Gradual Exposure to Salinity Improves Tolerance to Salt Stress in Rapeseed (Brassica napus L.)


 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Growth Conditions and Saline Treatments
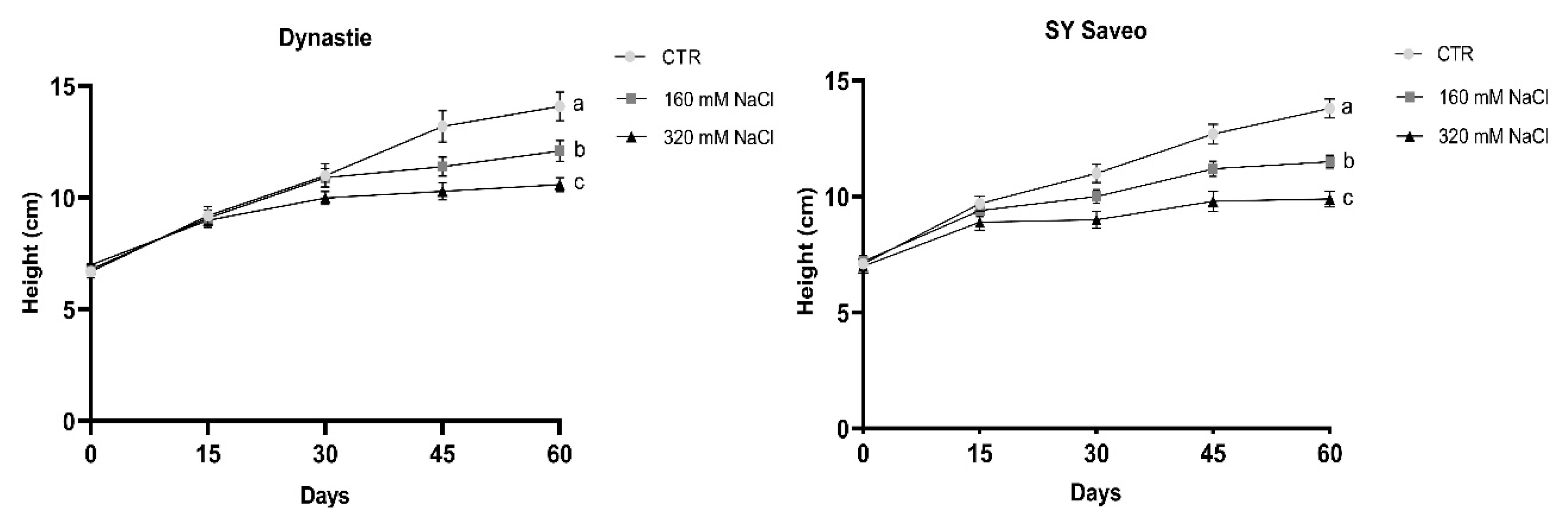
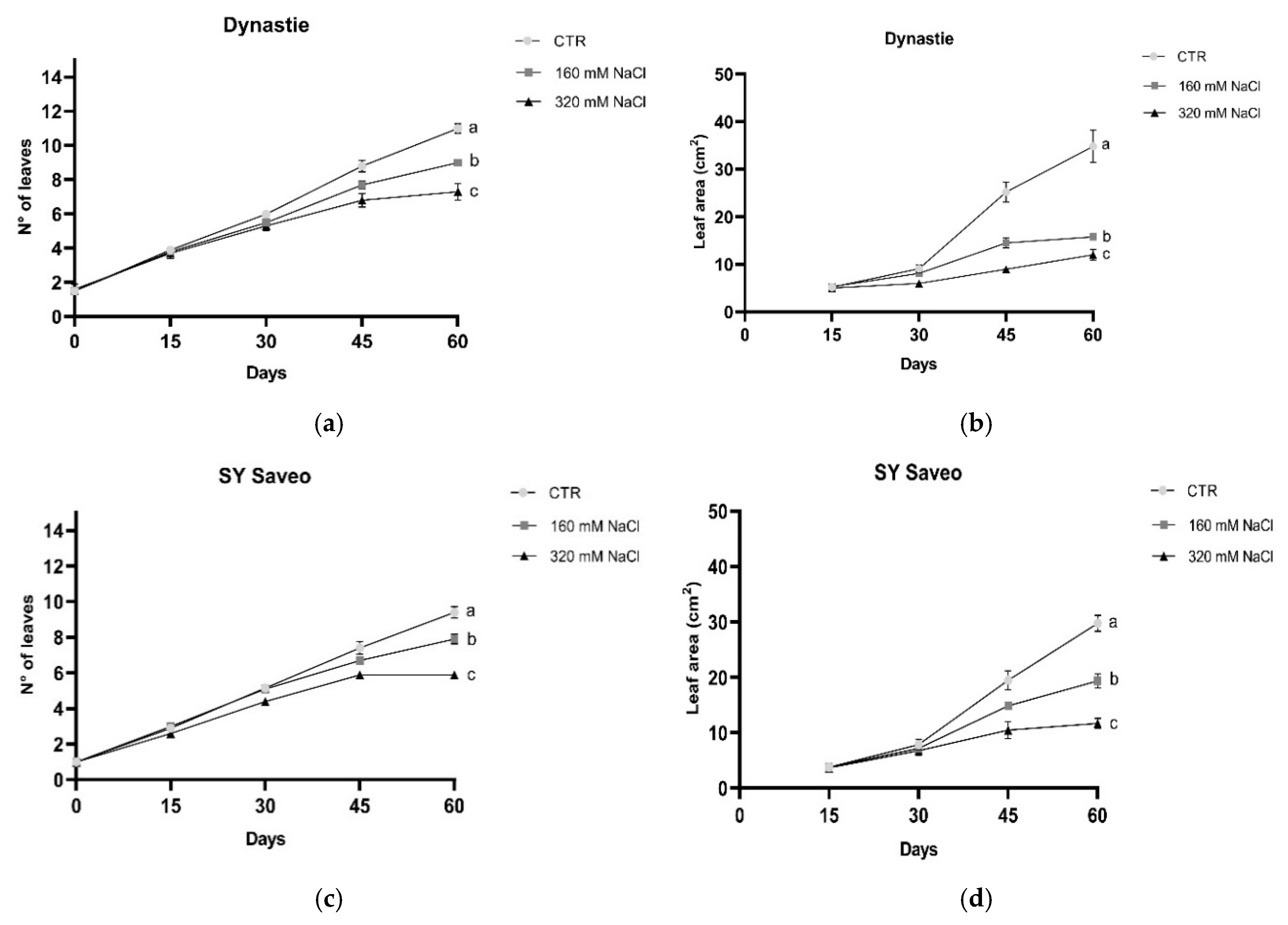
2.3. Growth and Morphological Parameters
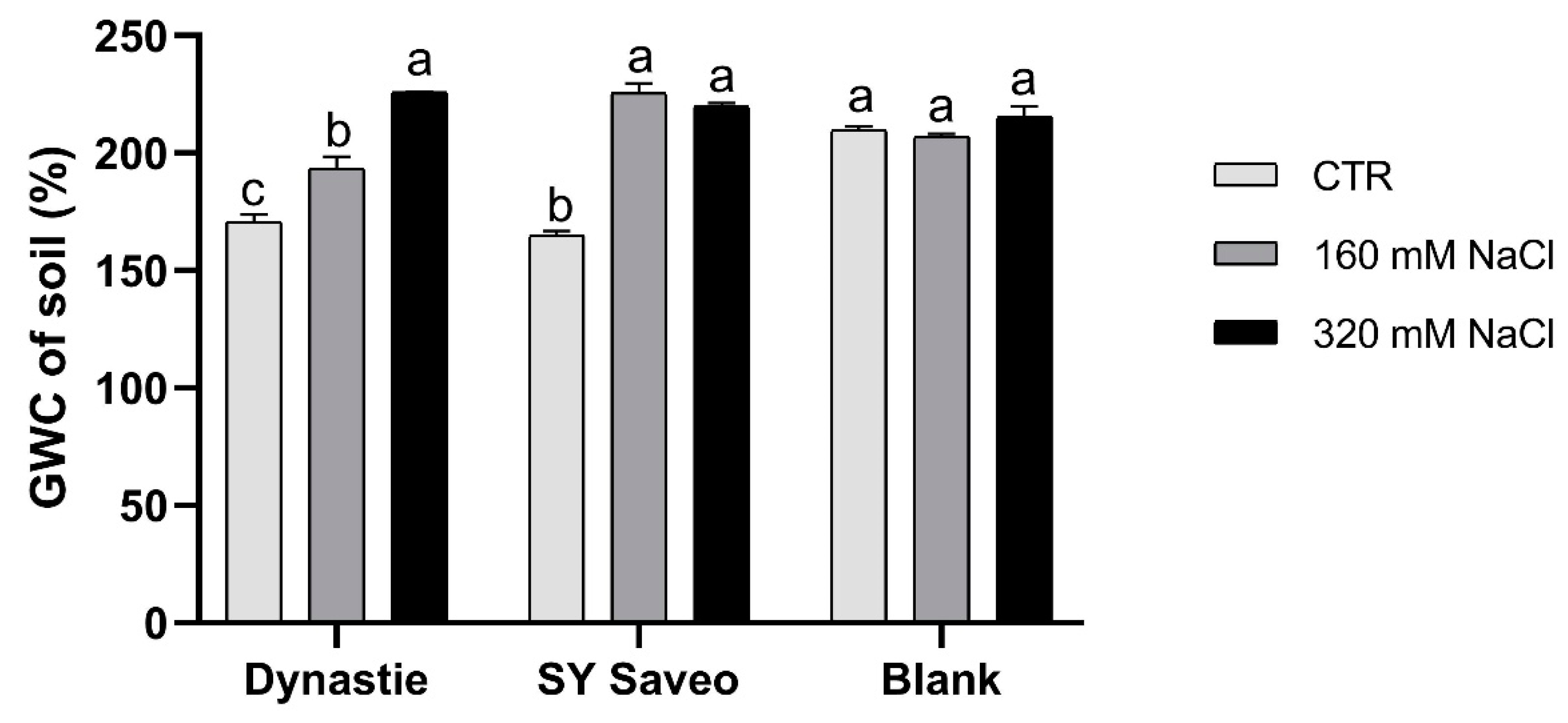
2.4. Water Content of the Plants
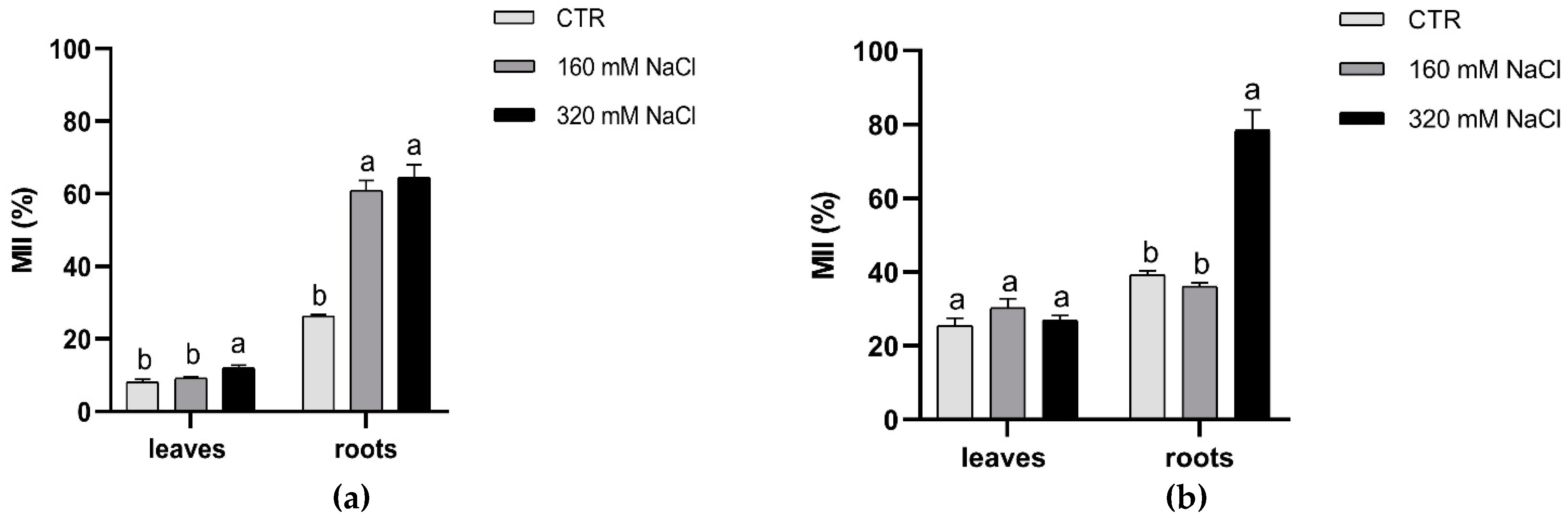
2.5. Membrane Injury Index (MII)
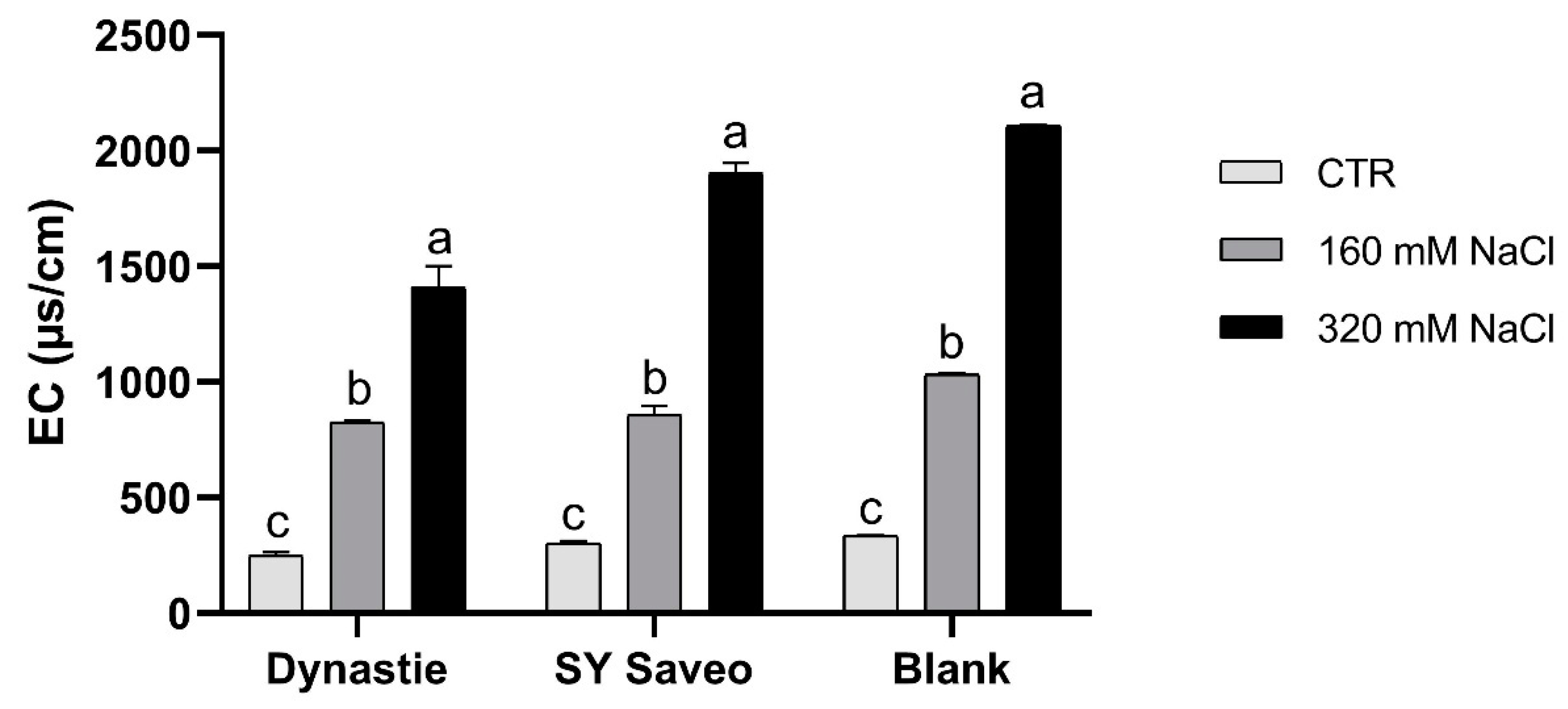
2.6. Soil Analysis
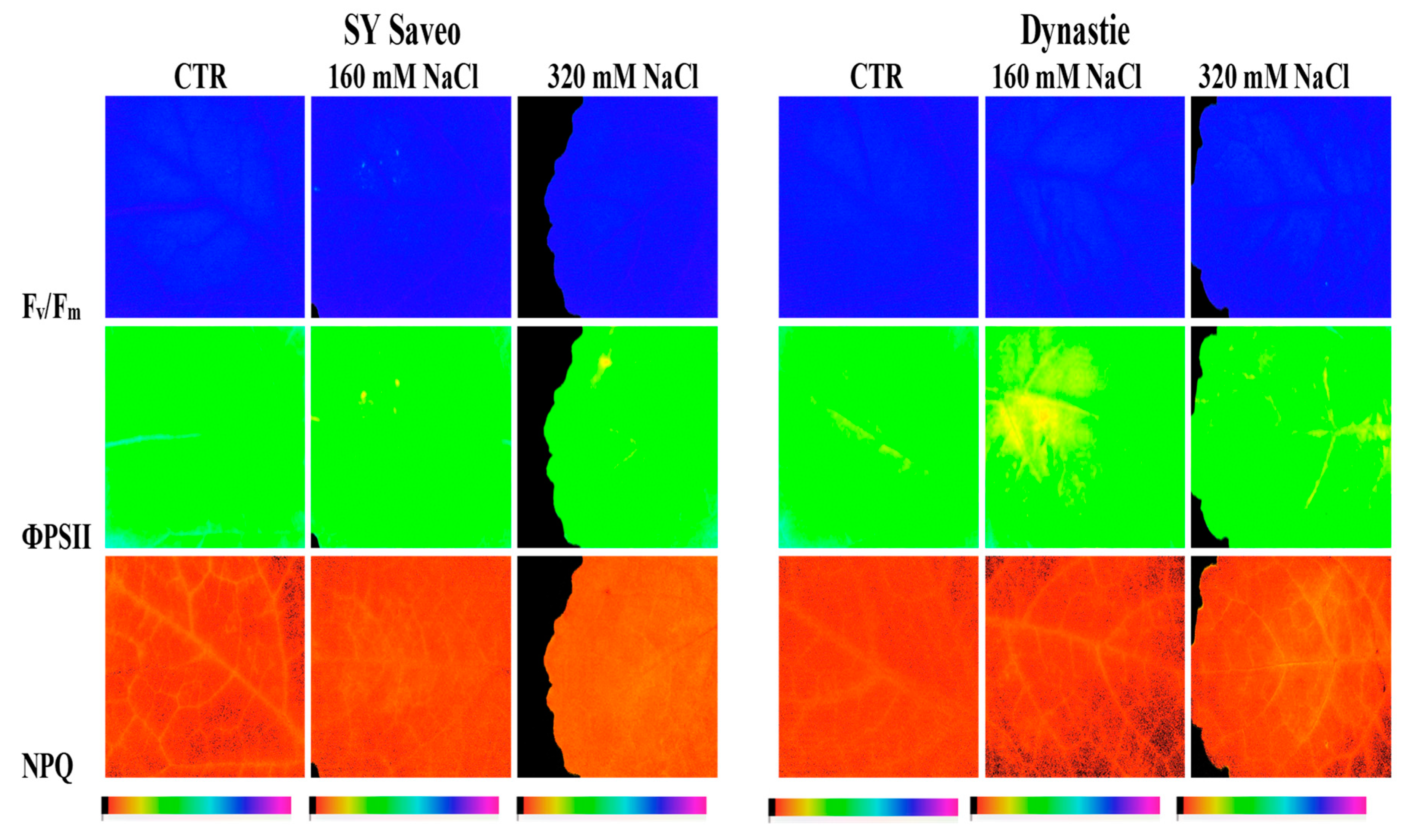
2.7. Imaging of Chlorophyll Fluorescence and Leaf Total Chlorophyll Content
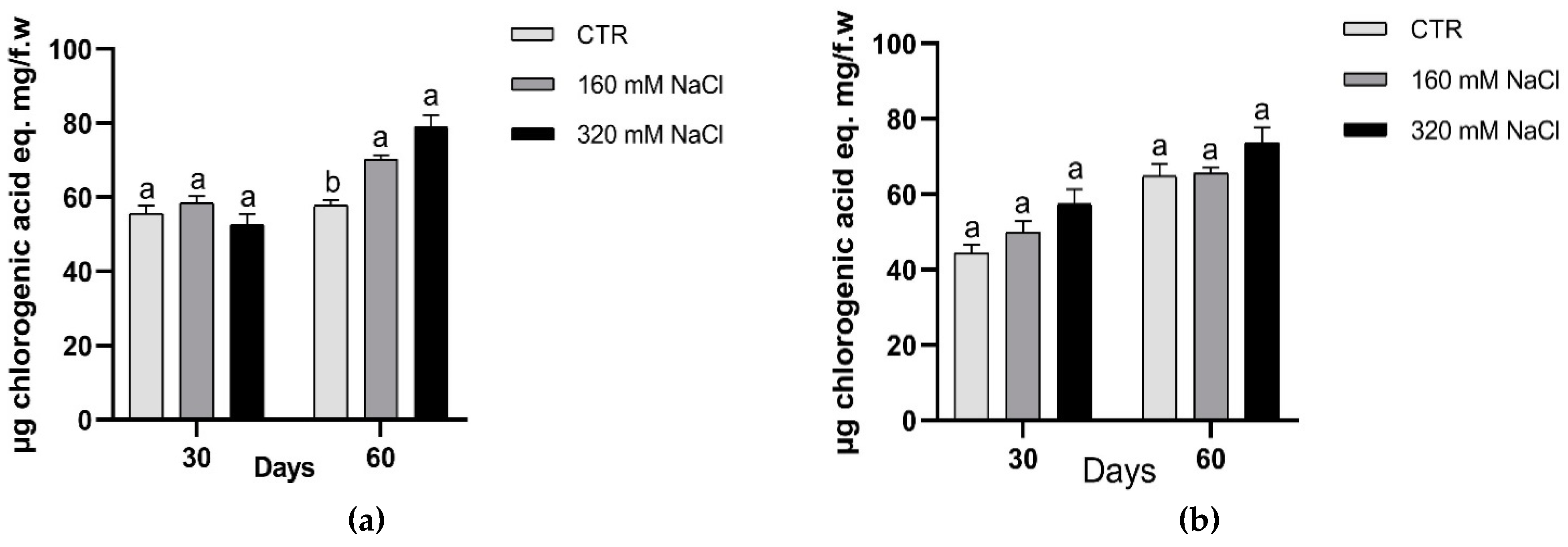
2.8. Phenolic Compounds
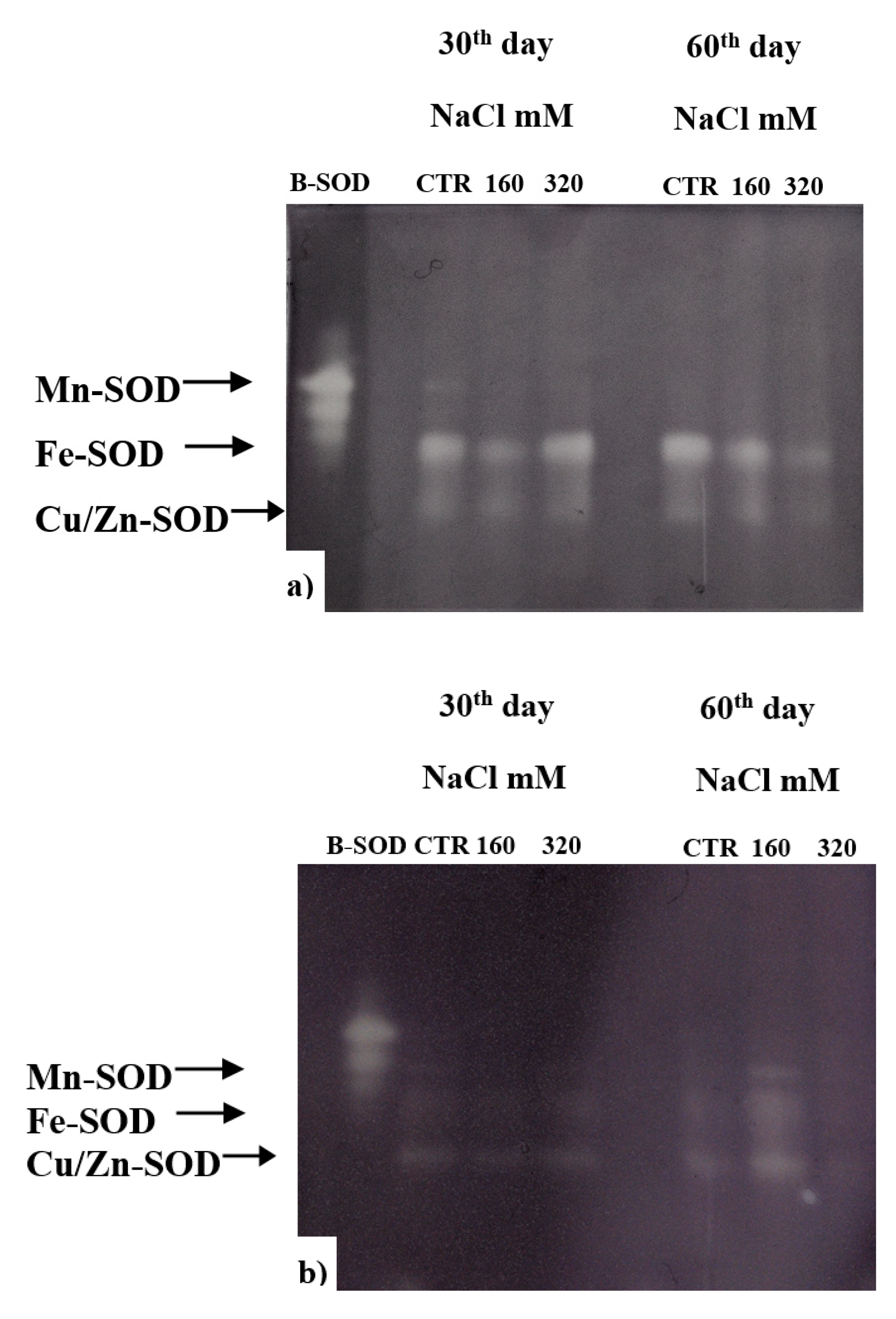
2.9. Determination of Enzymatic Activities
2.10. Polyamines
2.11. Proline
2.12. Statistical Analysis
3. Results
3.1. Effects of Soil Salinization On Plants
3.2. Plant Response to Salt Stress
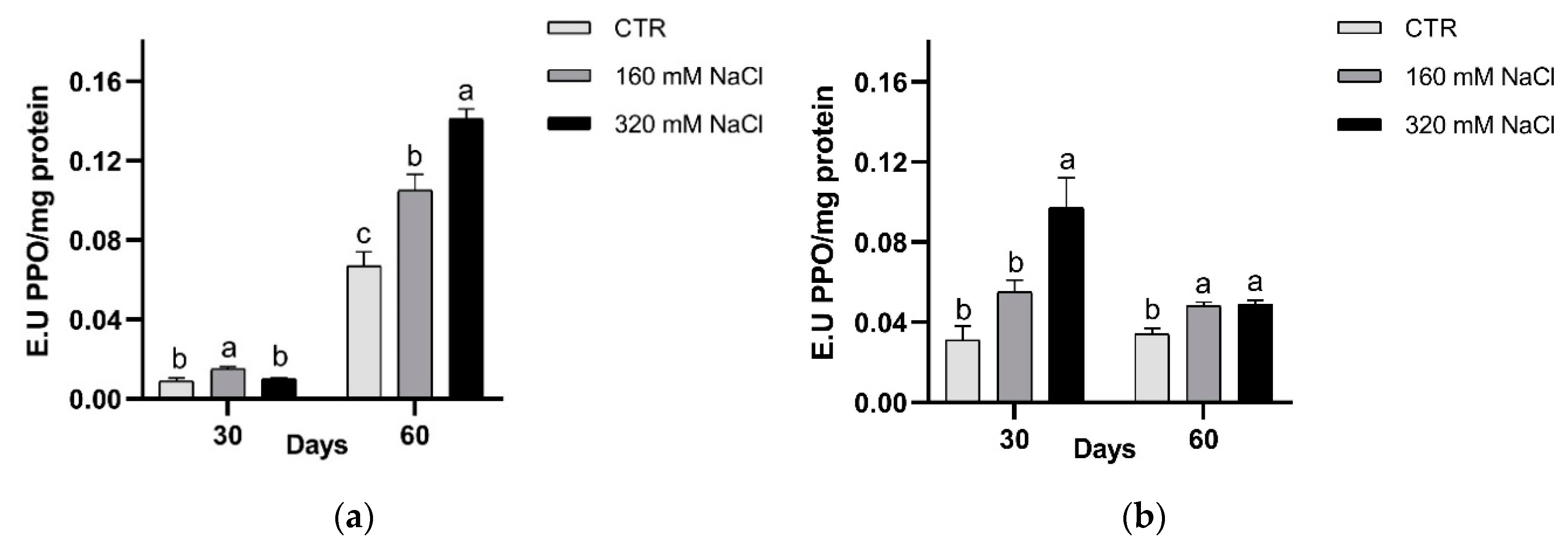
3.2.1. Antioxidant Defence
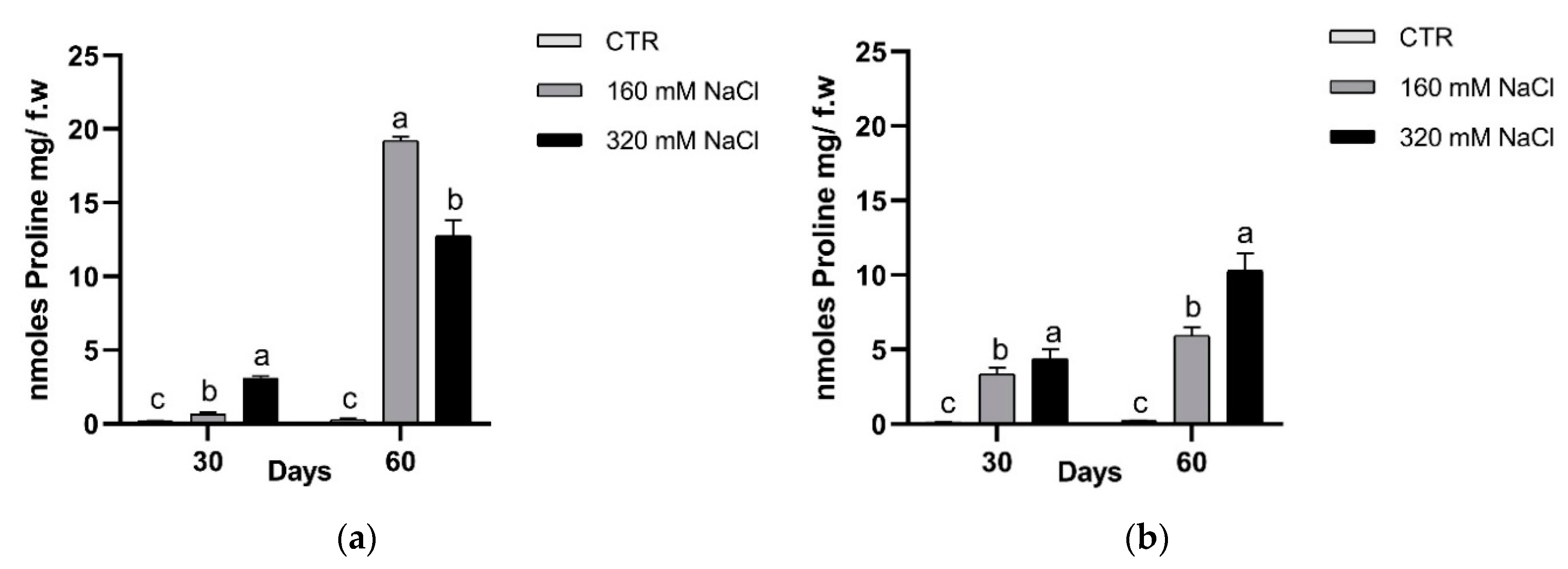
3.2.2. Osmotic Balancing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| APX | ascorbate peroxidase |
| PA | polyamines |
| SOD | superoxide dismutase |
| ROS | reactive oxygen species |
| SPM | spermine |
| SPD | spermidine |
| PUT | putrescine |
References
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef]
- Canfora, L.; Salvati, L.; Benedetti, A.; Dazzi, C.; Lo Papa, G. Saline soils in Italy: Distribution, ecological processes and socioeconomic issues. Riv. Econ. Agrar. 2017, 1, 63–77. [Google Scholar]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef]
- Forni, C.; Duca, D.; Glick, B.R. Mechanisms of plant response to salt and drought stress and their alteration by rhizobacteria. Plant Soil 2017, 410, 335–356. [Google Scholar] [CrossRef]
- Flexas, J.; Diaz-Espejo, A.; Galmés, J.; Kaldenhoff, R.; Medrano, H.; Ribas-Carbo, M. Rapid variations of mesophyll conductance in response to changes in CO2 concentration around leaves. Plant Cell Environ. 2007, 30, 1284–1298. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Bartels, D.; Sunkar, R. Drought and salt tolerance in plants. Crit. Rev. Plant Sci. 2005, 24, 23–28. [Google Scholar] [CrossRef]
- Wanasundara, J.P.D. Proteins of Brassicaceae oilseeds and their potential as a plant protein source. Crit. Rev. Food Sci. Nutr. 2011, 51, 635–677. [Google Scholar] [CrossRef]
- Shokri-Gharelo, R.; Noparvar, P.M. Molecular response of canola to salt stress: Insights on tolerance mechanisms. Peer J. 2018, 6, e4822. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Potential biochemical indicators of salinity tolerance in plants. Plant Sci. 2004, 166, 3–16. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Calif. Agric. Exp. Stn. Circ. 1950, 347, 1–32. [Google Scholar]
- Zeng, F.; Shabala, L.; Zhou, M.; Zhang, G.; Shabala, S. Barley responses to combined waterlogging and salinity stress: Separating effects of oxygen deprivation and elemental toxicity. Front. Plant Sci. 2013, 4, 313. [Google Scholar] [CrossRef]
- Blum, A.; Ebercon, A. Cell membrane stability as a measure of drought and heat tolerance in wheat. Crop Sci. 1981, 21, 43–47. [Google Scholar] [CrossRef]
- Merchant, A.; Tausz, M.; Arndt, S.K.; Adams, M.A. Cyclitols and carbohydrates in leaves and roots of 13 Eucalyptus species suggest contrasting physiological responses to water deficit. Plant Cell Environ. 2006, 29, 2017–2029. [Google Scholar] [CrossRef]
- Sairam, R.K.; Deshmukh, P.S.; Shukla, D.S. Tolerance of drought and temperature stress in relation to increased antioxidant enzyme activity in wheat. J. Agron. Crop Sci. 1997, 178, 171–178. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Bilger, W.; Björkman, O. Temperature dependence of violaxanthin de-epoxidation and non-photochemical fluorescence quenching in intact leaves of Gossypium hirsutum L. and Malva parviflora L. Planta 1991, 184, 226–234. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence-a practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Cerovic, Z.G.; Masdoumier, G.; Ghozlen, N.B.; Latouche, G. A new optical leaf-clip meter for simultaneous non-destructive assessment of leaf chlorophyll and epidermal flavonoids. Physiol. Plant 2012, 146, 251–260. [Google Scholar] [CrossRef]
- Orzali, L.; Forni, C.; Riccioni, L. Effect of chitosan seed treatment as elicitor of resistance to Fusarium graminearum in wheat. Seed Sci. Technol. 2014, 42, 132–149. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Mittler, R.; Zilinskas, B.A. Detection of ascorbate peroxidase activity in Native Gels by inhibition of the ascorbate-dependent reduction of nitroblue tetrazolium. Anal. Biochem. 1993, 212, 540–546. [Google Scholar] [CrossRef]
- Beninati, S.; Martinet, N.; Folk, J.E. High-performance liquid chromatographic method for the determination of ε-(γ-glutamyl) lysine and mono- and bis-γ-glutamyl derivatives of putrescine and spermidine. J. Chromatogr. A 1988, 443, 329–335. [Google Scholar] [CrossRef]
- Carillo, P.; Gibon, Y. PROTOCOL: Extraction and Determination of Proline. PrometheusWiki 2011. Available online: http://prometheuswiki.publish.csiro.au/tiki (accessed on 10 January 2018).
- Yamaguchi, T.; Blumwald, E. Developing salt-tolerant crop plants: Challenges and opportunities. Trends Plant Sci. 2005, 10, 615–620. [Google Scholar] [CrossRef]
- Khan, M.H.; Panda, S.K. Alterations in root lipid peroxidation and antioxidative responses in two rice cultivars under NaCl-salinity stress. Acta Physiol. Plant. 2008, 30, 81–89. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Ann. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Kapoor, K.; Srivastava, A. Assessment of salinity tolerance of Vinga mungo var. Pu-19 using ex vitro and in vitro methods. Asian J. Biotechnol. 2010, 2, 73–85. [Google Scholar]
- Ashraf, M.Y.; Akhtar, K.; Sarwar, G.; Ashraf, M. Role of rooting system in salt tolerance potential of different guar accessions. Agron. Sustain. Dev. 2005, 25, 243–249. [Google Scholar] [CrossRef]
- Khan, M.A.; Shirazi, M.U.; Khan, M.A.; Mujtaba, S.M.; Islam, E.; Mumtaz, S.; Shereen, A.; Ansari, R.U.; Ashraf, M.Y. Role of proline, K+/Na+ ratio and chlorophyll content in salt tolerance of wheat (Triticum aestivum L.). Pak. J. Bot. 2009, 41, 633–638. [Google Scholar]
- Panda, D.; Dash, P.K.; Dhal, N.K.; Rout, N.C. Chlorophyll fluorescence parameters and chlorophyll content in mangrove species grown in different salinity. Gen. Appl. Plant Physiol. 2006, 32, 175–180. [Google Scholar]
- Guidi, L.; Landi, M.; Penella, C.; Calatayud, A. Application of modulated chlorophyll fluorescence and modulated chlorophyll fluorescence imaging to study the environmental stresses effect. Ann. Bot. 2016, 6, 39–56. [Google Scholar]
- Kalaji, H.M.; Bosa, K.; Kościelniak, J.; Żuk-Gołaszewska, K. Effects of salt stress on photosystem II efficiency and CO2 assimilation of two Syrian barley landraces. Environ. Exp. Bot. 2011, 73, 64–72. [Google Scholar] [CrossRef]
- Singh, D.P.; Sarkar, R.K. Distinction and characterisation of salinity tolerant and sensitive rice cultivars as probed by the chlorophyll fluorescence characteristics and growth parameters. Funct. Plant Biol. 2014, 41, 727–736. [Google Scholar] [CrossRef]
- Baker, N.R.; Rosenqvist, E. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities. J. Exp. Bot. 2004, 55, 1607–1621. [Google Scholar] [CrossRef]
- Bongi, G.; Loreto, F. Gas-exchange properties of salt-stressed olive (Olea europea L.) leaves. Plant Physiol. 1989, 90, 1408–1416. [Google Scholar] [CrossRef]
- Pietrini, F.; Zacchini, M.; Iori, V.; Pietrosanti, L.; Bianconi, D.; Massacci, A. Screening of poplar clones for cadmium phytoremediation using photosynthesis, biomass and cadmium content analyses. Int. J. Phytorem. 2010, 12, 105–120. [Google Scholar] [CrossRef]
- Mahlooji, M.; Sharifi, R.S.; Razmjoo, J.; Sabzalian, M.R.; Sedghi, M. Effect of salt stress on photosynthesis and physiological parameters of three contrasting barley genotypes. Photosynthetica 2018, 56, 549–556. [Google Scholar] [CrossRef]
- Acosta-Motos, J.; Ortuño, M.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.; Hernandez, J. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Schreiber, U.; Bilger, W.; Neubauer, C. Chlorophyll fluorescence as a nonintrusive indicator for rapid assessment of in vivo photosynthesis. In Ecophysiology of Photosynthesis; Schulze, E.D., Caldwell, M.M., Eds.; Springer: Berlin, Germany, 1994; pp. 49–70. [Google Scholar]
- Shangguan, Z.; Shao, M.; Dyckmans, J. Effects of nitrogen nutrition and water deficit on net photosynthetic rate and chlorophyll fluorescence in winter wheat. J. Plant Physiol. 2000, 156, 46–51. [Google Scholar] [CrossRef]
- Epron, D.; Dreyer, E.; Bréda, N. Photosynthesis of oak trees [Quercus petraea (Matt.) Liebl.] during drought under field conditions: Diurnal courses of net CO2 assimilation and photochemical efficiency of photosystem II. Plant Cell Environ. 1992, 15, 809–820. [Google Scholar] [CrossRef]
- Stepien, P.; Johnson, G.N. Contrasting responses of photosynthesis to salt stress in the glycophyte Arabidopsis and the halophyte Thellungiella: Role of the plastid terminal oxidase as an alternative electron sink. Plant Physiol. 2009, 149, 1154–1165. [Google Scholar] [CrossRef]
- Dai, Y.J.; Shen, Z.G.; Liu, Y.; Wang, L.L.; Hannaway, D.; Lu, H.F. Effects of shade treatments on the photosynthetic capacity, chlorophyll fluorescence, and chlorophyll content of Tetrastigma hemsleyanum Diels et Gilg. Environ. Exp. Bot. 2009, 65, 177–182. [Google Scholar] [CrossRef]
- Jamil, M.; Lee, K.J.; Kim, J.M.; Kim, H.S.; Rha, E.S. Salinity reduced growth PS2 photochemistry and chlorophyll content in radish. Sci. Agric. 2007, 64, 111–118. [Google Scholar] [CrossRef]
- Ayyub, C.; Rashid Shaheen, M.; Raza, S.; Sarwar Yaqoob, M.; Khan Qadri, R.; Azam, M.; Ghani, M.; Khan, I.; Akhtar, N. Evaluation of different radish (Raphanus sativus) genotypes under different saline regimes. Am. J. Plant Sci. 2016, 7, 894–898. [Google Scholar] [CrossRef]
- Lin, Z.F.; Ehleringer, J.R. Effects of leaf age on photosynthesis and water use efficiency of papaya. Photosynthetica 1982, 16, 514–519. [Google Scholar]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic compounds in Brassica vegetables. Molecules 2011, 16, 251–280. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Sofo, A.; Scopa, A.; Nuzzaci, M.; Vitti, A. Ascorbate peroxidase and catalase activities and their genetic regulation in plants subjected to drought and salinity stresses. Int. J. Mol. Sci. 2015, 16, 13561–13578. [Google Scholar] [CrossRef]
- Eyidogan, F.; Oz, M.T. Effect of salinity on antioxidant responses of chickpea seedlings. Acta Physiol. Plant. 2005, 29, 485–493. [Google Scholar] [CrossRef]
- Aziz, A.; Martin-Tanguy, J.; Larher, F. Plasticity of polyamine metabolism associated with high osmotic stress in rape leaf discs and with ethylene treatment. Plant Growth Regul. 1997, 21, 153–163. [Google Scholar] [CrossRef]
- Takahashi, T.; Kakehi, J.I. Polyamines: Ubiquitous polycations with unique roles in growth and stress responses. Ann. Bot. 2010, 105, 1–6. [Google Scholar] [CrossRef]
- Janicka-Russak, M.; Kabała, K.; Młodzińska, E.; Kłobus, G. The role of polyamines in the regulation of the plasma membrane and the tonoplast proton pumps under salt stress. J. Plant Physiol. 2010, 167, 261–269. [Google Scholar] [CrossRef]
- Zapata, P.J.; Serrano, M.; Pretel, M.T.; Amoros, A.; Botella, M.A. Polyamines and ethylene changes during germination of different plant species under salinity. Plant Sci. 2004, 167, 781–788. [Google Scholar] [CrossRef]
- Quinet, M.; Ndayiragije, A.; Lefèvre, I.; Lambillotte, B.; Dupont-Gillain, C.C.; Lutts, S. Putrescine differently influences the effect of salt stress on polyamine metabolism and ethylene synthesis in rice cultivars differing in salt resistance. J. Exp. Bot. 2010, 61, 2719–2733. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Biomass | ||||
|---|---|---|---|---|
| Dynastie | SY Saveo | |||
| NaCl | 30 d | 60 d | 30 d | 60 d |
| CTR | 1.02 ± 0.03 a | 4.31 ± 0.03 a | 1.00 ± 0.05 a | 3.27 ± 0.22 a |
| 160 mM | 0.94 ± 0.04 a | 1.94 ± 0.08 b | 0.76 ± 0.03 b | 1.86 ± 0.07 b |
| 320 mM | 0.74 ± 0.04 b | 1.05 ± 0.02 c | 0.66 ± 0.02 c | 0.96 ± 0.05 c |
| Dry Weight/Fresh Weight | Soluble Proteins (µg Protein/mg f.w.) | |||||
|---|---|---|---|---|---|---|
| Dynastie | SY Saveo | Dynastie | SY Saveo | |||
| NaCl | 60 d | 60 d | 30 d | 60 d | 30 d | 60 d |
| CTR | 0.018 ± 0.0005 c | 0.069 ± 0.001 b | 4.62 ± 0.06 b | 7.30 ± 0.12 a | 3.21 ± 0.14 c | 4.32 ± 0.14 b |
| 160 mM | 0.053 ± 0.003 b | 0.077 ± 0.008 ab | 4.62 ± 0.06 b | 5.50 ± 0.09 b | 4.40 ± 0.07 a | 4.69 ± 0.07 b |
| 320 mM | 0.083 ± 0.0005 a | 0.086 ± 0.005 a | 7.66 ± 0.08 a | 7.82 ± 0.16 a | 3.75 ± 0.07 b | 8.98 ± 0.15 a |
| Plants Water Content (%) | ||
|---|---|---|
| NaCl | Dynastie | SY Saveo |
| CTR | 92.37 ± 0.12 a | 93.10 ± 0.12 a |
| 160 mM | 91.67 ± 0.50 a | 91.39 ± 0.17 b |
| 320 mM | 89.38 ± 0.07 b | 90.82 ± 0.10 c |
| Cvs. | NaCl | Fv/Fm (r.u.) | ΦPSII (r.u.) | NPQ (r.u.) | ETR (μmol Electrons m−2s−1) | Abs (rel. un.) | Tot Chl Content (µg cm−2) |
|---|---|---|---|---|---|---|---|
| SY Saveo | CTR | 0.810 ± 0.001 a | 0.371 ± 0.013 ab | 0.257 ± 0.005 b | 8.10 ± 0.28 ab | 0.804 ±0.003 d | 27.64 ± 0.63 a |
| 160 mM | 0.814 ± 0.001 a | 0.349 ± 0.009 bc | 0.278 ± 0.010 b | 7.98 ± 0.21 ab | 0.846 ± 0.002 c | 37.22 ± 0.06 b | |
| 320 mM | 0.812 ± 0.001 a | 0.334 ± 0.002 cd | 0.333 ± 0.010 a | 7.74 ± 0.05 bc | 0.859 ± 0.002 ab | 41.26 ± 0.75 c | |
| Dynastie | CTR | 0.805 ± 0.001 b | 0.385 ± 0.002 a | 0.241 ± 0.010 b | 8.35 ± 0.04 a | 0.806 ± 0.004 d | 28.15 ± 0.90 a |
| 160 mM | 0.804 ± 0.003 b | 0.316 ± 0.002 d | 0.198 ± 0.019 c | 7.33 ± 0.06 c | 0.848 ± 0.005 bc | 37.93 ± 1.66 b | |
| 320 mM | 0.805 ± 0.001 b | 0.320 ± 0.005 d | 0.277 ± 0.017 b | 7.43 ± 0.12 c | 0.860 ± 0.001 a | 41.38 ± 0.30 c | |
| p-Value (ANOVA) | Cv. | *** | ns | *** | ns | ns | ns |
| Treat. | ns | *** | *** | ** | *** | *** | |
| Cv.x Treat. | ns | * | ns | * | ns | ns |
| Polyamines (nmol/mg f.w.) | ||||||
|---|---|---|---|---|---|---|
| 30 days | 60 days | |||||
| NaCl | SPM | SPD | PUT | SPM | SPD | PUT |
| CTR | 18.37± 1.92 b | 100.71 ± 4.34 a | 19.74 ± 1.31 | 27.29 ± 7.60 a | 62.01 ± 4.61 b | 14.08 ± 2.30 b |
| 160 mM | 29.20 ± 1.53 a | 67.04 ± 1.83 b | N.D.* | 24.83 ± 1.93 a | 65.25 ± 3.69 b | 39.66 ± 3.92 a |
| 320 mM | 22.76 ± 2.65 b | 68.33 ± 2.31 b | N.D.* | 30.76 ± 3.54 a | 166.78 ± 12.18 a | 35.78 ± 3.11 a |
| Polyamines (nmol/mg f.w.) | ||||||
|---|---|---|---|---|---|---|
| 30 days | 60 days | |||||
| NaCl | SPM | SPD | PUT | SPM | SPD | PUT |
| CTR | 32.70 ± 1.98 b | 97.83 ± 4.78 b | 18.17 ± 1.54 a | 13.89 ± 0.31 b | 46.65 ± 0.34 c | 12.58 ± 2.27 a |
| 160 mM | 44.70 ± 2.67 a | 92.32 ± 4.76 b | 12.63 ± 1.08 b | 44.31 ± 7.29 a | 99.83 ± 4.90 b | 23.87 ± 4.65 a |
| 320 mM | 37.57 ± 2.71 ab | 150.81 ± 9.97 a | 22.76 ± 2.65 a | 40.55 ± 7.81 a | 151.08 ± 11.10 a | 19.73 ± 5.62 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santangeli, M.; Capo, C.; Beninati, S.; Pietrini, F.; Forni, C. Gradual Exposure to Salinity Improves Tolerance to Salt Stress in Rapeseed (Brassica napus L.). Water 2019, 11, 1667. https://doi.org/10.3390/w11081667
Santangeli M, Capo C, Beninati S, Pietrini F, Forni C. Gradual Exposure to Salinity Improves Tolerance to Salt Stress in Rapeseed (Brassica napus L.). Water. 2019; 11(8):1667. https://doi.org/10.3390/w11081667
Chicago/Turabian StyleSantangeli, Michael, Concetta Capo, Simone Beninati, Fabrizio Pietrini, and Cinzia Forni. 2019. "Gradual Exposure to Salinity Improves Tolerance to Salt Stress in Rapeseed (Brassica napus L.)" Water 11, no. 8: 1667. https://doi.org/10.3390/w11081667
APA StyleSantangeli, M., Capo, C., Beninati, S., Pietrini, F., & Forni, C. (2019). Gradual Exposure to Salinity Improves Tolerance to Salt Stress in Rapeseed (Brassica napus L.). Water, 11(8), 1667. https://doi.org/10.3390/w11081667