutilus | small and medium | 0.4 | 4.4 | 3.0 |
| Chub | Squalius squalus | 0.3 | 4.5 | 2.7 | |
| Rudd1 | Scardinius hesperidicus | medium | 0.3 | 4.6 | 2.9 |
| Perch1 | Perca fluviatilis | medium | 0.5 | 4.7 | 4.4 |
| Pumpkinseed | Lepomis gibbosus | 0.5 | 4.7 | 3.3 | |
| Rudd2 | Scardinius hesperidicus | large | 0.4 | 4.7 | 2.9 |
| European whitefish1 | Coregonus lavaretus | medium | 0.2 | 4.8 | 3.1 |
| Roach2 | Rutilus rutilus | large | 0.4 | 4.8 | 3.0 |
| European whitefish2 | Coregonus lavaretus | large | 0.1 | 5.0 | 3.1 |
| Ruffe | Gymnocephalus cernuus | 0.3 | 5.0 | 3.3 | |
| Largemouth bass1 | Micropterus salmoides | large | 0.4 | 5.1 | 3.8 |
| Pikeperch | Sander lucioperca | 0.4 | 5.1 | 4.0 | |
| Largemouth bass2 | Micropterus salmoides | medium | 0.5 | 5.2 | 3.8 |
| Perch2 | Perca fluviatilis | large | 0.3 | 5.2 | 4.4 |
| Largemouth bass3 | Micropterus salmoides | medium | 0.4 | 5.3 | 3.8 |
| Pike | Esox cisalpinus | 0.4 | 5.3 | 4.1 | |
| Burbot | Lota lota | 0.0 | 5.5 | 3.8 | |
| Char | Salvelinus alpinus | 0.0 | 5.7 | 4.4 |
| Chemical | Slope | TMF | (95%) | R2 | p-Value |
|---|---|---|---|---|---|
| Hg (mg kg−1 dw) | 0.36 | 2.3 | (1.45; 3.55) | 0.42 | 0.001 |
| Hg (mg kg−1 ww) | 0.38 | 2.4 | (1.57; 3.61) | 0.49 | 0.000 |
| PCB 153 (µg kg−1 lw) | 1.00 | 10.0 | (3.55; 28.27) | 0.52 | 0.000 |
| Penta-CB (µg kg−1 lw) | 0.99 | 9.7 | (1.05; 88.76) | 0.16 | 0.045 |
| Hexa-CB (µg kg−1 lw) | 1.10 | 12.5 | (4.14; 37.67) | 0.54 | 0.000 |
| Hepta-CB (µg kg−1 lw) | 0.69 | 4.9 | (1.68; 14.36) | 0.32 | 0.006 |
| p,p′-DDD (µg kg−1 lw) | 1.15 | 14.1 | (4.78; 41.56) | 0.57 | 0.000 |
| p,p′-DDT tot (µg kg−1 lw) | 0.47 | 2.9 | (1.05; 8.13) | 0.17 | 0.042 |
| PFOS (µg kg−1 ww) | 0.48 | 3.0 | (1.17; 7.71) | 0.21 | 0.025 |
| PFDA (µg kg−1 ww) | 0.094 | ||||
| PFUnDA (µg kg−1 ww) | 0.631 | ||||
| PFDoDA (µg kg−1 ww) | 0.515 |
| Chemical | Slope | TMF | (95%) | R2 | p-Value |
|---|---|---|---|---|---|
| Hg (mg kg−1 dw) | 0.46 | 2.9 | (1.33; 6.13) | 0.29 | 0.01 |
| Hg (mg kg−1 ww) | 0.48 | 3.0 | (1.47; 6.06) | 0.35 | 0.00 |
| PCB 153 (µg kg−1 lw) | 0.10 | ||||
| Penta-CB (µg kg−1 lw) | 0.10 | ||||
| Hexa-CB (µg kg−1 lw) | 0.08 | ||||
| Hepta-CB (µg kg−1 lw) | 0.87 | 7.5 | (1.17; 47.24) | 0.19 | 0.03 |
| p,p′-DDD (µg kg−1 lw) | 0.10 | ||||
| p,p′-DDT tot (µg kg−1 lw) | 0.11 | ||||
| PFOS (µg kg−1 ww) | 0.75 | ||||
| PFDA (µg kg−1 ww) | 0.82 | ||||
| PFUnDA (µg kg−1 ww) | 0.83 | ||||
| PFDoDA (µg kg−1 ww) | 0.88 |
| Common Name | TL | PFOS Measured | PFOS TL-Adjusted | Hg Measured | Hg TL-Adjusted |
|---|---|---|---|---|---|
| µg kg−1 ww | µg kg−1 ww | µg kg−1 ww | µg kg−1 ww | ||
| Zooplankton | 2.5 | 0.1 | 0.3 | 20.0 | 73.2 |
| Shad | 4.1 | 5.7 | 5.1 | 35.9 | 32.8 |
| Roach1 | 4.4 | 3.4 | 2.2 | 111 | 78.2 |
| Chub | 4.5 | 2.4 | 1.5 | 54.9 | 36.7 |
| Rudd1 | 4.6 | 0.9 | 0.5 | 68.7 | 42.4 |
| Perch1 | 4.7 | 12.9 | 6.1 | 84.4 | 46.7 |
| Pumpkinseed | 4.7 | 3.7 | 1.7 | 73.0 | 39.6 |
| Rudd2 | 4.7 | 13.7 | 6.6 | 103 | 57.9 |
| European whitefish1 | 4.8 | 8.1 | 3.2 | 70.9 | 34.0 |
| Roach2 | 4.8 | 3.8 | 1.6 | 107 | 54.7 |
| European whitefish2 | 5.0 | 5.4 | 1.7 | 160 | 65.0 |
| Ruffe | 5.0 | 3.6 | 1.2 | 50.1 | 20.8 |
| Largemouth bass1 | 5.1 | 13.7 | 3.9 | 213 | 78.6 |
| Pikeperch | 5.1 | 13.3 | 3.9 | 501 | 191 |
| Largemouth bass2 | 5.2 | 6.2 | 1.7 | 143 | 50.1 |
| Perch2 | 5.2 | 38.4 | 10.0 | 472 | 163 |
| Largemouth bass3 | 5.3 | 13.8 | 3.2 | 222 | 70.3 |
| Northern pike | 5.3 | 2.9 | 0.7 | 188 | 61.6 |
| Burbot | 5.5 | 0.3 | 0.1 | 260 | 68.2 |
| Char | 5.7 | 1.7 | 0.3 | 87.6 | 19.8 |
| Min | 0.1 | 0.1 | 20.0 | 19.8 | |
| Max | 38.4 | 10.0 | 501 | 191 | |
| Geometric Mean | 3.9 | 1.6 | 111 | 54.6 | |
| EQS | 9.1 | 9.1 | 20 | 20 | |
| N samples | 20 | 20 | 20 | 20 | |
| N > EQS | 6 | 1 | 19 | 19 | |
| % Exceedance | 30 | 5 | 95 | 95 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazzoni, M.; Ferrario, C.; Bettinetti, R.; Piscia, R.; Cicala, D.; Volta, P.; Borgå, K.; Valsecchi, S.; Polesello, S. Trophic Magnification of Legacy (PCB, DDT and Hg) and Emerging Pollutants (PFAS) in the Fish Community of a Small Protected Southern Alpine Lake (Lake Mergozzo, Northern Italy). Water 2020, 12, 1591. https://doi.org/10.3390/w12061591
Mazzoni M, Ferrario C, Bettinetti R, Piscia R, Cicala D, Volta P, Borgå K, Valsecchi S, Polesello S. Trophic Magnification of Legacy (PCB, DDT and Hg) and Emerging Pollutants (PFAS) in the Fish Community of a Small Protected Southern Alpine Lake (Lake Mergozzo, Northern Italy). Water. 2020; 12(6):1591. https://doi.org/10.3390/w12061591
Chicago/Turabian StyleMazzoni, Michela, Claudia Ferrario, Roberta Bettinetti, Roberta Piscia, Davide Cicala, Pietro Volta, Katrine Borgå, Sara Valsecchi, and Stefano Polesello. 2020. "Trophic Magnification of Legacy (PCB, DDT and Hg) and Emerging Pollutants (PFAS) in the Fish Community of a Small Protected Southern Alpine Lake (Lake Mergozzo, Northern Italy)" Water 12, no. 6: 1591. https://doi.org/10.3390/w12061591
APA StyleMazzoni, M., Ferrario, C., Bettinetti, R., Piscia, R., Cicala, D., Volta, P., Borgå, K., Valsecchi, S., & Polesello, S. (2020). Trophic Magnification of Legacy (PCB, DDT and Hg) and Emerging Pollutants (PFAS) in the Fish Community of a Small Protected Southern Alpine Lake (Lake Mergozzo, Northern Italy). Water, 12(6), 1591. https://doi.org/10.3390/w12061591