What Triggers the Annual Cycle of Cyanobacterium Oscillatoria sp. in an Extreme Environmental Sulfide-Rich Spa?

 ,
,  ,
,  and
and
Abstract
1. Introduction
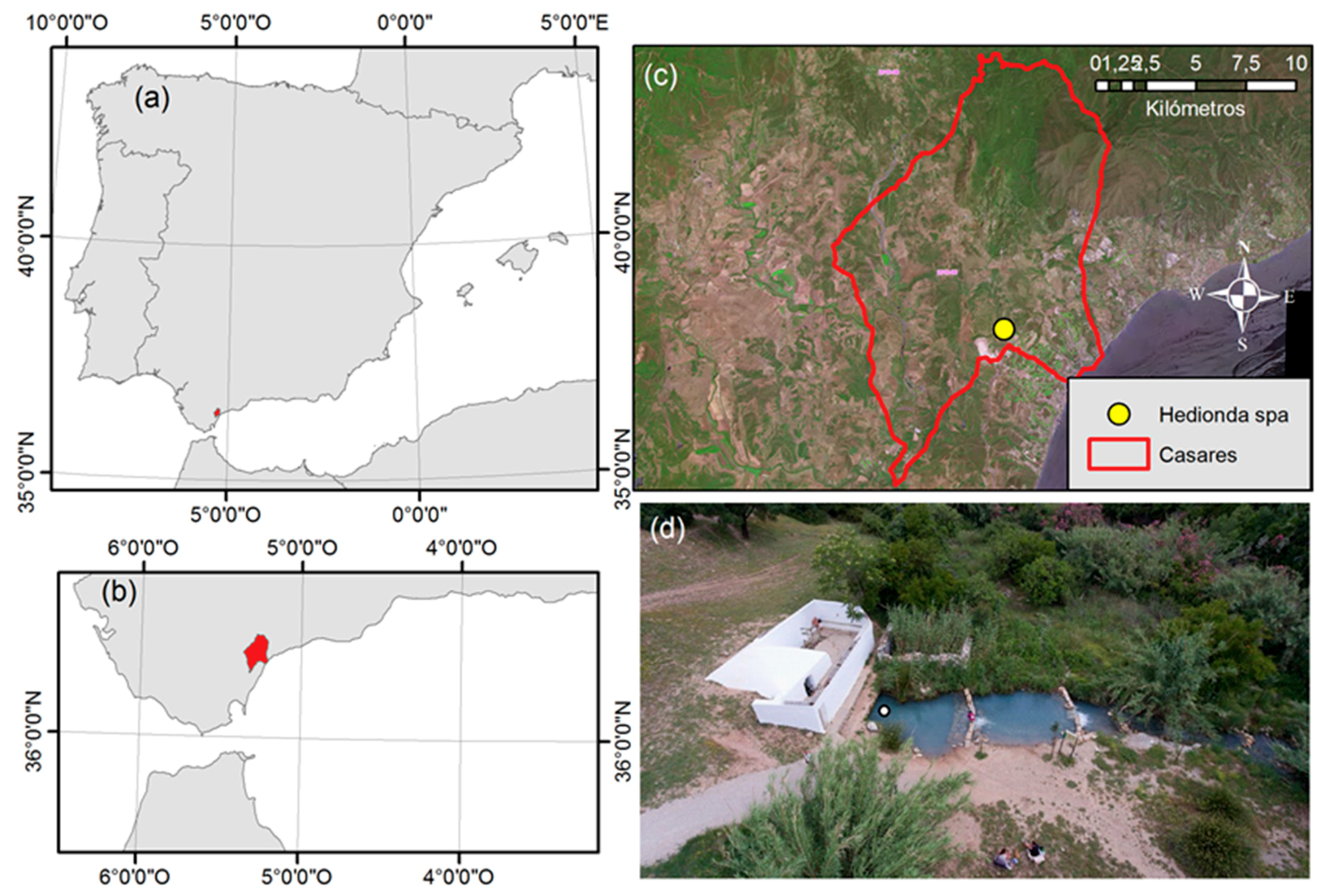
2. Materials and Methods
3. Results
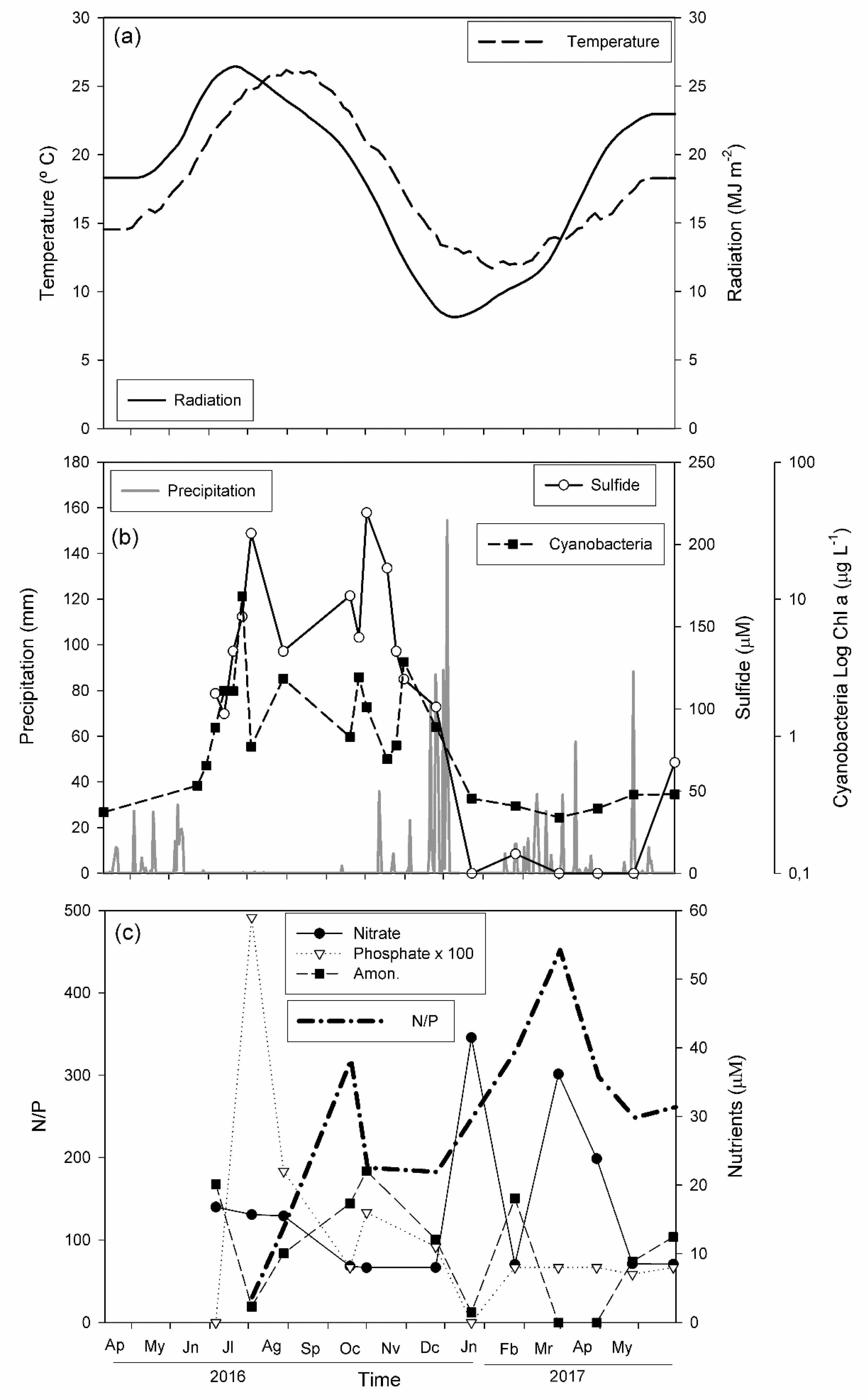
3.1. Abiotic Factors
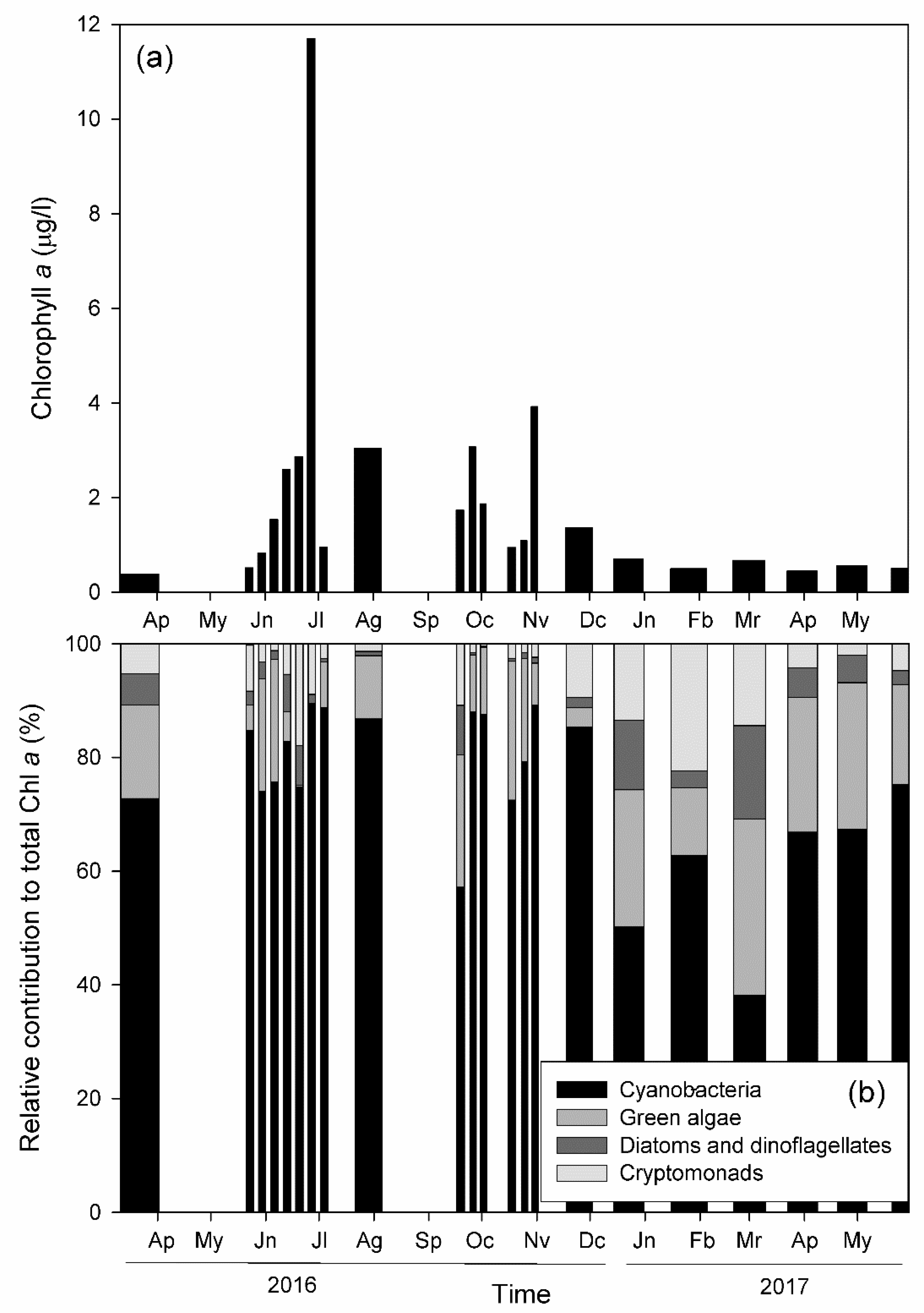
3.2. Phytoplankton Abundance and Diversity
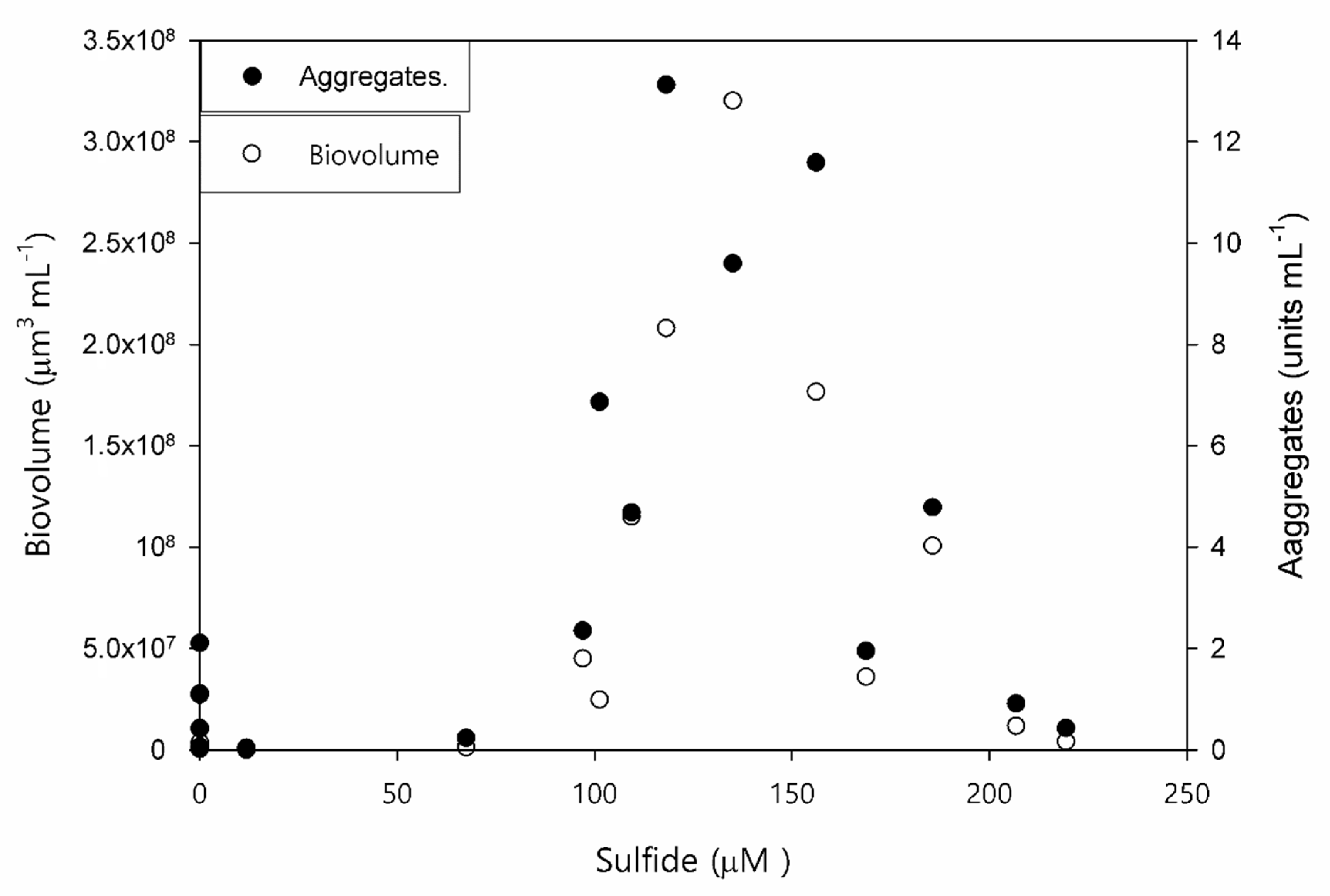
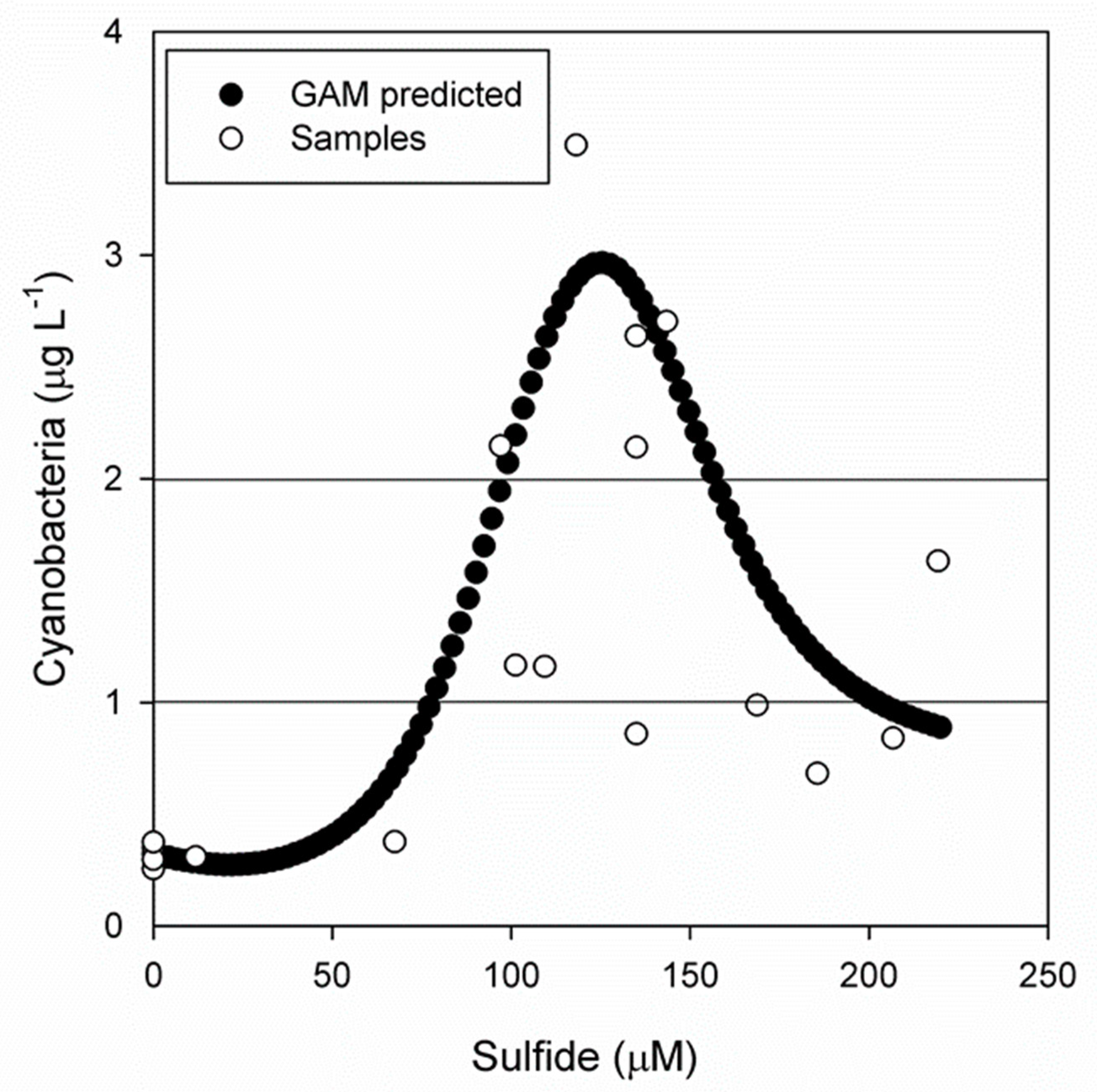
3.3. What Triggers Cyanobacteria (Oscillatoria sp.) Concentration?
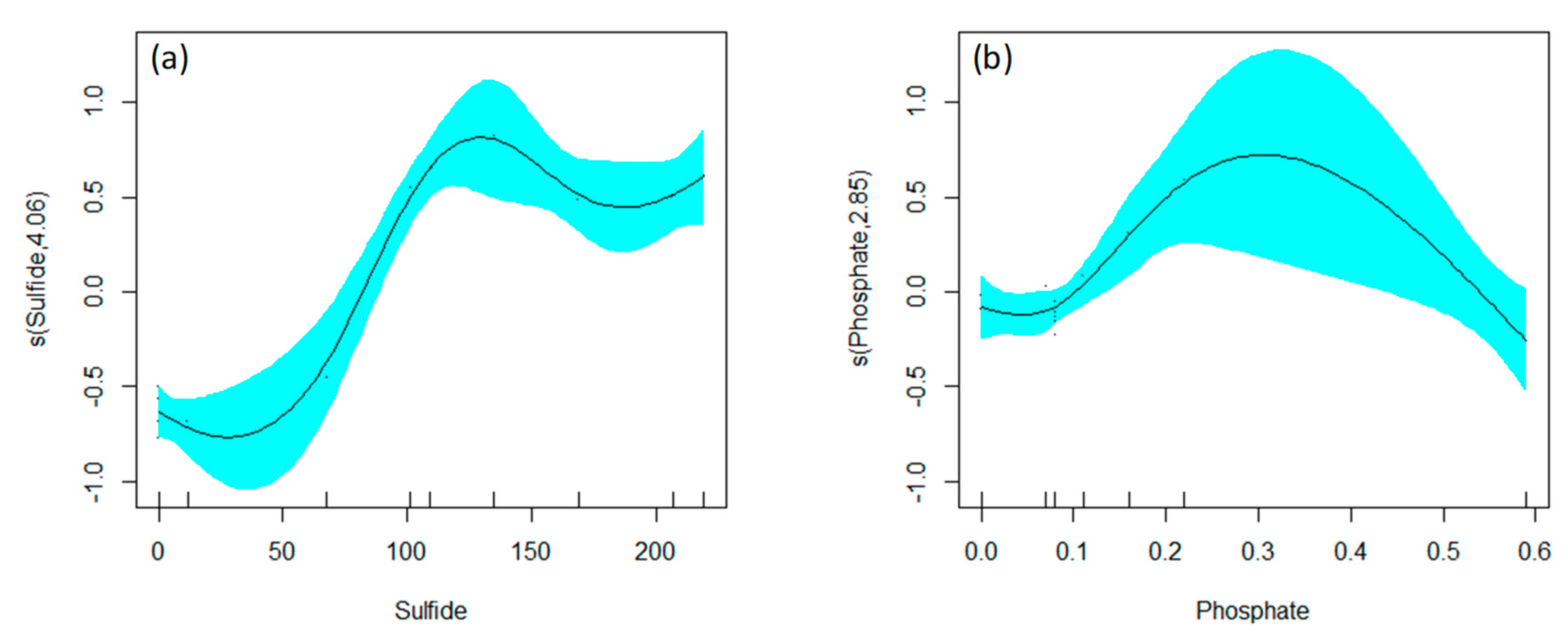
GAM Analysis
4. Discussion
5. Conclusions
- Total sulfide concentration in La Hedionda Spa shows an annual cycle, with concentrations around 40 μM in winter and up to 200 μM in summer.
- Regardless of cyanobacteria, other phytoplankton groups show consistently similar and low concentrations throughout the year.
- The fact that cyanobacteria (Oscillatoria sp.) reaches the highest concentration at high sulfide concentrations suggests the presence of a high sulfide-adapted Oscillatoria sp. population.
- In contrast to generally accepted succession models in epicontinental waters, neither nutrient nor light, but the selective agent (sulfide) positively triggers Oscillatoria sp. proliferation in summer.
- While Oscillatoria sp. are distributed worldwide, the present strain might be the result of an almost 2000 year adaptation with the annual sulfide cycle of La Hedionda Spa.
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family: Tweedie (p = 1.116) |
|---|
| Link function: log |
| Formula: |
| Cyanobacteria ~ s(Sulfide, k = 6) + s(Phosphate, k = 6) |
| Parametric coefficients: |
| Estimate Standard Error t value Pr(>|t|) |
| (Intercept) −0.42269 0.03862 −10.94 0.00035 *** |
| Approximate significance of smooth terms: |
| edf Ref.df F p-value |
| s(Sulfide) 4.056 4.305 38.795 0.000548 *** |
| s(Phosphate) 2.851 3.023 5.898 0.060124. |
| R2 (adjusted) = 0.993 Deviance explained = 99.3% |
| −REML = 2.5089 Scale est. = 0.010243 n = 12 |

References
- Jiménez-Sánchez, J.; De la Hera Portillo, A.; Rubio Campos, J.C.; Hueso-Quesada, L.M. Informe de caracterización hidrogeológica y propuesta de protección de manantiales y lugares de interés hidrogeológico (Málaga). In Plan de Conservación, Recuperación y Puesta en Valor de Manantiales y Lugares de Interés Hidrogeológico de Andalucía (Estrategia de Conservación de los Ecosistemas Acuáticos Relacionados con las Masas de Agua Subterránea)—MA-4 Baños de la Hedionda; Estirado Oliet, M., Rubio Campos, J.C., Espina Argüello, J., García Padilla, M., Fernández-Palacios Carmona, J.M., Cañizares García, M.I., Eds.; 2011; Available online: http://info.igme.es/SidPDF/167000/081/167081_0000001.pdf (accessed on 1 December 2019).
- Martín-Clemente, E.; Melero-Jiménez, I.J.; Bañares-España, E.; Flores-Moya, A.; García-Sánchez, M.J. Adaptation dynamics and evolutionary rescue under sulfide selection in cyanobacteria: a comparative study between Microcystis aeruginosa and Oscillatoria sp. (cyanobacteria). J. Phycol. 2019, 55, 1348–1360. [Google Scholar] [CrossRef] [PubMed]
- IGME-JA. Evaluación del Estado Actual de las Aguas Minerales en la Comunidad Autónoma de Andalucía (Convenio Marco de asistencia técnica entre el Instituto Tecnológico y Geominero de España y la Consejería de Economía y Hacienda de la Junta de Andalucía) 1991. Available online: http://info.igme.es/SidPDF%5C034000%5C671%5CMemoria%5C34671_0001.pdf (accessed on 1 December 2019).
- Flores-Moya, A.; Costas, E.; Bañares-España, E.; García-Villada, L.; Altamirano, M.; López-Rodas, V. Adaptation of Spirogyra insignis (Chlorophyta) to an extreme natural habitat environment (sulphureous waters) through preselective mutations. New Phytolog. 2005, 166, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Oren, A.; Padan, E.; Malkin, S. Sulfide inhibition of Photosystem II in cyanobacteria (blue-green algae) and tobacco chloroplasts. BBA Bioenerg. 1979, 546, 270–279. [Google Scholar] [CrossRef]
- Miller, S.R.; Bebout, B.M. Variation in sulfide tolerance of photosystem II in phylogenetically diverse cyanobacteria from sulfidic habitats. Appl. Environ. Microb. 2004, 70, 736–744. [Google Scholar] [CrossRef]
- Bagarinao, T. Sulphide as an environmental factor and toxicant: tolerance and adaptations in aquatic organisms. Aquat. Toxicol. 1992, 24, 21–62. [Google Scholar] [CrossRef]
- Stal, L.J. Physiological ecology of cyanobacteria in microbial mats and other communities. New Phytol. 1995, 131, 1–32. [Google Scholar] [CrossRef]
- Stal, L.J. Cyanobacterial mats and stromatolites. In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Whitton, B.A., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 65–125. [Google Scholar]
- Castenholz, R.W. The effect of sulfide on the blue-green algae of hot springs II. Yellowstone National Park. Microb. Ecol. 1977, 3, 79–105. [Google Scholar] [CrossRef]
- Cohen, Y.; Jørgensen, B.B.; Revsbech, N.P.; Poplawski, R. Adaptation to hydrogen sulfide of oxygenic and anoxygenic photosynthesis among cyanobacteria. Appl. Environ. Microbiol. 1986, 51, 398–407. [Google Scholar] [CrossRef]
- Dodds, W.K.; Castenholz, R.W. Sulfide and pH effects on variable fluorescence of photosystem II in two strains of the cyanobacterium Oscillatoria amphigranulata. Photosynth. Res. 1990, 24, 265–271. [Google Scholar] [CrossRef]
- Cohen, Y.; Jørgensen, B.B.; Paden, E.; Shilo, M. Sulfide dependent anoxygenic photosynthesis in the cyanobacterium Oscillatoria limnetica. Nature 1975, 257, 489–492. [Google Scholar] [CrossRef]
- Cohen, Y.; Padan, E.; Shilo, M. Facultative anoxygenic photosynthesis in the cyanobacterium Oscillatoria limnetica. J. Bacteriol. 1975, 123, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Leister, D. Experimental evolution in photoautotrophic microorganisms as a means of enhancing chloroplast functions. Essays Biochem. 2017, 62, 77–84. [Google Scholar] [CrossRef]
- Walter, J.M.; Coutinho, F.H.; Dutilh, B.E.; Swings, J.; Thompson, F.L.; Thompson, C.C. Ecogenomics and taxonomy of Cyanobacteria phylum. Front. Microbiol. 2017, 8, 2132. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Arjona, M.M.; Bañares-España, E.; García-Sánchez, M.J.; Hernández-López, M.; López-Rodas, V.; Costas, E.; Flores-Moya, A. Disentangling mechanisms involved in the adaptation of photosynthetic microorganisms to the extreme sulphureous water from Los Baños de Vilo (S Spain). Microb. Ecol. 2013, 66, 742–751. [Google Scholar] [CrossRef]
- Bañares-España, E.; Fernández-Arjona, M.M.; García-Sánchez, M.J.; Hernández-López, M.; Reul, A.; Mariné, M.H.; Flores-Moya, A. Sulphide resistance in the cyanobacterium Microcystis aeruginosa: a comparative study of morphology and photosynthetic performance between the sulphide-resistant mutant and the wild-typestrain. Microb. Ecol. 2016, 71, 860–872. [Google Scholar] [CrossRef]
- Sommer, U.; Adrian, R.; De Senerpont Domis, L.; Elser, J.J.; Gaedke, U.; Ibelings, B.; Jeppesen, E.; Lürling, M.; Molinero, J.C.; Mooij, W.M.; et al. Beyond the Plankton Ecology Group (PEG) Model: Mechanisms Driving Plankton Succession. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 429–448. [Google Scholar] [CrossRef]
- Demoulin, C.F.; Lara, Y.L.; Cornet, L.; François, C.; Baurain, D.; Wilmotte, A.; Javaux, E.J. Cyanobacteria evolution: Insight from the fossil record. Free Radic. Biol. Med. 2019, 140, 206–223. [Google Scholar] [CrossRef]
- Whitton, B.A.; Potts, M. Introduction to the Cyanobacteria. In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Whitton, B.A., Ed.; Springer: Dordrecht, Netherlands, 2012; pp. 1–13. [Google Scholar] [CrossRef]
- Pachmayr, F. Vorkommen und Bestimmung von Schwefelverbindungen in Mineralwasser. Ph.D. Thesis, University München, München, Germany, 1960. [Google Scholar]
- Cline, J.D. Spectrophotometric determination of hydrogen sulfide in natural waters 1. Limnol. Oceanogr. 1969, 14, 454–458. [Google Scholar] [CrossRef]
- Slawyk, G.; MacIsaac, J.J. Comparison of two automated ammonium methods in a region of coastal upwelling. Deep Sea Res. 1972, 19, 521–524. [Google Scholar] [CrossRef]
- Beutler, M.; Wiltshire, K.H.; Meyer, B.; Moldaenke, C.; Lüring, M.; Meyerhöfer, U.-P.; Hansen, H.D. A fluorometric method for the differentiation of algal populations in vivo and in situ. Photosynth. Res. 2002, 72, 39–53. [Google Scholar] [CrossRef]
- Leboulanger, C.; Bouvy, M.; Carré, C.; Cecchi, P.; Amalric, L.; Bouchez, A.; Pagano, M.; Sarazin, G. Comparison of the effects of two herbicides and an insecticide on tropical freshwater plankton in microcosms. Arch. Environ. Contam. Toxicol. 2011, 61, 599–613. [Google Scholar] [CrossRef] [PubMed]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota 2. Teil/Part 2: Oscillatoriales Süßwasserflora von Mitteleuropa. In Freshwater Flora of Central Europe; Publisher Akademischer Verlag: Heidelberg, Germany, 2005; p. 772. [Google Scholar]
- Reul, A.; Muñoz, M.; Bautista, B.; Neale, P.J.; Sobrino, C.; Mercado, J.M.; Segovia, M.; Salles, S.; Kulk, G.; León, P.; et al. Effect of CO2, nutrients and light on coastal plankton. III. Trophic cascade, size structure and composition. Aquat. Biol. 2014, 22, 59–76. [Google Scholar] [CrossRef]
- Metorological Station of Estepona. Available online: http://www.juntadeandalucia.es/agriculturaypesca/ifapa/ria/servlet/FrontController?action=Static&url=coordenadas.jsp&c_provincia=29&c_estacion=4 (accessed on 15 June 2017).
- Köppen, W. Versuch einer Klassifikation der Klimate, vorzugsweise nach ihren Beziehungen zur Pflanzenwelt. Geogr. Zeitschr. 1900, 6, 593–611. [Google Scholar]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rube, F. World Map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Hastie, T.J.; Tibshirani, R. Generalized Additive Models. Stat. Sci. 1986, 1, 297–310. [Google Scholar] [CrossRef]
- Sivaraja, M.; Hunziker, D.; Dismukes, G.C. The reaction of H2S with the photosynthetic water-oxidizing complex and its lack of reaction with the primary electron acceptor in spinach. Biochim. Biophys. Acta 1988, 936, 228–235. [Google Scholar] [CrossRef]






| Summer | Winter | Differences | |
|---|---|---|---|
| Cyanobacteria concentration (Oscillatoria sp., Chl a µg L−1) | 1.6 (13) | 0.4 (9) | ** |
| Relative contribution of cyanobacteria (Oscillatoria sp.) to total Chl a concentration | 85% (13) | 67% (9) | * |
| Sulfide concentration (µM) | 147 ± 39 (13) | 13 ± 27 (9) | ** |
| Diatoms and dinoflagellates concentration(Chl a µg L−1) | 0.02 (13) | 0.02 (9) | ns |
| Green algae concentration(Chl a µg L−1) | 0.2 (13) | 0.1 (9) | ns |
| Cryptomonads concentration(Chl a µg L−1) | 0.05 (13) | 0.03 (9) | ns |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reul, A.; Martín-Clemente, E.; Melero-Jiménez, I.J.; Bañares-España, E.; Flores-Moya, A.; García-Sánchez, M.J. What Triggers the Annual Cycle of Cyanobacterium Oscillatoria sp. in an Extreme Environmental Sulfide-Rich Spa? Water 2020, 12, 883. https://doi.org/10.3390/w12030883
Reul A, Martín-Clemente E, Melero-Jiménez IJ, Bañares-España E, Flores-Moya A, García-Sánchez MJ. What Triggers the Annual Cycle of Cyanobacterium Oscillatoria sp. in an Extreme Environmental Sulfide-Rich Spa? Water. 2020; 12(3):883. https://doi.org/10.3390/w12030883
Chicago/Turabian StyleReul, Andreas, Elena Martín-Clemente, Ignacio J. Melero-Jiménez, Elena Bañares-España, Antonio Flores-Moya, and María J. García-Sánchez. 2020. "What Triggers the Annual Cycle of Cyanobacterium Oscillatoria sp. in an Extreme Environmental Sulfide-Rich Spa?" Water 12, no. 3: 883. https://doi.org/10.3390/w12030883
APA StyleReul, A., Martín-Clemente, E., Melero-Jiménez, I. J., Bañares-España, E., Flores-Moya, A., & García-Sánchez, M. J. (2020). What Triggers the Annual Cycle of Cyanobacterium Oscillatoria sp. in an Extreme Environmental Sulfide-Rich Spa? Water, 12(3), 883. https://doi.org/10.3390/w12030883