Feces from Piscivorous and Herbivorous Birds Stimulate Differentially Phytoplankton Growth

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
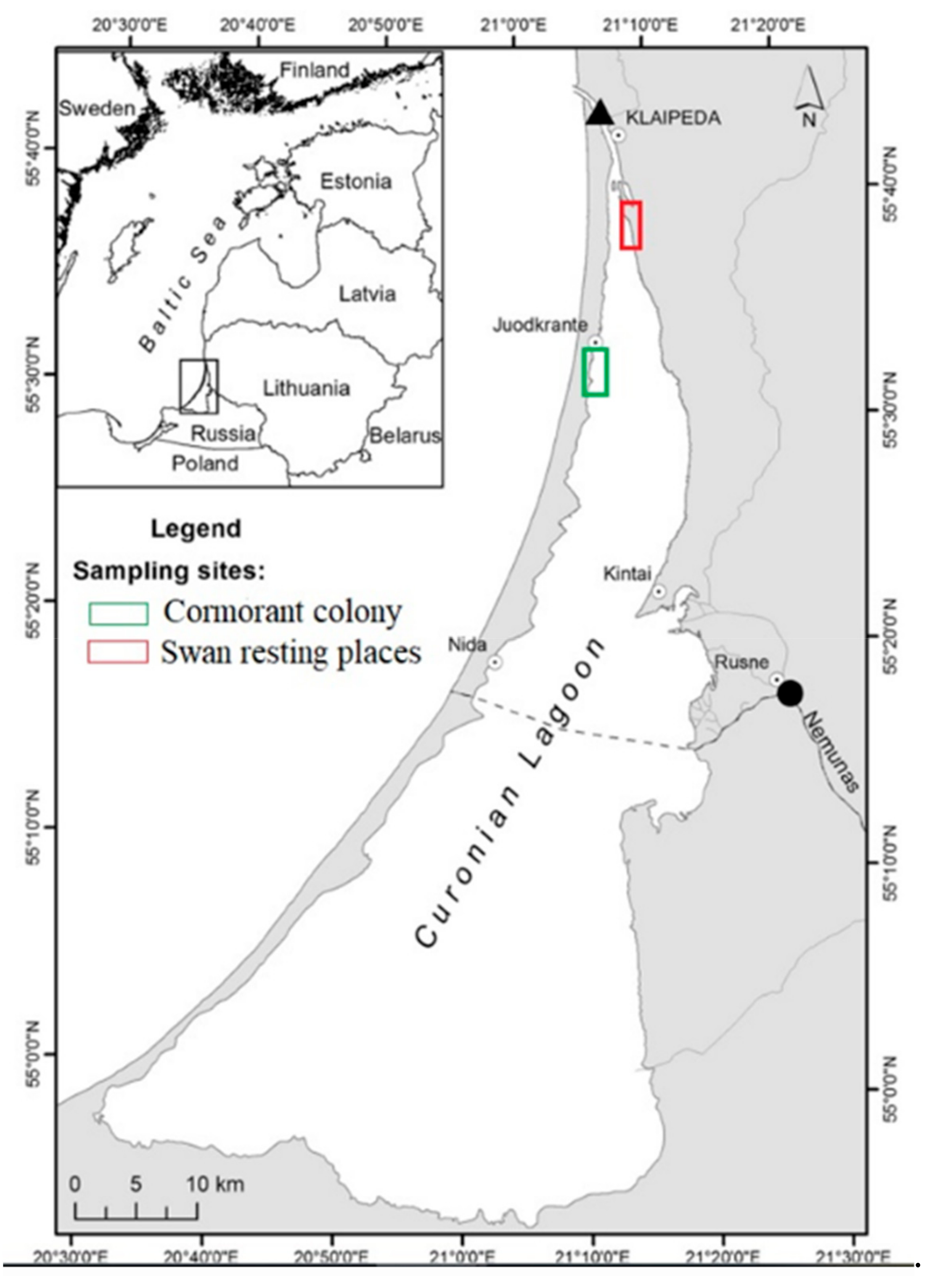
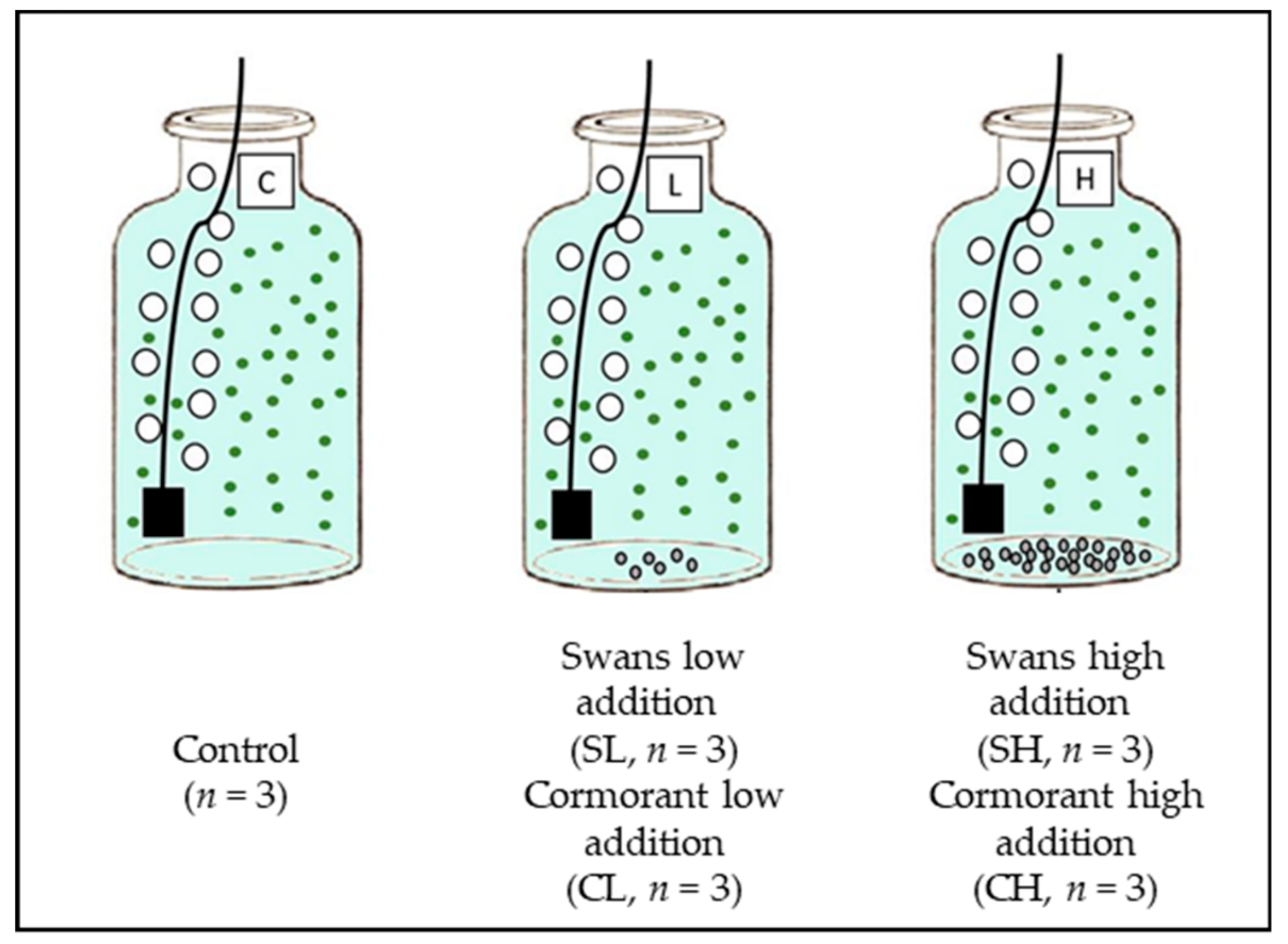
2.1. Sampling Procedure and Microcosms Set Up
2.2. Nutrients, Phytoplankton and Heterotrophic Bacteria Analysis
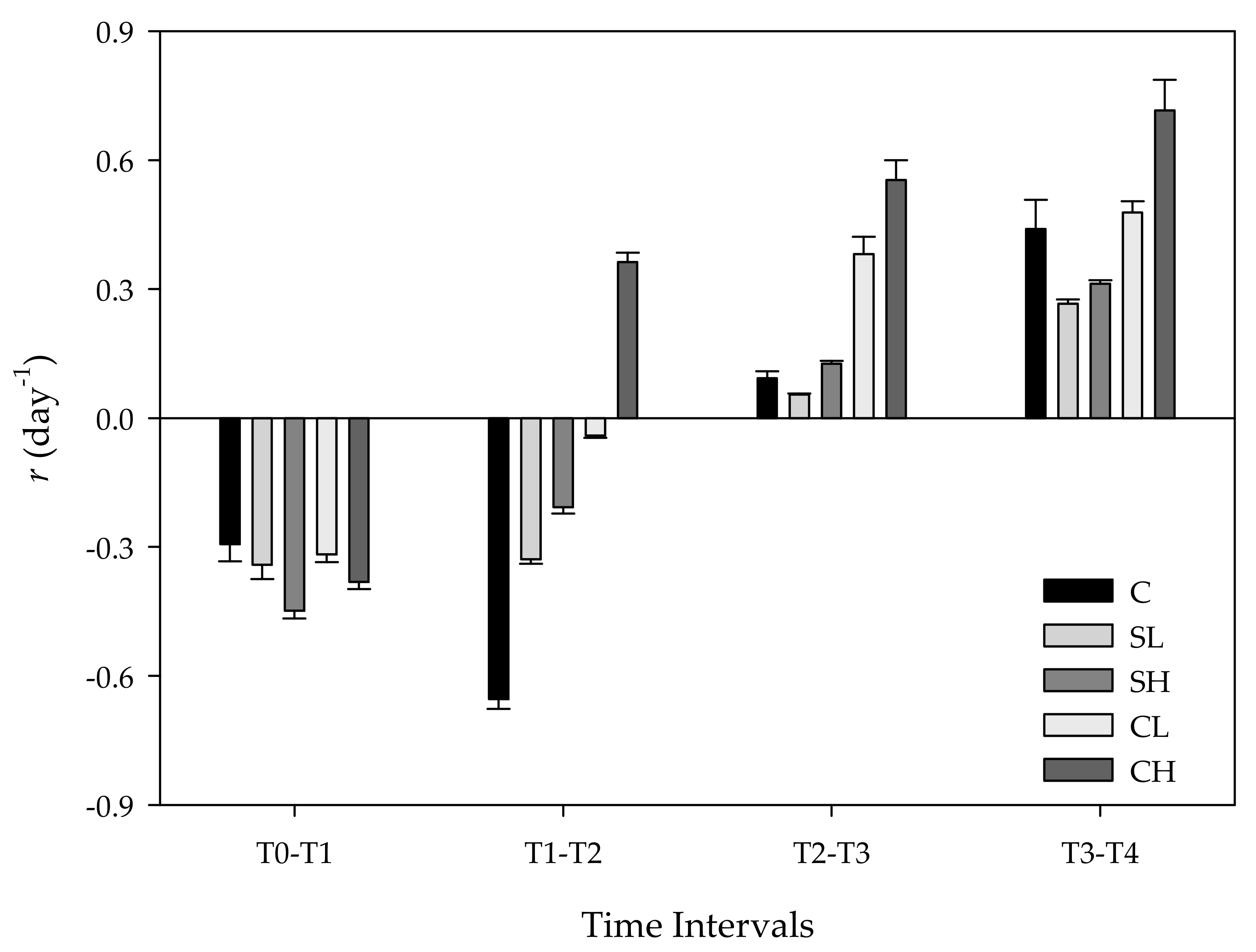
3. Results
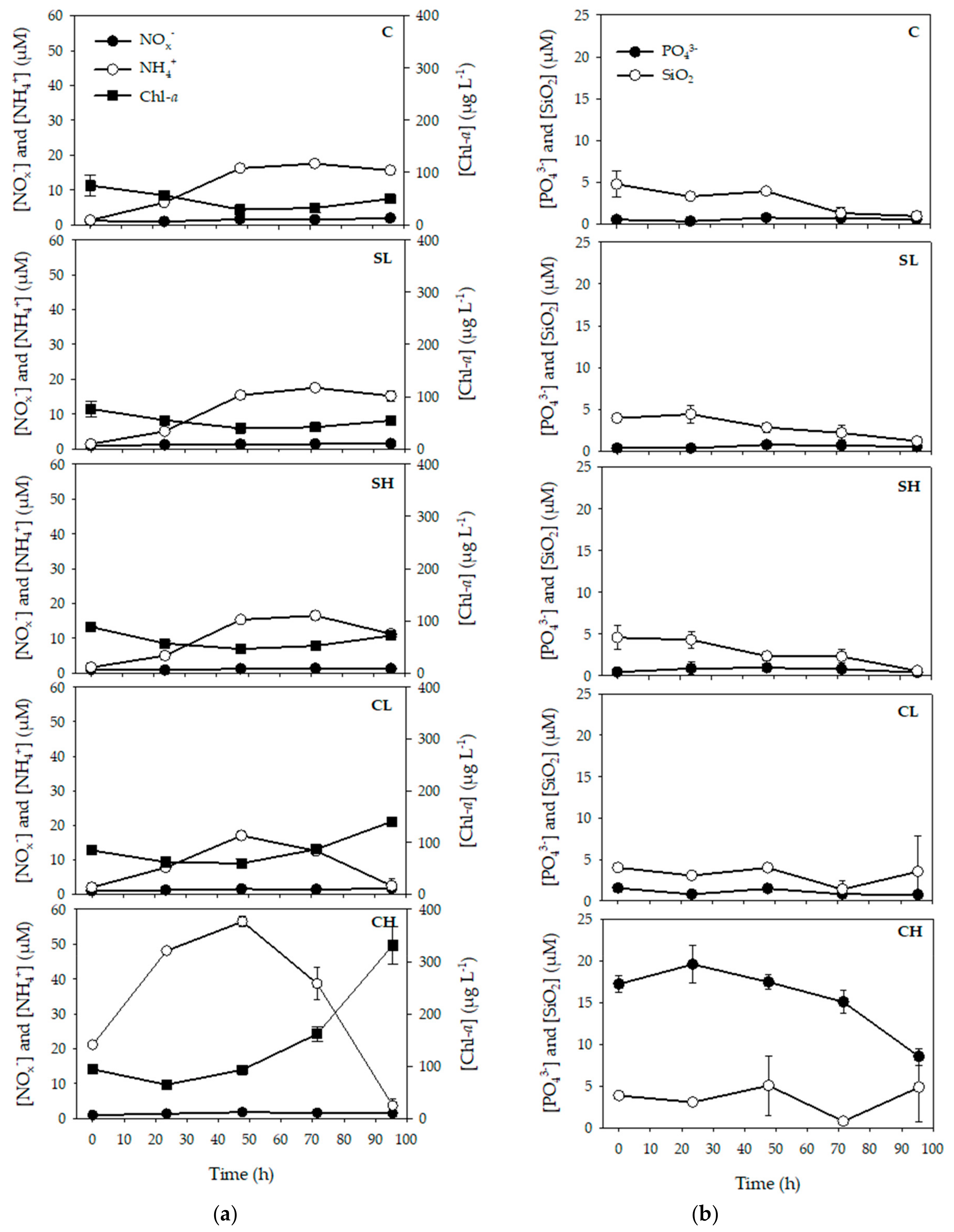
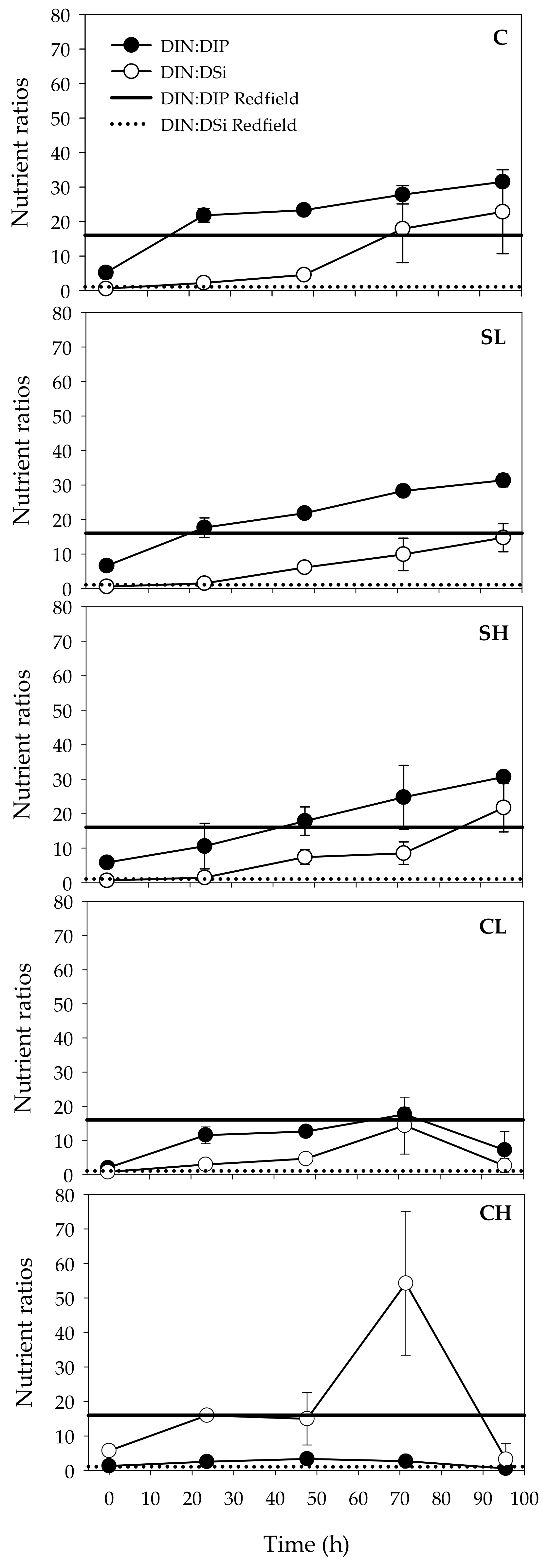
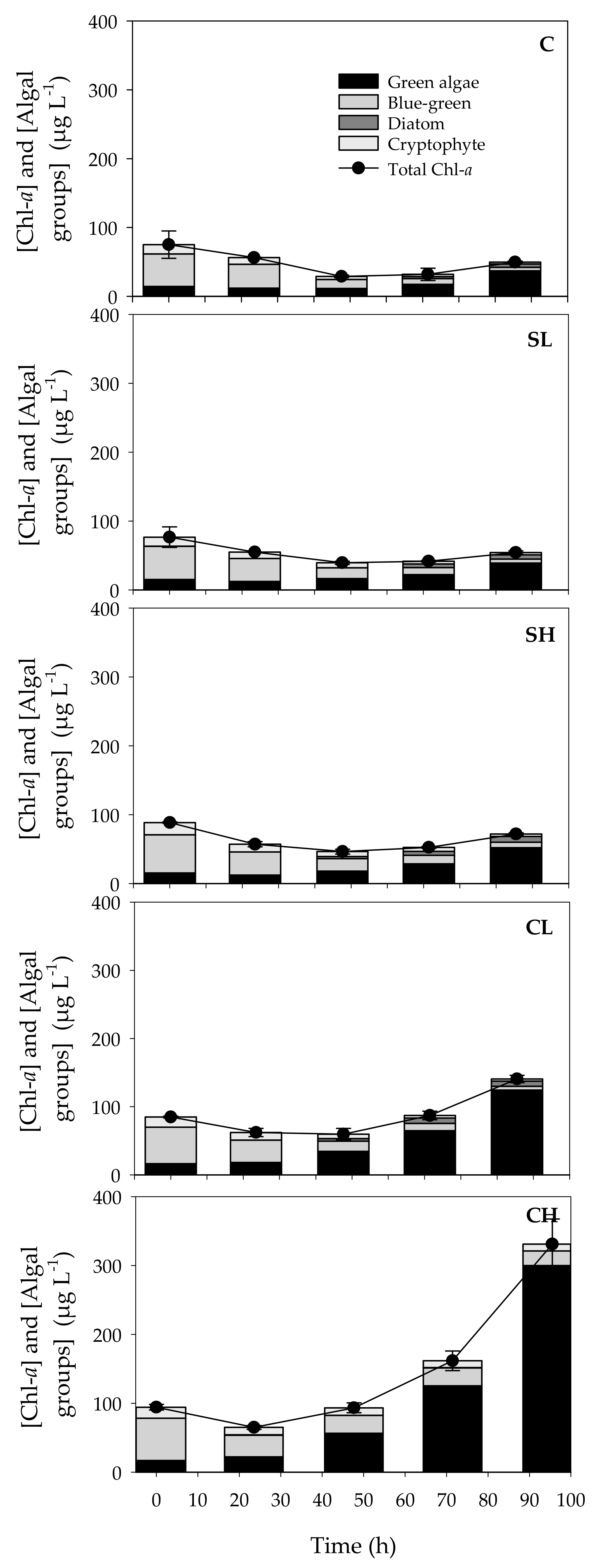
3.1. Effects of Bird Feces Addition on Inorganic Nutrients and Chl-a Concentrations
3.2. Phytoplankton Community Changes
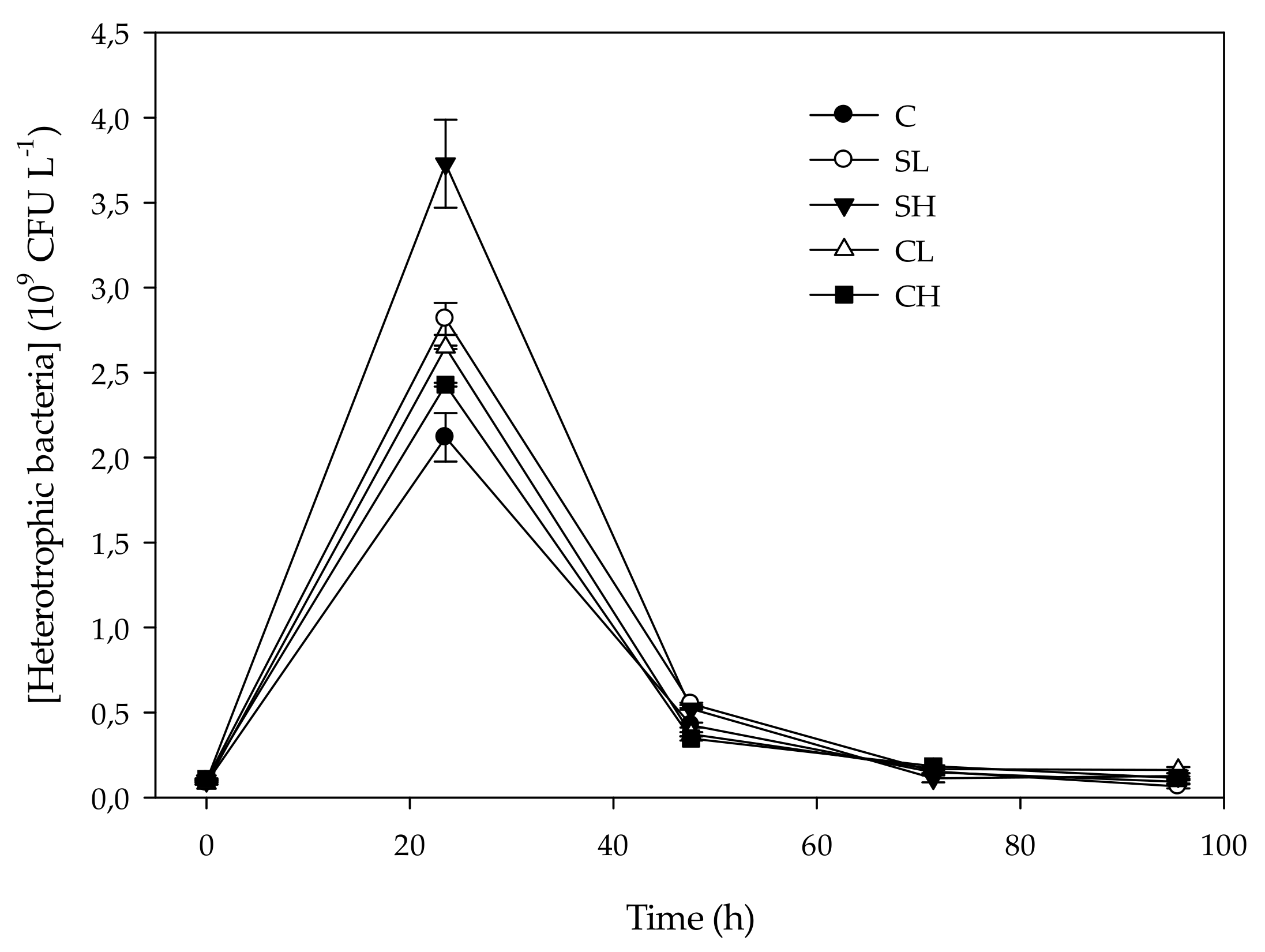
3.3. Heterotrophic Bacteria Abundance
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Viaroli, P.; Soana, E.; Pecora, S.; Laini, A.; Naldi, M.; Fano, E.A.; Nizzoli, D. Space and time variations of watershed N and P budgets and their relationships with reactive N and P loadings in a heavily impacted river basin (Po river, Northern Italy). Sci. Total Environ. 2018, 639, 1574–1587. [Google Scholar] [CrossRef]
- Vybernaite-Lubiene, I.; Zilius, M.; Saltyte-Vaisiauske, L.; Bartoli, M. Recent trends (2012–2016) of N, Si, and P export from the Nemunas River Watershed: Loads, unbalanced stoichiometry, and threats for downstream aquatic ecosystems. Water 2018, 10, 1178. [Google Scholar] [CrossRef]
- Powell, G.V.; Fourqurean, J.W.; Kenworthy, W.J.; Zieman, J.C. Bird colonies cause seagrass enrichment in a subtropical estuary: Observational and experimental evidence. Estuar. Coast. Shelf Sci. 1991, 32, 567–579. [Google Scholar] [CrossRef]
- Søndergaard, M.; Jensen, J.P.; Jeppesen, E. Role of sediment and internal loading of phosphorus in shallow lakes. Hydrobiologia 2003, 506, 135–145. [Google Scholar] [CrossRef]
- Orlova, M.; Golubkov, S.; Kalinina, L.; Ignatieva, N. Dreissena polymorpha (Bivalvia: Dreissenidae) in the Neva Estuary (eastern Gulf of Finland, Baltic Sea): Is it a biofilter or source for pollution? Mar. Pollut. Bull. 2004, 49, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Petkuviene, J.; Zilius, M.; Lubiene, I.; Ruginis, T.; Giordani, G.; Razinkovas-Baziukas, A.; Bartoli, M. Phosphorus cycling in a freshwater estuary impacted by cyanobacterial blooms. Estuar. Coasts 2016, 39, 1386–1402. [Google Scholar] [CrossRef]
- Klimaszyk, P.; Rzymski, P.; Piotrowicz, R.; Joniak, T. Contribution of surface runoff from forested areas to the chemistry of a through-flow lake. Environ. Earth Sci. 2015, 73, 3963–3973. [Google Scholar] [CrossRef]
- Klimaszyk, P.; Rzymski, P. The complexity of ecological impacts induced by great cormorants. Hydrobiologia 2016, 771, 13–30. [Google Scholar] [CrossRef]
- Klimaszyk, P.; Brzeg, A.; Rzymski, P.; Piotrowicz, R. Black spots for aquatic and terrestrial ecosystems: Impact of a perennial cormorant colony on the environment. Sci. Total Environ. 2015, 517, 222–231. [Google Scholar] [CrossRef]
- Purcell, S.L. The Significance of Waterfowl Feces as a Source of Nutrients to Algae in a Prairie Wetland. Ph.D. Thesis, University of Manitoba, Winnipeg, MB, Canada, 1999. [Google Scholar]
- Gwiazda, R.; Jarocha, K.; Szarek-Gwiazda, E. Impact of a small cormorant (Phalacrocorax carbo sinensis) roost on nutrients and phytoplankton assemblages in the littoral regions of a submontane reservoir. Biologia 2010, 65, 742–748. [Google Scholar] [CrossRef]
- Jauffrais, T.; Drouet, S.; Turpin, V.; Méléder, V.; Jesus, B.; Cognie, B.; Raimbault, P.; Cosson, R.P.; Decottignies, P.; Martin-Jézéquel, V. Growth and biochemical composition of a microphytobenthic diatom (Entomoneis paludosa) exposed to shorebird (Calidris alpina) droppings. J. Exp. Mar. Biol. Ecol. 2015, 469, 83–92. [Google Scholar] [CrossRef]
- Klimaszyk, P.; Piotrowicz, R.; Rzymski, P. Changes in physico-chemical conditions and macrophyte abundance in a shallow soft-water lake mediated by a Great Cormorant roosting colony. J. Limnol. 2014, 74. [Google Scholar] [CrossRef]
- Han, I.; Yoo, K.; Wee, G.N.; No, J.H.; Park, J.; Min, S.J.; Kim, S.H.; Leea, T.K. Short-term effects of great cormorant droppings on water quality and microbial community of an artificial agricultural reservoir. J. Environ. Qual. 2017, 46, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Scherer, N.M.; Gibbons, H.L.; Stoops, K.B.; Muller, M. Phosphorus loading of an urban lake by bird droppings. Lake Reserv. Manag. 1995, 11, 317–327. [Google Scholar] [CrossRef]
- Hahn, S.; Bauer, S.; Klaassen, M. Estimating the contribution of carnivorous waterbirds to nutrient loading in freshwater habitats. Freshw. Biol. 2007, 52, 2421–2433. [Google Scholar] [CrossRef]
- Hahn, S.; Bauer, S.; Klaassen, M. Quantification of allochthonous nutrient input into freshwater bodies by herbivorous waterbirds. Freshw. Biol. 2008, 53, 181–193. [Google Scholar] [CrossRef]
- Sitters, J.; Bakker, E.S.; Veldhuis, M.P.; Veen, G.F.; Olde Venterink, H.; Vanni, M.J. The stoichiometry of nutrient release by terrestrial herbivores and its ecosystem consequences. Front. Earth Sci. 2017, 5, 32. [Google Scholar] [CrossRef]
- Bartoli, M.; Zilius, M.; Bresciani, M.; Vaičiūtė, D.; Lubiene-Vybernaite, I.; Petkuviene, J.; Giordani, G.; Daunys, D.; Ruginis, T.; Benelli, S.; et al. Drivers of cyanobacterial blooms in a hypertrophic lagoon. Front. Mar. Sci. 2018, 5, 434. [Google Scholar] [CrossRef]
- Vybernaite-Lubiene, I.; Zilius, M.; Giordani, G.; Petkuviene, J.; Vaiciute, D.; Bukaveckas, P.A.; Bartoli, M. Effect of algal blooms on retention of N, Si and P in Europe’s largest coastal lagoon. Estuar. Coast. Shelf Sci. 2017, 194, 217–228. [Google Scholar] [CrossRef]
- Švažas, S.; Chukalova, N.; Grishanov, G.; Pūtys, Ž.; Sruoga, A.; Butkauskas, D.; Raudonikis, L.; Prakas, P. The role of Great Cormorant (Phalacrocorax carbo sinensis) for fish stock and dispersal of helminthes parasites in the Curonian Lagoon area. Vet. Med. Zoot. 2011, 55, 79–85. [Google Scholar]
- Dagys, M.; Zarankaitė, J. Status of the breeding population of Great Cormorants in Lithuania in 2012. In National Reports from the 2012 Breeding Census of Great Cormorants Phalacrocorax Carbo in Parts of the Western Palearctic. IUCN-Wetlands International Cormorant Research Group Report; Technical Report from DCE—Danish Centre for Environment and Energy; Bregnballe, T., Lynch, J., Parz-Gollner, R., Marion, L., Volponi, S., Paquet, J.-Y., van Eerden, M.R., Eds.; Aarhus University: Allus, Denmark, 2013; No. 22; pp. 69–71. Available online: http://dce2.au.dk/pub/TR22.pdf (accessed on 4 September 2019).
- Grasshoff, K.; Ehrhardt, M.; Kremling, K. Methods of Seawater Analysis, 2nd ed.; Verlag Berlin Chemie: Weinheim, Germany, 1983. [Google Scholar]
- Bower, C.E.; Holm-Hansen, T. A salicylate-hypochlorite method for determining ammonia in seawater. Can. J. Fish. Aquat. Sci. 1980, 37, 794–798. [Google Scholar] [CrossRef]
- Færøvig, P.J.; Hessen, D.O. Allocation strategies in crustacean stoichiometry: The potential role of phosphorus in the limitation of reproduction. Freshw. Biol. 2003, 48, 1782–1792. [Google Scholar] [CrossRef]
- Catherine, A.; Escoffier, N.; Belhocine, A.; Nasri, A.B.; Hamlaoui, S.; Yéprémian, C.; Bernard, C.; Troussellier, M. On the use of the FluoroProbe®, a phytoplankton quantification method based on fluorescence excitation spectra for large-scale surveys of lakes and reservoirs. Water Res. 2012, 46, 1771–1784. [Google Scholar] [CrossRef] [PubMed]
- Gould, D.J.; Fletcher, M.R. Gull droppings and their effects on water quality. Water Res. 1978, 12, 665–672. [Google Scholar] [CrossRef]
- Damare, J.M.; Hussong, D.; Weiner, R.M.; Colwell, R.R. Aerobic and facultatively anaerobic bacteria associated with the gut of Canada geese (Branta canadensis) and whistling swans (Cygnus columbianus columbianus). Appl. Environ. Microbiol. 1979, 38, 258–266. [Google Scholar]
- Alderisio, K.A.; DeLuca, N. Seasonal enumeration of fecal coliform bacteria from the feces of ring-billed gulls (Larus delawarensis) and Canada Geese (Branta canadensis). Appl. Environ. Microbiol. 1999, 65, 5628–5630. [Google Scholar]
- Lauridsen, T.L.; Jeppesen, E.; Andersen, F.Ø. Colonization of submerged macrophytes in shallow fish manipulated Lake Væng: Impact of sediment composition and waterfowl grazing. Aquat. Bot. 1993, 46, 1–15. [Google Scholar] [CrossRef]
- Valiela, I.; Alber, M.; LaMontagne, M. Fecal coliform loadings and stocks in Buttermilk Bay, Massachusetts, USA, and Management Implications. Environ. Manage. 1991, 15, 659–674. [Google Scholar] [CrossRef]
- Levesque, B.; Brousseau, P.; Simard, P.; Dewailly, E.; Meisels, M.; Ramsay, D.; Joly, J. Impact of the ring-billed gull (Larus delawarensis) on the microbiological quality of recreational water. Appl. Environ. Microbiol. 1993, 59, 1228–1230. [Google Scholar]
- Kataržytė, M.; Mėžinė, J.; Vaičiūtė, D.; Liaugaudaitė, S.; Mukauskaitė, K.; Umgiesser, G.; Schernewski, G. Fecal contamination in shallow temperate estuarine lagoon: Source of the pollution and environmental factors. Mar. Pollut. Bull. 2018, 133, 762–772. [Google Scholar] [CrossRef]
- Umgiesser, G.; Čerkasova, N.; Erturk, A.; Mėžinė, J.; Kataržytė, M. New beach in a shallow estuarine lagoon: A model-based E. coli pollution risk assessment. J. Coast. Conserv. 2018, 22, 573–586. [Google Scholar] [CrossRef]
- Manny, B.A.; Wetzel, R.G.; Johnon, W.C. Annual contribution of carbon, nitrogen and phosphorus by migrant Canada geese to hardwater lake. Ver. Internat. Verein. Limnol. 1975, 9, 949–951. [Google Scholar] [CrossRef]
- Gere, G.; Andrikovics, S. Effects of waterfowl on water quality. Hydrobiologia 1992, 243, 445–448. [Google Scholar] [CrossRef]
- Manny, B.A.; Johnson, W.C.; Wetzel, R.G. Nutrient additions by waterfowl to lakes and reservoirs: Predicting their effects on productivity and water quality. Hydrobiologia 1994, 279, 121–132. [Google Scholar] [CrossRef]
- Post, D.M.; Taylor, J.P.; Kitchell, J.F.; Olson, M.H.; Schindler, D.E.; Herwig, B.R. The role of migratory waterfowl as nutrient vectors in a managed wetland. Conserv. Biol. 1998, 12, 910–920. [Google Scholar] [CrossRef]
- Hansson, L.A.; Nicolle, A.; Brönmark, C.; Hargeby, A.; Lindström, Å.; Andersson, G. Waterfowl, macrophytes, and the clear water state of shallow lakes. Hydrobiologia 2010, 646, 101–109. [Google Scholar] [CrossRef]
- Wang, L.; Wang, C.; Deng, D.; Zhao, X.; Zhou, Z. Temporal and spatial variations in phytoplankton: Correlations with environmental factors in Shengjin Lake, China. Environ. Sci. Pollut. Res. 2015, 22, 14144–14156. [Google Scholar] [CrossRef] [PubMed]
- Cronin, G.; Lodge, D.M. Effects of light and nutrient availability on the growth, allocation, carbon/nitrogen balance, phenolic chemistry, and resistance to herbivory of two freshwater macrophytes. Oecologia 2003, 137, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Boros, E.; Nagy, T.; Pigniczki, C.; Kotymán, L.; Balogh, K.V.; Vörös, L. The effect of aquatic birds on the nutrient load and water quality of soda pans in Hungary. Acta Zool. Acad. Sci. Hung. 2008, 54, 207–224. [Google Scholar]
- Gwiazda, R.; Woźnica, A.; Łozowski, B.; Kostecki, M.; Flis, A. Impact of waterbirds on chemical and biological features of water and sediments of a large, shallow dam reservoir. Oceanol. Hydrobiol. Stud. 2014, 43, 418–426. [Google Scholar] [CrossRef]
- Liu, Y.; Hefting, M.M.; Verhoeven, J.T.; Klaassen, M. Nutrient release characteristics from droppings of grass-foraging waterfowl (Anser brachyrhynchus) roosting in aquatic habitats. Ecohydrology 2014, 7, 1216–1222. [Google Scholar] [CrossRef]
- Bresciani, M.; Adamo, M.; De Carolis, G.; Matta, E.; Pasquariello, G.; Vaičiūtė, D.; Giardino, C. Monitoring blooms and surface accumulation of cyanobacteria in the Curonian Lagoon by combining MERIS and ASAR data. Remote Sens. Environ. 2014, 146, 124–135. [Google Scholar] [CrossRef]
- Pilkaityte, R.; Razinkovas, A. Seasonal changes in phytoplankton composition and nutrient limitation in a shallow Baltic lagoon. Boreal Environ. Res. 2007, 12, 551–559. [Google Scholar]
- Zilius, M. Oxygen and Nutrient Exchange at the Sedimentwater Interface in the Eutrophic Boreal Lagoon (Baltic Sea). Ph.D. Thesis, Klaipėda University, Klaipėda, Lithuania, 2011. [Google Scholar]
- Zilius, M.; Giordani, G.; Petkuviene, J.; Lubiene, I.; Ruginis, T.; Bartoli, M. Phosphorus mobility under short-term anoxic conditions in two shallow eutrophic coastal systems (Curonian and Sacca di Goro lagoons). Estuar. Coast. Shelf Sci. 2015, 164, 134–146. [Google Scholar] [CrossRef]
- Dobrowolski, K.A.; Halba, R.; Nowicki, J. The role of birds in eutrophication by import and export of trophic sub-stances of various waters. Limnologica 1976, 10, 543–549. [Google Scholar]
- Zilius, M.; Bartoli, M.; Bresciani, M.; Katarzyte, M.; Ruginis, T.; Petkuviene, J.; Lubiene, I.; Giardino, C.; Bukaveckas, P.A.; de Wit, R.; et al. Feedback mechanisms between cyanobacterial blooms, transient hypoxia, and benthic phosphorus regeneration in shallow coastal environments. Estuar. Coasts 2014, 37, 680–694. [Google Scholar] [CrossRef]
- Risgaard-Petersen, N.; Nicolaisen, M.H.; Revsbech, N.P.; Lomstein, B.A. Competition between ammonia-oxidizing bacteria and benthic microalgae. Appl. Environ. Microbiol. 2004, 70, 5528–5537. [Google Scholar] [CrossRef] [PubMed]
- Racchetti, E.; Longhi, D.; Ribaudo, C.; Soana, E.; Bartoli, M. Nitrogen uptake and coupled nitrification–denitrification in riverine sediments with benthic microalgae and rooted macrophytes. Aquat. Sci. 2017, 79, 487–505. [Google Scholar] [CrossRef]
- Soana, E.; Naldi, M.; Bonaglia, S.; Racchetti, E.; Castaldelli, G.; Brüchert, V.; Viaroli, P.; Bartoli, M. Benthic nitrogen metabolism in a macrophyte meadow (Vallisneria spiralis L.) under increasing sedimentary organic matter loads. Biogeochemistry 2015, 124, 387–404. [Google Scholar] [CrossRef]
- Žydelis, R.; Kontautas, A. Piscivorous birds as top predators and fishery competitors in the lagoon ecosystem. Hydrobiologia 2008, 611, 45–54. [Google Scholar] [CrossRef]
- Pūtys, Ž.; Zarankaitė, J. Diet of the Great Cormorant (Phalacrocorax carbo sinensis) at the Juodkrantė colony, Lithuania. Acta Zool. Litu. 2010, 20, 179–189. [Google Scholar] [CrossRef]
- Paerl, H.W.; Gardner, W.S.; Havens, K.E.; Joyner, A.R.; McCarthy, M.J.; Newell, S.E.; Qin, B.; Scott, J.T. Mitigating cyanobacterial harmful algal blooms in aquatic ecosystems impacted by climate change and anthropogenic nutrients. Harmful Algae 2016, 54, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Levine, S.N.; Schindler, D.W. Influence of nitrogen to phosphorus supply ratios and physicochemical conditions on cyanobacteria and phytoplankton species composition in the Experimental Lakes Area, Canada. Can. J. Fish. Aquat. Sci. 1999, 56, 451–466. [Google Scholar] [CrossRef]
- Downing, J.A.; Watson, S.B.; McCauley, E. Predicting cyanobacteria dominance in lakes. Can. J. Fish. Aquat. Sci. 2001, 58, 1905–1908. [Google Scholar] [CrossRef]
- Smith, V.H. Light and nutrient effects on the relative bio-mass of blue-green algae in lake phytoplankton. Can. J. Fish. Aquat. Sci. 1986, 43, 148–153. [Google Scholar] [CrossRef]
- Klemer, A.R. Effects of nutritional status on cyanobacterial buoyancy, blooms, and dominance, with special reference to inorganic carbon. Can. J. Bot. 1991, 69, 1133–1138. [Google Scholar] [CrossRef]
- De Nobel, W.; Matthijs, H.; Von Elert, E.; Mur, L. Comparison of the light-limited growth of the nitrogen-fixing cyanobacteria Anabaena and Aphanizomenon. New Phytol. 1998, 138, 579–587. [Google Scholar] [CrossRef]
- Carey, C.C.; Ibelings, B.W.; Hoffmann, E.P.; Hamilton, D.P.; Brookes, J.D. Eco-physiological adaptations that favour freshwater cyanobacteria in a changing climate. Water Res. 2012, 46, 1394–1407. [Google Scholar] [CrossRef]
- Jones, A.K.; Cannon, R.C. The release of micro-algal photosynthate and associated bacterial uptake and heterotrophic growth. Eur. J. Phycol. 1986, 21, 341–358. [Google Scholar] [CrossRef]
- Vahtera, E.; Laamanen, M.; Rintala, J.M. Use of different phosphorus sources by the bloom-forming cyanobacteria Aphanizomenon flos-aquae and Nodularia spumigena. Aquat. Microb. Ecol. 2007, 46, 225–237. [Google Scholar] [CrossRef]
- Kirchman, D.L. The ecology of Cytophaga–Flavobacteria in aquatic environments. FEMS Microbiol. Ecol. 2002, 39, 91–100. [Google Scholar] [CrossRef]
- Ducklow, H. Bacterial production and biomass in the oceans. In Microbial Ecology of the Oceans; Kirchman, D.L., Ed.; John Wiley and Sons: New York, NY, USA, 2000; pp. 85–120. [Google Scholar]
- Klimaszyk, P. May a cormorant colony be a source of coliform and chemical pollution in a lake? Oceanol. Hydrobiol. Stud. 2012, 41, 67–73. [Google Scholar] [CrossRef]
- Klimaszyk, P.; Rzymski, P. Impact of cormorant (Phalacrocorax carbo sinensis L.) colonies on microbial pollution in lakes. Limnol. Rev. 2013, 13, 139–145. [Google Scholar] [CrossRef][Green Version]
- Smolders, A.; Smolders, K.; Watkinson, A.; Ryder, D. Reassessing the risk of microbial contamination from roosting cormorants in source water supply reservoirs. Lake Reserv. Manage. 2014, 30, 23–31. [Google Scholar] [CrossRef]
- Gogu-Bogdan, M.; Damoc, I.; Pall, E.; Niculae, M.; Spinu, M. Wild birds as potential vectors forpathogen dissemination on migration routes in the Danube Delta Wetlands. Int. J. Curr. Microbiol. App. Sci. 2014, 3, 890–897. [Google Scholar]
- Currie, D.J.; Kalff, J. Can bacteria outcompete phytoplankton for phosphorus? A chemostat test. Microb. Ecol. 1984, 10, 205–216. [Google Scholar] [CrossRef]
- Cotner, J.B., Jr.; Wetzel, R.G. Uptake of dissolved inorganic and organic bphosphorus compounds by phytoplankton and bacterioplankton. Limnol. Oceanogr. 1992, 37, 232–243. [Google Scholar] [CrossRef]
- Amon, R.M.; Benner, R. Bacterial utilization of different size classes of dissolved organic matter. Limnol. Oceanogr. 1996, 41, 41–51. [Google Scholar] [CrossRef]
- Salcher, M.M.; Hofer, J.; Horňák, K.; Jezbera, J.; Sonntag, B.; Vrba, J.; Posch, T. Modulation of microbial predator-prey dynamics by phosphorus availability: Growth patterns and survival strategies of bacterial phylogenetic clades. FEMS Microbiol. Ecol. 2007, 60, 40–50. [Google Scholar] [CrossRef]
- Ger, K.A.; Urrutia-Cordero, P.; Frost, P.C.; Hansson, L.A.; Sarnelle, O.; Wilson, A.E.; Lürling, M. The interaction between cyanobacteria and zooplankton in a more eutrophic world. Harmful Algae 2016, 54, 128–144. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petkuviene, J.; Vaiciute, D.; Katarzyte, M.; Gecaite, I.; Rossato, G.; Vybernaite-Lubiene, I.; Bartoli, M. Feces from Piscivorous and Herbivorous Birds Stimulate Differentially Phytoplankton Growth. Water 2019, 11, 2567. https://doi.org/10.3390/w11122567
Petkuviene J, Vaiciute D, Katarzyte M, Gecaite I, Rossato G, Vybernaite-Lubiene I, Bartoli M. Feces from Piscivorous and Herbivorous Birds Stimulate Differentially Phytoplankton Growth. Water. 2019; 11(12):2567. https://doi.org/10.3390/w11122567
Chicago/Turabian StylePetkuviene, Jolita, Diana Vaiciute, Marija Katarzyte, Iveta Gecaite, Giorgio Rossato, Irma Vybernaite-Lubiene, and Marco Bartoli. 2019. "Feces from Piscivorous and Herbivorous Birds Stimulate Differentially Phytoplankton Growth" Water 11, no. 12: 2567. https://doi.org/10.3390/w11122567
APA StylePetkuviene, J., Vaiciute, D., Katarzyte, M., Gecaite, I., Rossato, G., Vybernaite-Lubiene, I., & Bartoli, M. (2019). Feces from Piscivorous and Herbivorous Birds Stimulate Differentially Phytoplankton Growth. Water, 11(12), 2567. https://doi.org/10.3390/w11122567