Assessment of Biological and Sanitary Condition of Alien Fish from a High-Mountain Lake (Cottian Alps)

 ,
,  ,
, 
 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Physicochemical Parameters of Water and Nutrients
2.3. Fish Sampling
2.4. Stomach Content Analysis
2.5. Sanitary Condition Evaluation
2.6. Biometric Measures and Statistical Analysis
3. Results
3.1. Hydrochemistry
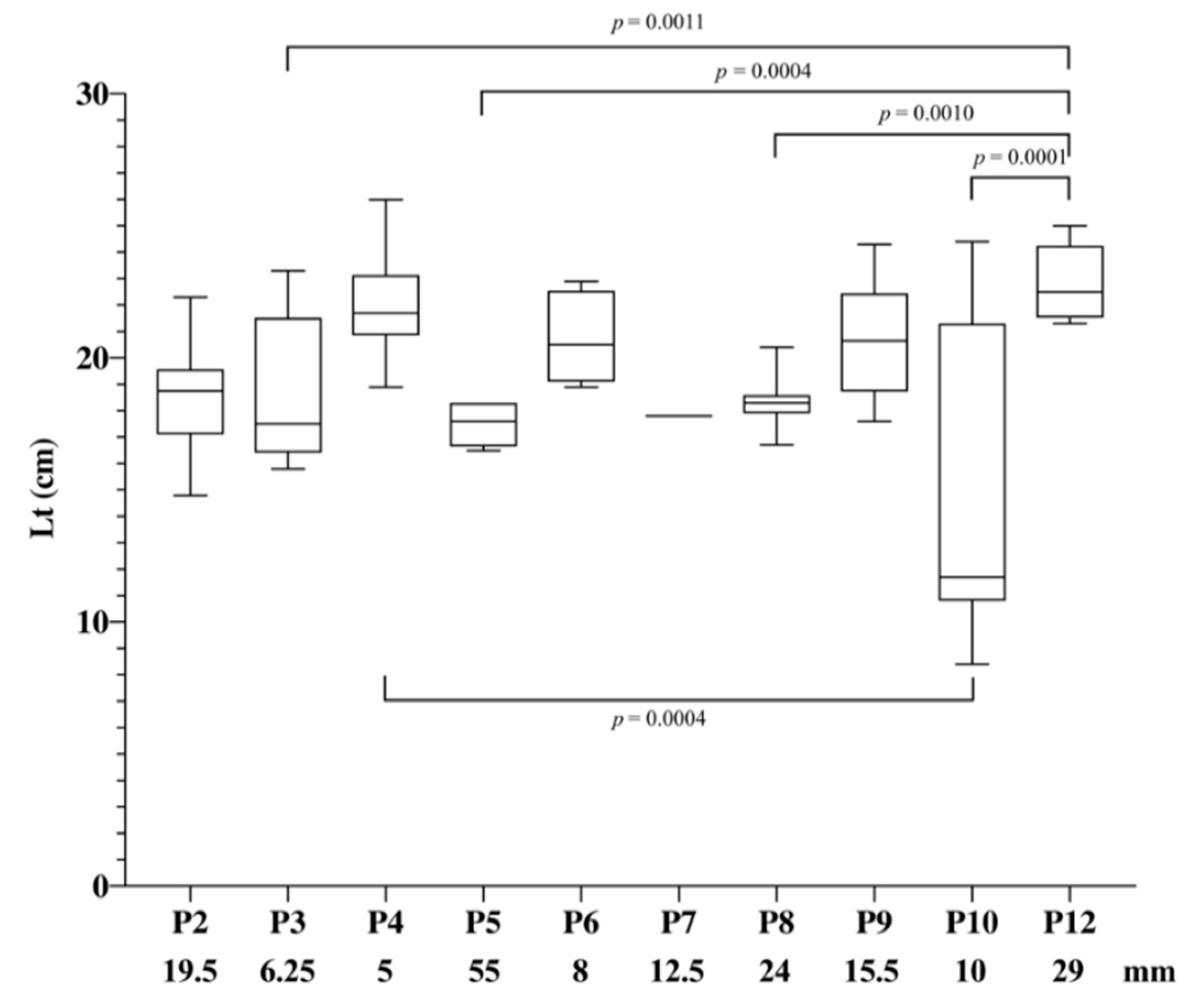
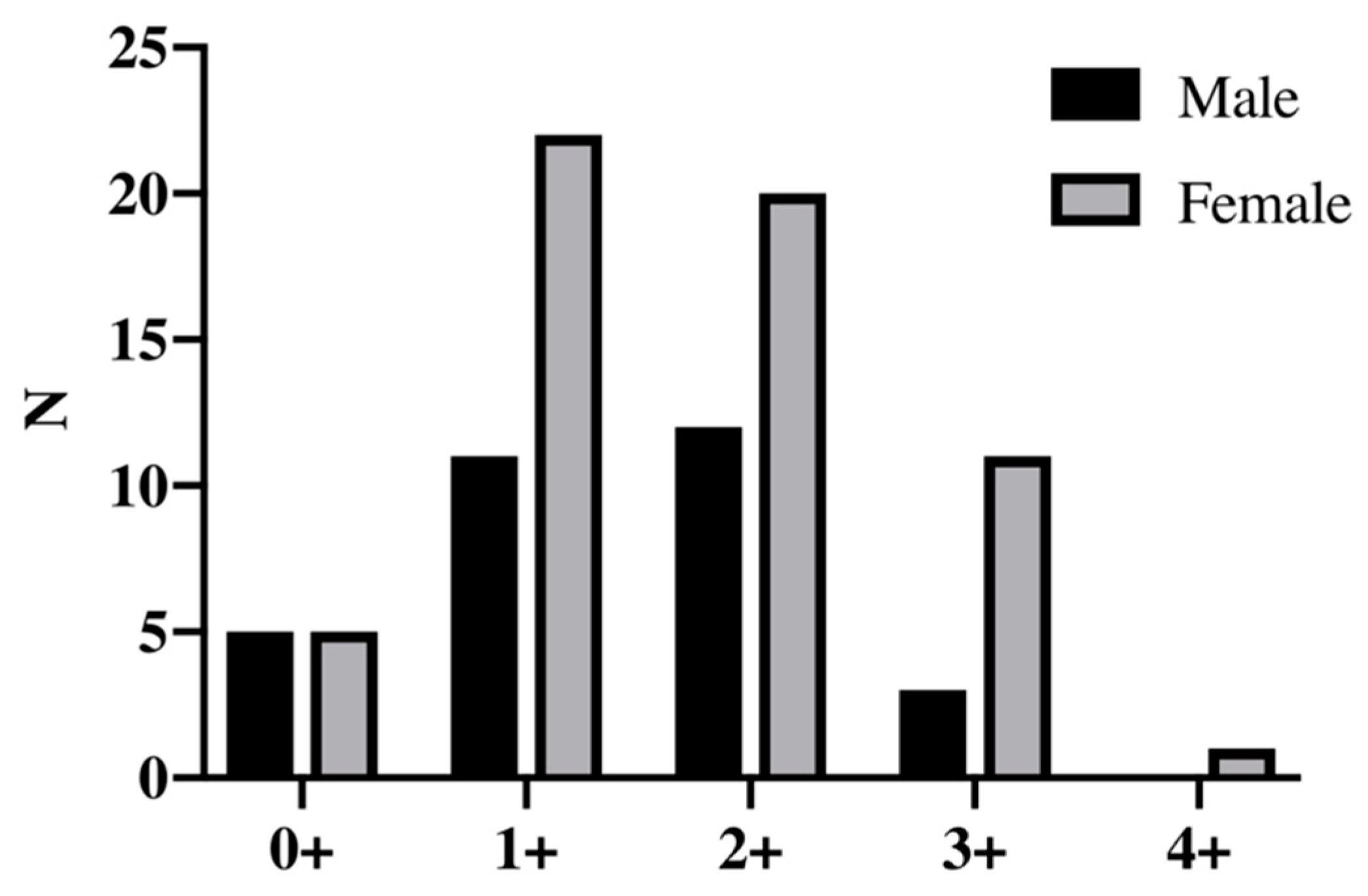
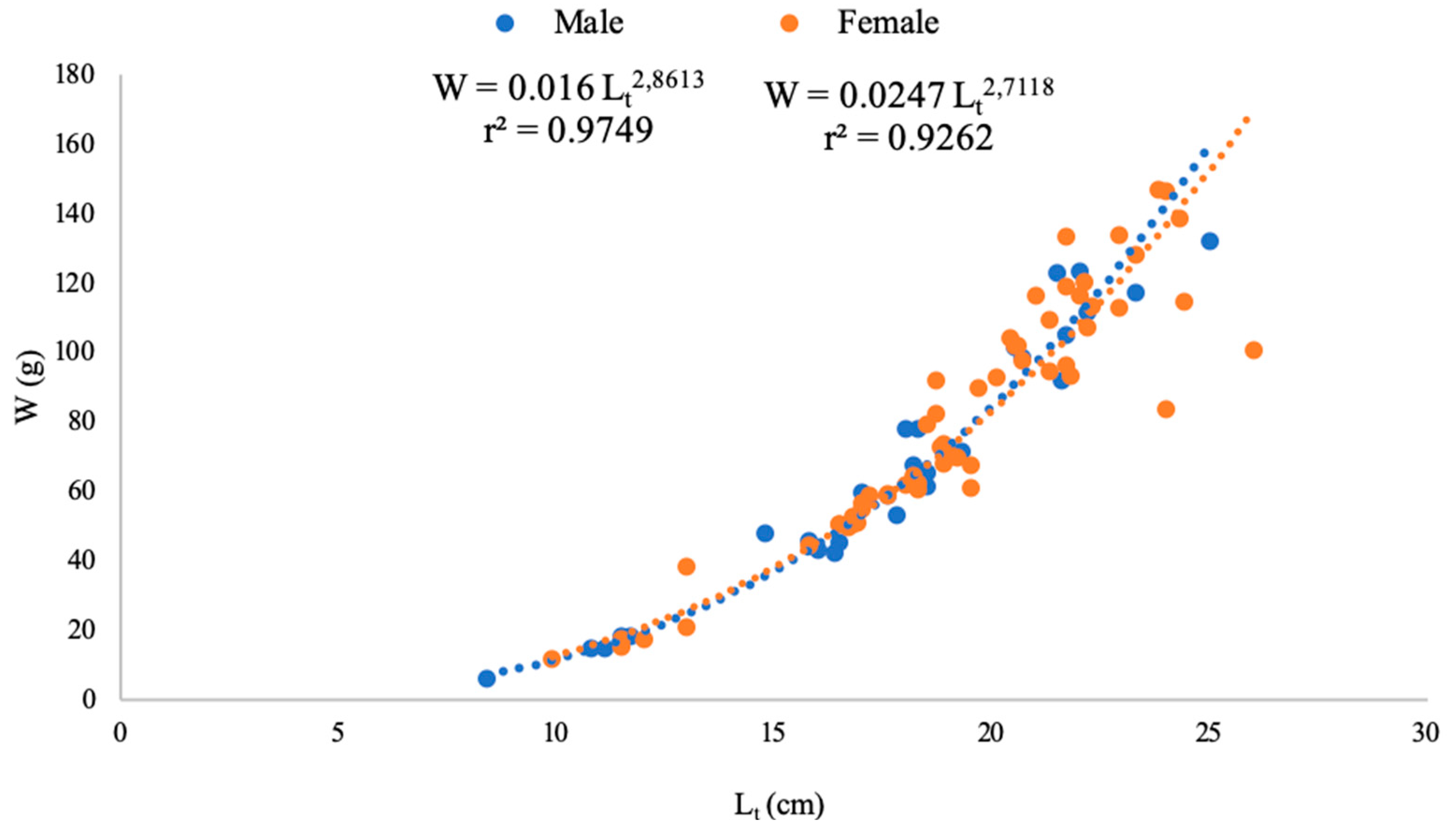
3.2. Fish Population
3.3. Sanitary Condition of Fish
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Catalan, J.; Camarero, L.; Felip, M.; Pla, S.; Ventura, M.; Buchaca, T.; Medina-Sánchez, J.M. High mountain lakes: Extreme habitats and witnesses of environmental changes. Limnetica 2006, 25, 551–584. [Google Scholar]
- Pastorino, P.; Pizzul, E.; Bertoli, M.; Perilli, S.; Brizio, P.; Salvi, G.; Esposito, G.; Abete, M.C.; Prearo, M.; Squadrone, S. Macrobenthic invertebrates as bioindicators of trace elements in high-mountain lakes. Environ. Sci. Pollut. Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Knapp, R.A.; Corn, P.S.; Schindler, D.E. The introduction of nonnative fish into wilderness lakes: Good intentions, conflicting mandates, and unintended consequences. Ecosystems 2001, 4, 275–278. [Google Scholar] [CrossRef]
- Miró, A.; Ventura, M. Historical use, fishing management and lake characteristics explain the presence of non-native trout in Pyrenean lakes: Implications for conservation. Biol. Conserv. 2013, 167, 17–24. [Google Scholar] [CrossRef]
- Pister, E.P. Wilderness fish stocking: History and perspective. Ecosystems 2001, 4, 279–286. [Google Scholar] [CrossRef]
- Schindler, D.E.; Parker, B.R. Biological pollutants: Alien fishes in mountain lakes. Water Air Soil Poll. 2002, 2, 379–397. [Google Scholar] [CrossRef]
- Tiberti, R.; von Hardenberg, A.; Bogliani, G. Ecological impact of introduced fish in high altitude lakes: A case of study from the European Alps. Hydrobiologia 2014, 724, 1–19. [Google Scholar] [CrossRef]
- Pastorino, P.; Bona, C.; Foglini, C.; Maurella, C.; Righetti, M.; Burioli, E.; Ru, G.; Pizzul, E.; Prearo, M. Health risks related to freshwater fish handling and introduction of alien species. IJFI 2017, 4, 109–116. [Google Scholar]
- Gozlan, R.E.; St-Hilaire, S.; Feist, S.W.; Martin, P.; Kent, M.L. Disease threat to European fish. Nature 2005, 435, 1046. [Google Scholar] [CrossRef]
- Pastorino, P.; Polazzo, F.; Bertoli, M.; Santi, M.; Righetti, M.; Pizzul, E.; Prearo, M. Consequences of fish introduction in fishless Alpine lakes: Preliminary notes from a sanitary point of view. Turk. J. Fish. Aquat. Sci. 2020, 20, 1–8. [Google Scholar] [CrossRef]
- Cantonati, M.; Lapini, L.; Paradisi, S.; Stoch, F. Conservation and management. In High-Altitude Lakes, Quaderni Habitat. Ministero dell’Ambiente e Della Tutela del Territorio, English ed.; Stoch, F., Ed.; Museo Friulano di Storia Naturale: Udine, Italy, 2006; Volume 4, pp. 123–135. [Google Scholar]
- MacCrimmon, H.R.; Gots, B.L.; Campbell, J.S. World distribution of Brook Trout, Salvelinus fontinalis: Further observations. Can. J. Fish. Aquat. Sci. 1971, 28, 452–456. [Google Scholar] [CrossRef]
- Savini, D.; Occhipinti-Ambrogi, A.; Marchini, A.; Tricarico, E.; Gherardi, F.; Olenin, S.; Gollasch, S. The top 27 animal alien species introduced into Europe for aquaculture and related activities. J. Appl. Ichthyol. 2010, 26, 1–7. [Google Scholar] [CrossRef]
- Pastorino, P.; Elia, A.C.; Caldaroni, B.; Menconi, V.; Abete, M.C.; Brizio, P.; Bertoli, M.; Zaccaroni, A.; Magara, G.; Dörr, A.M.J.; et al. Oxidative stress ecology in brook trout (Salvelinus fontinalis) from a high-mountain lake (Cottian Alps). Sci. Total Environ. 2020. [Google Scholar] [CrossRef] [PubMed]
- APHA; AWWA; WEF. Standard Methods for the Examination of Water and Wastewater, 20th ed.; American Public Health Association, American Water Work Association, Water Environment Federation: Washington, DC, USA, 1998. [Google Scholar]
- Volta, P. Indice per L’analisi Dello Stato di Qualità Della Fauna Ittica Finalizzato Alla Valutazione Dello Stato Ecologico dei Laghi Italiani: Lake Fish Index (LFI). Indici per la Valutazione Della Qualità Ecologica dei Laghi. CNR, Istituto per lo Studio degli Ecosistemi, Verbania Pallanza. Available online: https://core.ac.uk/download/pdf/37834548.pdf (accessed on 30 December 2019).
- CEN. Water Quality-Sampling of Fish with Multi-mesh Gillnets (EN 14757: 2005); CEN: Brussels, Belgium, 2005. [Google Scholar]
- Tiberti, R.; Brighenti, S.; Canedoli, C.; Iacobuzio, R.; Rolla, M. The diet of introduced brook trout (Salvelinus fontinalis; Mitchill, 1814) in an alpine area and a literature review on its feeding ecology. J. Limnol. 2016, 75, 488–507. [Google Scholar] [CrossRef]
- Squadrone, S.; Brizio, P.; Stella, C.; Prearo, M.; Pastorino, P.; Serracca, L.; Ercolini, C.; Abete, M.C. Presence of trace metals in aquaculture marine ecosystems of the northwestern Mediterranean Sea (Italy). Environ. Pollut. 2016, 215, 77–83. [Google Scholar] [CrossRef]
- Cavalli, L.; Chappaz, R.; Bouchard, P.; Brun, G. Food availability and growth of the brook trout, Salvelinus fontinalis (Mitchill), in a French Alpine lake. Fish. Manag. Ecol. 1997, 4, 167–177. [Google Scholar] [CrossRef]
- Alexander, T.J.; Vonlanthen, P.; Periat, G.; Degiorgi, F.; Raymond, J.C.; Seehausen, O. Estimating whole-lake fish catch per unit effort. Fish. Res. 2015, 172, 287–302. [Google Scholar] [CrossRef]
- Zymonas, N.D.; McMahon, T.E. Comparison of pelvic fin rays, scales and otoliths for estimating age and growth of bull trout, Salvelinus confluentus. Fish. Manag. Ecol. 2009, 16, 155–164. [Google Scholar] [CrossRef]
- Ricker, W.E. Computation and Interpretation of Biological Statistics of Fish Populations; Department of the Environment, Fisheries and Marine Service: Ottawa, ON, Canada, 1975; pp. 1–382.
- Treer, T.; Sprem, N.; Torcu-Koc, H.; Sun, Y.; Piria, M. Short communication Length-weight relationships of freshwater fishes of Croatia. J. Appl. Ichthyol. 2008, 24, 626–628. [Google Scholar] [CrossRef]
- Froese, R. Cube law, condition factor and weight-length relationships: History, meta-analysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- Tiberti, R.; Tartari, G.A.; Marchetto, A. Geomorphology and hydrochemistry of 12 Alpine lakes in the Gran Paradiso National Park, Italy. J. Limnol. 2010, 69, 242–256. [Google Scholar] [CrossRef]
- Camarero, L.; Rogora, M.; Mosello, R.; Anderson, N.J.; Barbieri, A.; Botev, I.; Kernan, M.; Kopàćek, J.; Korhola, A.; Lotter, A.F.; et al. Regionalization of chemical variability in European mountain lakes. Freshw. Biol. 2009, 54, 2452–2469. [Google Scholar] [CrossRef]
- Boggero, A.; Füreder, L.; Lencioni, V.; Simcic, T.; Thaler, B.; Ferrarese, U.; Lotter, A.F.; Ettinger, R. Littoral chironomid communities of Alpine lakes in relation to environmental factors. Hydrobiologia 2006, 562, 145–165. [Google Scholar] [CrossRef]
- Füreder, L.; Ettinger, R.; Boggero, A.; Thaler, B.; Thies, H. Macroinvertebrate diversity in Alpine lakes: Effects of altitude and catchment properties. Hydrobiologia 2006, 652, 123–144. [Google Scholar] [CrossRef]
- Fjellheim, A.; Raddum, G.V.G.; Vandvik, V.; Cogălniceanu, D.; Boggero, A.; Brancelj, A.; Galas, J.; Sporka, F.; Vidinova, Y.; Bitusik, P.; et al. Diversity and distribution patterns of benthic invertebrates along alpine gradients. A study of remote European freshwater lakes. Adv. Limnol. 2009, 62, 167–190. [Google Scholar]
- Tiberti, R.; Nelli, L.; Brighenti, S.; Iacobuzio, R.; Rolla, M. Spatial distribution of introduced brook trout Salvelinus fontinalis (Salmonidae) within alpine lakes: Evidences from a fish eradication campaign. Eur. Zool. J. 2017, 84, 73–88. [Google Scholar] [CrossRef]
- Adams, P.; James, C.; Speas, C. Brook trout (Salvelinus fontinalis). Species and Conservation Assessment. Grand Mesa, Uncompahgre, and Gunnison National Forests. Available online: https://www.fs.usda.gov/Internet/FSE_DOCUMENTS/stelprdb5199816.pdf (accessed on 30 December 2019).
- Alessio, G.; Forneris, G.; Confortini, I. Indagini preliminari su accrescimento, biologia e ambientamento del salmerino di fontana, Salvelinus fontinalis Mitch., in un ecosistema lacustre di alta quota. Atti Società Italiana Scienze Naturali 1987, 128, 138–152. [Google Scholar]
- Forneris, G. Piemonte. Acque. Pesci. Pesca; EDA: Torino, Italy, 1984; pp. 1–96. [Google Scholar]
- Šanda, R.; Miroslav Švátora, M.; Vukić, J.; Hořicka, Z.; Sychrová, O.; Marić, S. Evaluation of the temporal development of three introduced populations of brook trout, Salvelinus fontinalis, in Jizerské Mountains (Czech Republic). Arch. Biol. Sci. 2015, 67, 775–784. [Google Scholar] [CrossRef]
- Dutil, J.D.; Power, G. Validité de la lecture das otolithes comparée à celle de la lecture des écailles pour la détermination de l’âge de l’omble de fontaine (Salvelinus fontinalis). Nat. Can. 1977, 104, 361–367. [Google Scholar]
- Kozel, S.J.; Hubert, W.A. Age estimates of brook trout from high-elevation Rocky Mountain streams using scales and otoliths. Northwest Sci. 1987, 61, 216–219. [Google Scholar]
- McFadden, J.T. A population study of the Brook Trout, Salvelinus fontinalis. Wildl. Monogr. 1961, 7, 3–73. [Google Scholar]
- Wydoski, R.S.; Cooper, E.L. Maturation and fecundity of brook trout from infertile streams. Can. J. Fish. Aquat. Sci. 1966, 23, 623–649. [Google Scholar] [CrossRef]
- Flick, W.A.; Webster, D.A. Production of wild, domestic, and interstrain hybrids of brook trout (Salvelinus fontinalis) in natural ponds. JFRBC 1976, 33, 1525–1539. [Google Scholar] [CrossRef]
- Quinn, N.W.; Korver, R.M.; Hicks, F.J.; Monroe, B.P.; Hawkins, R.R. An empirical model of lentic brook trout. N. Am. J. Fish. Manag. 1994, 14, 692–709. [Google Scholar] [CrossRef]
- Ospina-Alvarez, N.; Piferrer, F. Temperature-dependent sex determination in fish revisited: Prevalence, a single sex ratio response pattern, and possible effects of climate change. PLoS ONE 2008, 3, e2837. [Google Scholar] [CrossRef] [PubMed]
- Pavlidis, M.; Koumoundouros, G.; Sterioti, A.; Somarakis, S.; Divanach, P.; Kentouri, M. Evidence of temperature-dependent sex determination in the European sea bass (Dicentrarchus labrax L.). J. Exp. Zool. 2000, 287, 225–232. [Google Scholar] [CrossRef]
- Piferrer, F.; Anastasiadi, D.; Valdivieso, A.; Sánchez-Baizán, N.; Moraleda-Prados, J.; Ribas, L. The Model of the Conserved Epigenetic Regulation of Sex. Front. Genet. 2019, 10, 857. [Google Scholar] [CrossRef] [PubMed]
- Reimers, N. Conditions of existence, growth, and longevity of brook trout in a small high-altitude lake of the Eastern Sierra Nevada. Calif. Fish. Game 1958, 44, 319–333. [Google Scholar]
- Dawidowicz, P.; Gliwicz, Z.M. Food of brook charr in extreme oligotrophic conditions of an alpine lake. Environ. Biol. Fish. 1983, 8, 55–60. [Google Scholar] [CrossRef]
- Utz, R.M.; Hartman, K.J. Identification of critical prey items to Appalachian brook trout (Salvelinus fontinalis) with emphasis on terrestrial organisms. Hydrobiologia 2007, 575, 259–270. [Google Scholar] [CrossRef]
- Lacasse, S.; Magnan, P. Biotic and abiotic determinants of the diet of brook trout, Salvelinus fontinalis, in lakes of the Laurentian Shield. Can. J. Fish. Aquat. Sci. 1992, 49, 1001–1009. [Google Scholar] [CrossRef]
- Sotiropoulos, J.C.; Nislow, K.H.; Ross, M.R. Brook trout, Salvelinus fontinalis, microhabitat selection and diet under low summer stream flows. Fish. Manag. Ecol. 2006, 13, 149–155. [Google Scholar] [CrossRef]
- Kownacki, A.; Dumnicka, E.; Kwandrans, J.; Galas, J.; Ollik, M. Benthic communities in relation to environmental factors in small high mountain ponds threatened by air pollutants. Boreal Environ. Res. 2006, 11, 481–492. [Google Scholar]
- Dumnicka, E.; Boggero, A. Freshwater Oligochaeta in two mountain ranges in Europe: The Tatra Mountains (Poland) and the Alps (Italy). Fund. Appl. Limnol. 2007, 168, 231–242. [Google Scholar] [CrossRef]
- Hyslop, E.J. Stomach contents analysis - a review of methods and their application. J. Fish. Biol. 1980, 17, 411–429. [Google Scholar] [CrossRef]
- Eby, L.A.; Roach, W.J.; Crowder, L.B.; Stanford, J.A. Effects of stocking-up freshwater food webs. Trends Ecol. Evol. 2006, 21, 576–584. [Google Scholar] [CrossRef]
- Nakano, S.; Murakami, M. Reciprocal subsidies: Dynamic interdependence between terrestrial and aquatic food webs. Proc. Natl. Acad. Sci. USA 2001, 98, 166–170. [Google Scholar] [CrossRef]
- Baxter, C.V.; Fausch, K.D.; Carl Saunders, W. Tangled webs: Reciprocal flows of invertebrate prey link streams and riparian zones. Freshw. Biol. 2005, 50, 201–220. [Google Scholar] [CrossRef]
- Baxter, C.V.; Fausch, K.D.; Murakami, M.; Chapman, P.L. Fish invasion restructures stream and forest food webs by interrupting reciprocal prey subsidies. Ecology 2004, 85, 2656–2663. [Google Scholar] [CrossRef]
- Knight, T.M.; McCoy, M.W.; Chase, J.M.; McCoy, K.A.; Holt, R.D. Trophic cascades across ecosystems. Nature 2005, 437, 880. [Google Scholar] [CrossRef]
- Leisner, J.J.; Laursen, B.G.; Prévost, H.; Drider, D.; Dalgaard, P. Carnobacterium: Positive and negative effects in the environment and in foods. FEMS Microbiol. Rev. 2007, 31, 592–613. [Google Scholar] [CrossRef] [PubMed]
- Ramia, N.E.; El Kheir, S.M.; Taha, S.; Mangavel, C.; Revol-Junelles, A.M.; Borges, F. Multilocus sequence typing of Carnobacterium maltaromaticum strains associated with fish disease and dairy products. Appl. Microbiol. 2019, 126, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Loch, T.P.; Xu, W.; Fitzgerald, S.M.; Faisal, M. Isolation of a Carnobacterium maltaromaticum-like bacterium from systemically infected lake whitefish (Coregonus clupeaformis). FEMS Microbiol. Lett. 2008, 288, 76–84. [Google Scholar] [CrossRef]
- Zlotkin, A.; Chilmonczyk, S.; Eyngor, M.; Hurvitz, A.; Ghittino, C.; Eldar, A. Trojan horse effect: Phagocyte-mediated Streptococcus iniae infection of fish. Infect. Immun. 2003, 71, 2318–2325. [Google Scholar] [CrossRef]
- Köck, G.; Triendl, M.; Hofer, R. Seasonal patterns of metal accumulation in Arctic char (Salvelinus alpinus) from an oligotrophic Alpine lake related to temperature. Can. J. Fish. Aquat. Sci. 1996, 53, 780–786. [Google Scholar] [CrossRef]
- European Commission. Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. OJEC 2006, 364, 324–365. [Google Scholar]
- Rognerud, S.; Grimalt, J.O.; Rosseland, B.O.; Fernandez, P.; Hofer, R.; Lackner, R.; Lien, L.; Massabuau, J.C.; Ribes, A. Mercury and organochlorine contamination in Brown trout (Salmo trutta) and Arctic charr (Salvelinus alpinus) from high mountain lakes in Europe and the Svalbard archipelago. Water Air Soil Pollut. 2002, 2, 209–232. [Google Scholar] [CrossRef]
- Council of the European Commission. Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. OJEC 1992, 206, 1–9.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Panel (P) | Mesh Size (mm) |
|---|---|
| 1 | 43 |
| 2 | 19.5 |
| 3 | 6.25 |
| 4 | 5 |
| 5 | 55 |
| 6 | 8 |
| 7 | 12.5 |
| 8 | 24 |
| 9 | 15.5 |
| 10 | 10 |
| 11 | 35 |
| 12 | 29 |
| Parameter | Mean ± SD | ||
|---|---|---|---|
| Temperature (°C) | 16.08 | ± | 0.54 |
| Dissolved oxygen concentration (mg L−1) | 6.82 | ± | 0.42 |
| Oxygen saturation (%) | 87.82 | ± | 6.36 |
| pH | 6.70 | ± | 0.34 |
| Conductivity (µS cm−1) | 18.74 | ± | 1.03 |
| NH4+ (mg L−1) | 0.09 | ± | 0.07 |
| NO3− (mg L−1) | 9.59 | ± | 1.67 |
| PO43− (mg L−1) | 0.02 | ± | 0.01 |
| Gillnet | A1 (n = 16) | A2 (n = 16) | B1 (n = 12) | B2 (n = 14) | B3 (n = 15) | B4 (n = 17) |
|---|---|---|---|---|---|---|
| CPUE | 0.100 | 0.100 | 0.267 | 0.311 | 0.333 | 0.378 |
| W1 | 78.26 | 67.53 | 58.72 | 116.53 | 18.41 | 18.45 |
| W2 | 146.86 | 79.55 | 73.58 | 98.35 | 17.29 | 15.12 |
| W3 | 120.16 | 53.16 | 64.59 | 45.81 | 58.75 | 17.34 |
| W4 | 92.80 | 94.46 | 71.24 | 50.49 | 14.75 | 20.87 |
| W5 | 114.85 | 96.58 | 44.52 | 54.76 | 11.70 | 38.23 |
| W6 | 109.4 | 61.10 | 128.00 | 65.32 | 6.07 | 43.02 |
| W7 | 93.45 | 50.64 | 133.41 | 50.90 | 14.70 | 50.46 |
| W8 | 97.53 | 45.44 | 122.89 | 61.75 | 102.2 | 56.54 |
| W9 | 73.02 | 62.71 | 107.24 | 60.65 | 138.63 | 59.87 |
| W10 | 52.64 | 58.78 | 100.94 | 92.05 | 131.99 | 61.67 |
| W11 | 113.44 | 111.75 | 105.23 | 49.63 | 83.74 | 69.98 |
| W12 | 77.91 | 71.39 | 72.06 | 62.25 | 91.99 | 67.43 |
| W13 | 89.88 | 133.82 | 59.32 | 117.30 | 61.07 | |
| W14 | 69.64 | 101.88 | 104.17 | 112.74 | 101.4 | |
| W15 | 47.85 | 68.02 | 42.17 | 116.32 | ||
| W16 | 82.21 | 118.96 | 123.25 | |||
| W17 | 146.7 | |||||
| BPUE | 8.85 | 7.73 | 24.05 | 21.60 | 21.39 | 23.73 |
| Age Class | Females (F) | Males (M) | Mann−Whitney Test (F vs. M) | ||||
|---|---|---|---|---|---|---|---|
| Lt (cm) | W (g) | Lt (cm) | W (g) | Lt | W | ||
| 0+ | Mean | 11.58 | 16.46 | 10.70 | 14.48 | ||
| Median | 11.50 | 17.29 | 11.10 | 14.75 | |||
| SD | 1.12 | 3.37 | 1.33 | 5.05 | p = 0.3016 | p = 0.0659 | |
| Min | 9.90 | 11.70 | 8.40 | 6.07 | |||
| Max | 11.50 | 20.87 | 11.70 | 18.41 | |||
| CV % | 9.68 | 20.45 | 12.00 | 34.24 | |||
| No. | 5 | 5 | |||||
| 1+ | Mean | 17.46 | 58.38 | 17.14 | 55.69 | ||
| Median | 17.40 | 58.73 | 17.00 | 53.16 | |||
| SD | 1.45 | 9.93 | 1.36 | 12.14 | |||
| Min | 13.00 | 38.23 | 14.80 | 42.17 | p = 0.4333 | p = 0.4625 | |
| Max | 19.50 | 79.55 | 19.00 | 78.26 | |||
| CV % | 8.28 | 17.00 | 7.99 | 22.83 | |||
| No. | 22 | 11 | |||||
| 2+ | Mean | 20.48 | 97.41 | 19.92 | 86.38 | ||
| Median | 20.55 | 99.70 | 19.90 | 84.95 | |||
| SD | 1.38 | 18.84 | 1.87 | 22.68 | |||
| Min | 18.30 | 62.25 | 17.60 | 58.78 | p = 0.2589 | p = 0.1821 | |
| Max | 22.90 | 133.41 | 23.30 | 122.89 | |||
| CV % | 6.76 | 19.34 | 9.39 | 26.25 | |||
| No. | 20 | 12 | |||||
| 3+ | Mean | 23.06 | 119.64 | 23.07 | 122.33 | ||
| Median | 23.30 | 118.96 | 22.20 | 123.25 | |||
| SD | 1.14 | 21.50 | 1.68 | 10.15 | |||
| Min | 21.30 | 83.74 | 22.00 | 123.25 | p = 0.9725 | p = 0.999 | |
| Max | 24.40 | 146.86 | 25.00 | 131.99 | |||
| CV % | 4.96 | 17.97 | 7.27 | 8.30 | |||
| No. | 11 | 3 | |||||
| 4+ | Mean | 26.00 | 100.94 | - | - | ||
| Median | - | - | - | - | |||
| SD | - | - | - | - | |||
| Min | - | - | - | - | |||
| Max | - | - | - | - | |||
| CV % | - | - | - | - | |||
| No. | 1 | 0 | |||||
| Age | Sex | Kmean |
|---|---|---|
| 0+ | M | 1.03 ± 0.04 |
| F | 1.14 ± 0.01 | |
| 1+ | M | 0.89 ± 0.02 |
| F | 1.08 ± 0.01 | |
| 2+ | M | 0.86 ± 0.02 |
| F | 1.05 ± 0.01 | |
| 3+ | M | 0.82 ± 0.02 |
| F | 1.03 ± 0.01 | |
| 4+ | M | - |
| F | 1.01 |
| Prey Group | Fi (%) |
|---|---|
| Benthic invertebrates | 7.55 |
| Diptera Chironomidae | 7.55 |
| Terrestrial invertebrates | 83.55 |
| Imenoptera | 68.00 |
| Formicidae | 1.33 |
| Coleoptera | 15.55 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pastorino, P.; Prearo, M.; Bertoli, M.; Menconi, V.; Esposito, G.; Righetti, M.; Mugetti, D.; Pederiva, S.; Abete, M.C.; Pizzul, E. Assessment of Biological and Sanitary Condition of Alien Fish from a High-Mountain Lake (Cottian Alps). Water 2020, 12, 559. https://doi.org/10.3390/w12020559
Pastorino P, Prearo M, Bertoli M, Menconi V, Esposito G, Righetti M, Mugetti D, Pederiva S, Abete MC, Pizzul E. Assessment of Biological and Sanitary Condition of Alien Fish from a High-Mountain Lake (Cottian Alps). Water. 2020; 12(2):559. https://doi.org/10.3390/w12020559
Chicago/Turabian StylePastorino, Paolo, Marino Prearo, Marco Bertoli, Vasco Menconi, Giuseppe Esposito, Marzia Righetti, Davide Mugetti, Sabina Pederiva, Maria Cesarina Abete, and Elisabetta Pizzul. 2020. "Assessment of Biological and Sanitary Condition of Alien Fish from a High-Mountain Lake (Cottian Alps)" Water 12, no. 2: 559. https://doi.org/10.3390/w12020559
APA StylePastorino, P., Prearo, M., Bertoli, M., Menconi, V., Esposito, G., Righetti, M., Mugetti, D., Pederiva, S., Abete, M. C., & Pizzul, E. (2020). Assessment of Biological and Sanitary Condition of Alien Fish from a High-Mountain Lake (Cottian Alps). Water, 12(2), 559. https://doi.org/10.3390/w12020559